
茶树叶绿体基因组的研究与应用进展
2024-03-12 14:31杨雨青谭娟汪芳彭顺利陈婕谭明燕吕美艳周富裕刘声传
生物技术通报 2024年2期
杨雨青 谭娟 汪芳 彭顺利 陈婕 谭明燕 吕美艳 周富裕刘声传
(1.贵阳学院生物与环境工程学院,贵阳 550005;2.贵州省农业科学院茶叶研究所,贵阳 550006)
茶树(Camellia sinensis)是世界上最古老、最受欢迎的非酒精饮料经济植物,是一种多年生自交不亲合异花授粉作物,具有高度遗传异质性[1-2]。我国茶树种质资源丰富,由于频繁引种驯化、杂交和多倍体化,其起源、进化、分类等方面复杂多样,而难以有效保护开发利用,对其有效鉴定、明确其系统进化及亲缘关系等备受关注[3-4]。
越来越多的植物叶绿体基因组被破译,增强了对植物叶绿体生物学、胞内基因转移、多样性和遗传基础的理解,促进作物改良、生产高价值的农业或生物医药产品[5]。茶树等植物叶绿体基因组具有非重组、单倍体、单亲遗传、进化速率适中、序列和结构高度保守等特征,已应用于资源的鉴定、起源、进化和分类等方面研究[5-6]。Li 等[6]比较分析了一些山茶属植物叶绿体基因组,认为序列重复与插入缺失是诱导叶绿体基因变异的主要动力。Peng等[7]研究发现栽培型茶树叶绿体基因组的序列和结构较野生型茶树更保守,其变异为核苷酸多态性和序列插入。闫明慧等[8]通过叶绿体基因组测序分析,初步明确了‘信阳10 号’与其他茶树资源的进化关系。目前,仍有不少茶树资源的叶绿体基因组未被破译。本文论述了植物叶绿体基因组的起源、遗传方式、基本特征,着重述评了已公布茶树叶绿体基因组的测序技术、特征、基因类型、基因序列,概述了其应用现状,并探讨了其未来应用与发展方向,以期为相关研究提供参考。
1 植物叶绿体基因组
20 亿年前蓝细菌(Cyanobacteria)被真核细胞捕获,最终产生了叶绿体[9-10]。经过一个世纪的争论,20 世纪60 年代叶绿体DNA 的存在被证实[11-12]。1986 年,第一个完整叶绿体基因组在烟草(Nicotiana tabacum)中获得,随后蔬菜、水果、谷物、饮料、油类和糖类等植物的叶绿体基因组相继被破译[13]。20 世纪80 年代末,外源基因被整合至叶绿体基因组中稳定表达,在90 年代表现出典型的母系遗传[12]。Kaundun 等[14]基于简单重复序列(single sequence repeat,SSR)标记分析,表明茶树叶绿体基因组为母系遗传。
叶绿体是植物进行光合作用的细胞器,其完整的结构和功能是光合作用正常进行的前提。脂肪酸、氨基酸、多种植物激素前体和次生代谢物的合成也发生在叶绿体中。叶绿体还参与植物对多种生物与非生物胁迫的响应,被认为是植物响应外界环境的感受器[15]。利用叶绿体的这些特征,在重要作物中研发具有特异农艺性状的产品一直难以实现,但开发的酶等商业产品,已运用于果汁加工、提高棉纤维吸水率、天然清洁剂等[16]。
多数植物的叶绿体DNA 为双链环状,伞藻(Acetabularia)等少数植物为线状[17-19]。多数高等植物的叶绿体基因组为高度保守的四分体结构,包含一段大单拷贝区(large single copy,LSC)、小单拷贝区(small single copy,SSC)以及将这两段分开的、序列相同的一对反向重复(inverted repeat,IR)区(IRa和IRb,长度为20-30 kb)[5]。