
猪源和犬源ELF4蛋白的亚细胞定位比较研究
2024-03-11 03:23:48高翠翠喻娇唐青海侯鑫军刘婷全飞杨刘建行赵婷芳赵铖朱钰英危艳武
畜牧与兽医 2024年3期
高翠翠,喻娇,唐青海*,侯鑫军,刘婷,全飞杨,刘建行,赵婷芳,赵铖,朱钰英,危艳武*
(1. 衡阳师范学院生命科学学院/南岳山区生物资源保护与利用湖南省重点实验室,湖南 衡阳 421008;2. 中国农业科学院哈尔滨兽医研究所/兽医生物技术国家重点实验室,黑龙江 哈尔滨 150069)
E26转录因子(E26 transformation specific,ETS)家族分子是多细胞生物的一类转录因子,参与发育和分化等多个关键生物学过程[1]。E74 样因子4(E74-like factor 4,ELF4)是ETS家族的重要一员,该蛋白由N端激活区域、与DNA共识序列结合的ETS区域和富含丝氨酸、苏氨酸和脯氨酸的区域组成。人ELF4(hELF4)参与细胞多种生理或病理的过程,如肿瘤发生[2]、DNA损伤修复[3]、天然免疫[4]、细胞周期调控[5]和成骨分化[6]等。研究表明,hELF4高表达可促进人胰岛素瘤细胞增殖[7],然而在急性髓性白血病中,hELF4则是作为一个抑癌基因起作用[8]。hELF4对胃癌细胞的增殖和侵袭有正向调节作用[9],肿瘤来源的外泌体通过传递LINC01091调控microRNA-128-3p/ELF4/CDX2轴促进胃癌的生长和转移[10]。此外,hELF4在免疫调节中起着重要作用。You等[11]首次发现hELF4是一种Ⅰ型干扰素(IFNs)转录因子,并通过直接调节Ⅰ型IFNs反应来抑制病毒的复制。hELF4功能突变缺失导致人类自身炎症和免疫缺陷疾病[12]。肠道hELF4是维持肠道内稳态、减轻酒精诱导的肝脏脂肪变性和损伤的重要宿主保护因子[13]。hELF4蛋白功能区研究比较透彻,定位在细胞核中,与其作为转录因子的功能关联密切[14]。

A. cELF4基因截短示意图;B. sELF4基因截短示意图。图中数字表示片段长度,单位为bp。
目前尚无猪E74样因子4(sELF4)和犬E74样因子4(cELF4)的亚细胞定位比较研究的报道。本课题组成员前期成功克隆了sELF4基因,并制备了免疫活性和特异性良好的鸡抗sELF4多克隆抗体[15],克隆cELF4基因,体外表达重组cELF4蛋白并制备了相应的抗体[16]。为揭示sELF4和cELF4蛋白的亚细胞定位特征,阐明二者亚细胞定位的异同,本研究采用PCR截短扩增基因片段、以绿色荧光蛋白(GFP)为报告基因,构建真核表达载体,转染细胞,观察细胞定位,研究结果将为进一步探究sELF4 和cELF4功能奠定基础。
1 材料与方法
1.1 材料
感受态细胞Top 10购自天根生化科技(北京)有限公司;KOD FX Neo高保真酶购自东洋纺(上海)生物科技有限公司;Ligation Mix、内切酶以及XfectTM转染试剂均购自宝日医生物技术(北京)有限公司;Hoechst 33342购于Abbkine Scientific公司;DMEM和胎牛血清培养基购自Gibico公司;含sELF4基因全长质粒pMD19T-sELF4、含cELF4基因全长质粒pMD19T-cELF4 以及宫颈癌细胞HeLa由南岳山区生物资源保护与利用湖南省重点实验室构建或保存。
1.2 方法
1.2.1 引物的设计
根据本课题组前期已经测定的cELF4基因序列(GenBank No. MZ198105)、sELF4基因序列(GenBank No. KU097322),用Primer Premier 5.0软件分别设计cELF4和sELF4基因全长PCR扩增引物和截短扩增引物,各片段相对位置如图1所示,引物由深圳华大基因股份有限公司合成,引物序列信息见表1和表2。

表1 cELF4引物信息

表2 sELF4引物信息
1.2.2 sELF4和cELF4基因的PCR扩增与重组真核表达载体的构建
以pMD19T-cELF4 或pMD19T-sELF4载体为模板,利用KOD FX Neo高保真酶进行cELF4和sELF4截短序列的PCR扩增。PCR反应体系为:DNA模板2 μL、KOD FX Neo 1 μL、2× PCR Buffer for KOD FX Neo 25 μL,2 mmol/L dNTPs 10 μL,上下游引物各1 μL,灭菌去离子水10 μL。PCR反应程序为:95 ℃ 5 min;98 ℃ 10 s,60 ℃ 30 s,68 ℃ 2 min,35个循环;69 ℃ 7 min;4 ℃保存。PCR产物经1.2%琼脂糖凝胶电泳检测后,进行双酶切,与荧光蛋白报告载体pEGFP-C1连接、转化Top 10感受态细胞,阳性克隆采用双酶切和序列测定进行鉴定。
1.2.3 重组质粒的转染、染色与亚细胞定位的观察
将长满单层的HeLa细胞经胰蛋白酶消化、以适当密度接种96 孔板内,采用含10%胎牛血清的DMEM培养基在37 ℃、5% CO2条件下培养24 h;采用XfectTM转染试剂将重组真核表达质粒转染HeLa细胞。转染后48 h,弃培养基,0.85%的生理盐水洗涤1次,加入Hoechst 33342染液、室温孵育15 min,弃染液、用0.85%的生理盐水洗涤3次,荧光倒置显微镜下观察荧光蛋白细胞定位,绿色荧光显示为目的蛋白,蓝色荧光为细胞核。
1.2.4 sELF4和cELF4基因的序列比较分析
利用DNAStar 7.0 对cELF4和sELF4蛋白的氨基酸序列进行对比分析。
2 结果
2.1 sELF4和cELF4基因截短片段的真核表达载体的构建