植物叶绿体基因组为107-218 kb,其长度变化主要由IRs 的收缩和扩张引起,含95-145 个编码基因,按功能不同分为与光合作用、叶绿体基因表达、生物合成相关的基因,以及未知功能的开放阅读框[12,20]。植物叶绿体基因组进化速率适中(约为核基因组进化速率的1/3)[21]、基因含量相对稳定[22]、编码区和非编码区的进化速率差异显著[23-24]、重组率低[25-26]。叶绿体基因组序列揭示了绿色植物系统发育框架,以及叶绿体基因组转移到其他真核生物的复杂历史。叶绿体基因之间不一致的历史信号表明整个质体组可能存在可变的限制条件,进一步理解和缓解这些限制条件可能为生物工程提供新机会[16]。
2 茶树叶绿体基因组
2.1 测序技术
基因组测序技术不断发展,一代(Sanger 测序)、二代(454 焦磷酸、Solexa 和SOLiD 测序)到三代(PacBio SMRT、PacBio Sequl 和纳米孔测序)测序,费用降低、精度显著提高[27]。‘铁观音’ ‘黔茶1 号’‘武夷水仙’ ‘白叶1 号’等叶绿体基因组测序运用二代测序,‘Sangmok’ ‘大红袍’ ‘白鸡冠’ ‘铁罗汉’等以二代测序为主,辅以三代测序。随着测序技术的快速发展,破译更多、更准确的叶绿体基因组,将极大丰富茶树叶绿体基因组数据库。
2.2 基因组特征
目前,已在NCBI 上公布了33 份茶树资源的叶绿体基因组,大小为153 044-158 916 bp,GC 含量为37.2%-37.34%,LSC、SSC、IRs 长度分别为64 665-87 213 bp、16 463-19 155 bp、24 627-27 900 bp。IRs 边界随着叶绿体基因组的进化而有所扩张或收缩,不同茶树资源叶绿体基因组IRs 边界区的连接位置有细微差异,具体为LSC/IRb 或IRa/LSC 边界无差异,IRb/SSC 和SSC/IRa 边界存在微小差异(图1)。相较于野生茶树,栽培型茶树叶绿体基因组长度变化较小,基因数量和GC 含量较稳定[7]。
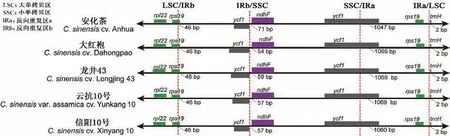
图1 五个茶树品种叶绿体基因组LSC、SSC 和IR 边界比较Fig.1 Comparison of the LSC,IR and SSC border regions among the chloroplast genomes of five tea cultivars
茶树叶绿体基因组有97-141 个编码基因,包括蛋白编码基因60-100 个、tRNA 基因24-47 个、rRNA 基因8 个(少数4-5 个)。多数编码基因含1个内含子,cloP、clpP 和ycf3 等一些特殊基因含2个内含子[28-29]。Yang 等[4]分析发现‘黔茶1 号’IRs区的ycf2 序列最长,编码2 298 个氨基酸。在‘龙井43’和‘武夷水仙’中发现rps12 是反式剪接基因[30]。基因选择压力分析表明,相较于其他茶组资源,‘西莲1 号’中的6 个基因(accD、ndhC、petB、rpl16、rpoC1 和rpoC2)可能处于正选择状态[31](表1)。
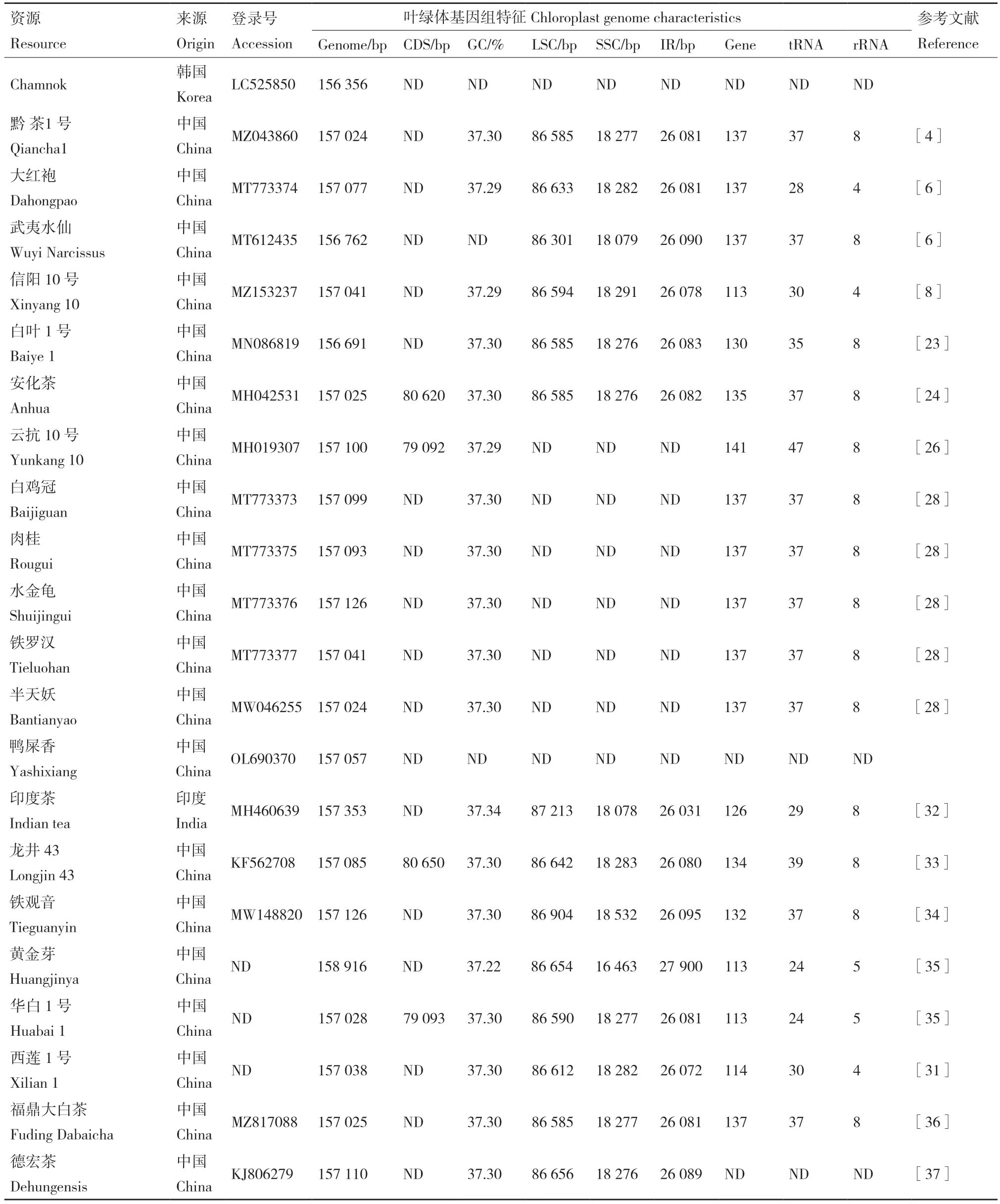
表1 已公布茶树叶绿体基因组Table 1 Published chloroplast genomes of tea plants
共显性SSR 广泛分布于叶绿体基因组中,是研究系统发育和群体遗传学的重要分子标记[43-44]。茶树叶绿体的SSR 主要为单核苷重复序列(A/T 重复为主),有少量双碱基及多碱基重复序列。在‘信阳10 号’叶绿体基因组中共检测到74 个SSR,其中,有56 个单核苷重复(A/T 重复)、二核苷酸重复4 个(AT/AT 重复)[8]。‘信阳10 号’等17 个茶树品种的LSC、IRs 区核苷酸多样性较低(平均为0.001 35),SSC 区相对较高(平均为0.058 08),区域边界高度保守(图1)[8]。成杨等[45]分析发现江华苦茶居群的32 份资源叶绿体DNA 变异率约为0.78%。密码子使用偏差(codon usage bias,CUB)是基因组的一种独特性质,是指编码序列中同义密码子的非随机使用。Yengkhom 等[46]比较分析了阿萨姆茶(C.sinensis var.assamica)、中国茶(C.sinensis var.sinensis)和毛肋茶(C.pubicosta)的叶绿体蛋白质编码基因发现,这些基因富含AT,其高表达与CUB 高度相关。Shi 等[40]研究发现,山茶属植物叶绿体基因ycf15 有完整的编码框,经转录产生但无任何功能,认为假基因转录普遍,叶绿体DNA 转录后加工可能涉及非功能基因的复杂剪接。
2.3 基因类型
茶树叶绿体的功能基因分为光合作用相关基因、自我复制相关基因、生物合成相关基因和其他未知功能基因4 类(表2[8,29,33],图2)。光合作用相关基因包括光合系统Ⅰ基因、光合系统Ⅱ基因、ATP合成酶基因、NADH 脱氢酶基因、二磷酸核酮糖羧化酶大亚基基因、细胞色素复合物基因等。自我复制相关基因包括tRNA 基因、rRNA 基因、RNA 聚合酶亚基基因、核糖体大亚基基因、核糖体小亚基基因等,多数为tRNA 基因。叶绿体中生物合成相关基因,主要包括成熟酶基因、囊膜蛋白乙酰辅酶A羧化酶亚基基因、c 型细胞色素合成基因、转录起始因子基因等。其他未知功能基因,即一些未知功能的开放阅读框,约有7 个。‘信阳10’叶绿体基因组中有47 个光合系统基因、59 个遗传系统基因、5 个生物合成相关基因、2 个未知功能基因[8]。‘龙井43’叶绿体基因组中有45 个光合系统基因、76个遗传系统基因、4 个生物合成相关基因、6 个未知功能基因[29]。‘黄金芽’叶片光系统电子传递途径酶基因CsLFNR1.1 表达响应光照强度变化,随着光强增加表达量提高,可能与其叶片黄化相关[47]。图3 显示,‘安化’‘龙井43’‘云抗10 号’分别存在特有叶绿体基因。结果表明,不同茶树资源的叶绿体基因类型存在一定的差异,具有一定的遗传特性。

表2 茶树叶绿体基因组基因类型Table 2 Gene types of the chloroplast genome in tea plants
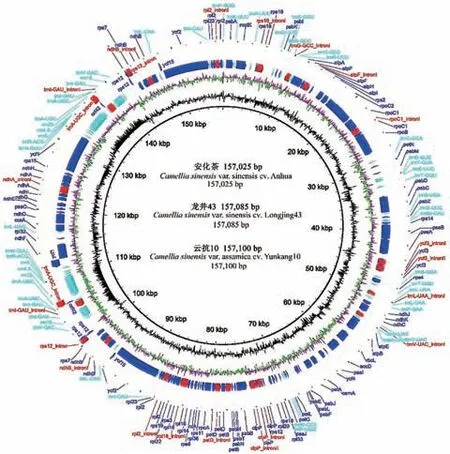
图2 栽培型茶树完整叶绿体基因组的基因图谱Fig.2 Gene map of the complete chloroplast genome of cultivated tea plants
2.4 基因序列
中国栽培型茶、印度阿萨姆茶叶绿体基因组基因序列存在一定差异,栽培型茶与野生型茶树之间的叶绿体基因组基因序列差异更大[8,48]。在栽培型茶树中,最可变的基因是ycf1;一些中国栽培型茶的rps12 无内含子;印度阿萨姆茶和‘云抗10 号’无orf42、ycf1 和ycf15(图2)[7]。栽培型茶树之间的psaA_ycf3、petL_petG 和ycf1_ndhF 一致性相对较低;栽培型茶树和野生型茶树之间的atpH_atpI、trnEUCC_trnT-GGU、psaA_ycf3、ycf15_trnLCAA、ycf1_ndhF 和ndhG_ndhI 一致性相对较低(图2)[7]。中国栽培型茶树之间的ycf1、trnV-GAC_rps12 序列不一致,如‘龙井43’的ycf1 有9 bp 插入序列(TCC TTC TTC/GAA GAA GGA);栽培型阿萨姆茶之间的rnN-GUU_ndhF、rrn5_trnR-ACG 序列不一致,如‘云抗10 号’的rnN-GUU_ndhF 有72 bp 插入序列[7]。野生型茶树叶绿体基因的核苷酸多样性水平约为栽培型茶树的6.6 倍,但栽培型茶树也有少数基因如rps16、rps4、trnL-UAA_intron 的核苷酸多样性水平高于野生型茶树,这些基因主要位于LSC,可用作鉴定资源的潜在分子标记[7]。总体上,栽培型茶树的叶绿体基因组较野生型茶树保守。
3 茶树叶绿体基因组的应用
3.1 分类鉴定
茶组植物是山茶属内分类学问题最多的类群,对其有效分类鉴定是进行这些资源保护与利用的前提与基础[49]。叶绿体基因组数据已应用于茶树系统发育研究,日本等产茶国家基于叶绿体基因组序列,研究山茶属植物间亲缘关系[50-51]。Zhu 等[52]构建30 种山茶属植物的叶绿体基因组系统发育树发现,安龙瘤果茶(C.anlungensis)与膜叶茶(C.leptophylla)、毛叶茶(C.ptilophylla)、毛肋茶(C.pubicosta)、大苞茶(C.grandibracteata)和茶(C.sinensis var.sinensis)亲缘关系较近。Hao 等[53]利用此方法,分析发现大厂茶(C.tachangensis)与秃房茶(C.gymnogyna)、大理茶(C.taliensis)聚为一类,与茶(C.sinensis var.sinensis)、阿萨姆(C.sinensis var.assamica)、白毛茶(C.sinensis var.pubilimba)亲缘关系远。此外,罗祥宗等[54]利用已公布的茶组植物叶绿体全基因组,筛选出16 对引物,共含25 个SNP 位点,可用于茶树品种的母系溯源和鉴别。
相较于核基因组的分子标记,叶绿体DNA 条形码更加简便、高效、准确,叶绿体基因组相关的条形码将显著提高不同物种的鉴别率[55]。生命条形码联盟建议将叶绿体基因rbcL、matK、trnH-psbA 和核基因ITS 作为陆地植物通用的DNA 条形码[56]。温贝贝[57]发现matK 和rbcL 组合对山茶属植物64 份资源的鉴别率最大。毛娟[58]对临沧6 个居群的453个茶树个体的rpl32-trnL 多态性分析,得到7 种叶绿体单倍型。聂传朋等[59]初步筛选出matK、rcbL 可用作茶树DNA 条形码。
叶绿体基因组具有单亲遗传、不发生重组和非编码区比编码区进化速率更快的特点,而用于居群遗传多样性有效评价[60-61]。基于核SSR 和叶绿体DNA 序列分析结果一致,城步峒茶的遗传变异主要存在于居群内[62]。进化速率较快的叶绿体基因组片段(rcbL、rpl16、trnH-psbA、trnL-F 和rpl32-trnL)在茶树等植物遗传多样性分析中得到了不少应用[63-66]。
3.2 起源、驯化研究
中国型茶树、中国型阿萨姆茶树与印度型阿萨姆茶树之间的谱系是否一致长期有争议。比较叶绿体基因分析表明,印度型阿萨姆茶树、中国型茶树可能经历了不同驯化,起源不同[67]。Li 等[48]通过比较分析‘武夷水仙’(三倍体)、中国型茶树、中国型阿萨姆茶、印度型阿萨姆茶等山茶属植物的叶绿体基因组,支持3 个独立驯化起源假说。吴艾琳[68]利用叶绿体rpl32-trnL 和trnG-S 分析了大理茶和厚轴茶(C.crassicolumna)27 个居群的遗传多样性和遗传结构,认为茶树存在分别以大理茶、厚轴茶为起源中心的2 条传播途径。比较叶绿体基因组分析表明,‘凤凰单丛’、福建乌龙茶品种与其他栽培型茶树聚为一类,但这两种乌龙茶资源相对独立地交叉嵌入山茶属植物中,‘凤凰单丛’与其他乌龙茶品种之间亲缘关系近,总体上乌龙茶资源的叶绿体基因组变异低、进化保守[48]。
3.3 白化机制研究
多数白化茶树品种适制名优绿茶,但其白化分子机制仍不明确[35]。茶树白化现象与叶绿体的发育等密切相关,一些叶绿体基因参与茶树叶绿体发育,但这些基因的RNA 编辑位点仍未明确[69]。在细胞器中,不同家族的RNA 编辑因子组装成RNA编辑复合体,特异识别编辑位点进行编辑[70]。Zhao等[69]对‘华白1 号’‘白叶1 号’和‘龙井43’中的11 个RNA 编辑位点差异编辑效率测定,鉴定到10 个多细胞器RNA 编辑因子(multiple organellar RNA editing factor,MORF),其中‘华 白1 号’的CsMORF9.2 表达水平显著下调,可能参与了其新梢白化。此外,Zhang 等[71]对6 个茶树品种叶绿体matK 和ndhD 的RNA 编辑分析发现,matK-701 可能参与了叶色变化。
4 总结与展望
我国茶树种质资源丰富,很有必要对其复杂的起源、进化、分类、特异性状等进行有效鉴评,进而对其高效保护与创新利用。相较于核基因组,结构简单的、较小的、保守的叶绿体基因组更有助于这方面的研究。不少茶树资源的起源、驯化机制还未明确。目前已破译了少量茶组植物叶绿体基因组,揭示了部分叶绿体基因组的基因类型和序列特征,但其结构变异机制仍未明确。叶绿体基因的RNA 编辑、水平基因转移、核质互作等方面仍需深入研究。
叶绿体基因组有约50%部分包含不保守的基因间隔区和调控序列,若缺乏这些序列,叶绿体基因组转化难以成功。茶树中还未明确通用的叶绿体DNA 条形码,一些单个叶绿体基因片段已被开发成DND 条形码,用于茶树分化研究,但对整个叶绿体基因组在茶树种内分化特别是新品种分化的有效性知之甚少。
随着高通量测序及生物技术的快速发展,茶组植物叶绿体基因组测序数量和精度的增加,对茶树叶绿体基因组的深入研究,将会提高茶树资源的分类鉴定效率,进一步明确其起源、演化、驯化机制,开发高效通用DNA 条形码,深入揭示白化茶树白化分子机理,推动叶绿体基因工程发展。
猜你喜欢
今日农业(2021年11期)2021-08-13
乡村地理(2018年2期)2018-09-19
湖南农业(2016年3期)2016-06-05
广西林业科学(2016年1期)2016-03-20
音乐天地(音乐创作版)(2016年11期)2016-02-05
遗传(2014年3期)2014-02-28
世界科学(2014年8期)2014-02-28
茶叶通讯(2014年2期)2014-02-27
食品科学(2013年6期)2013-03-11
世界科学(2013年6期)2013-03-11
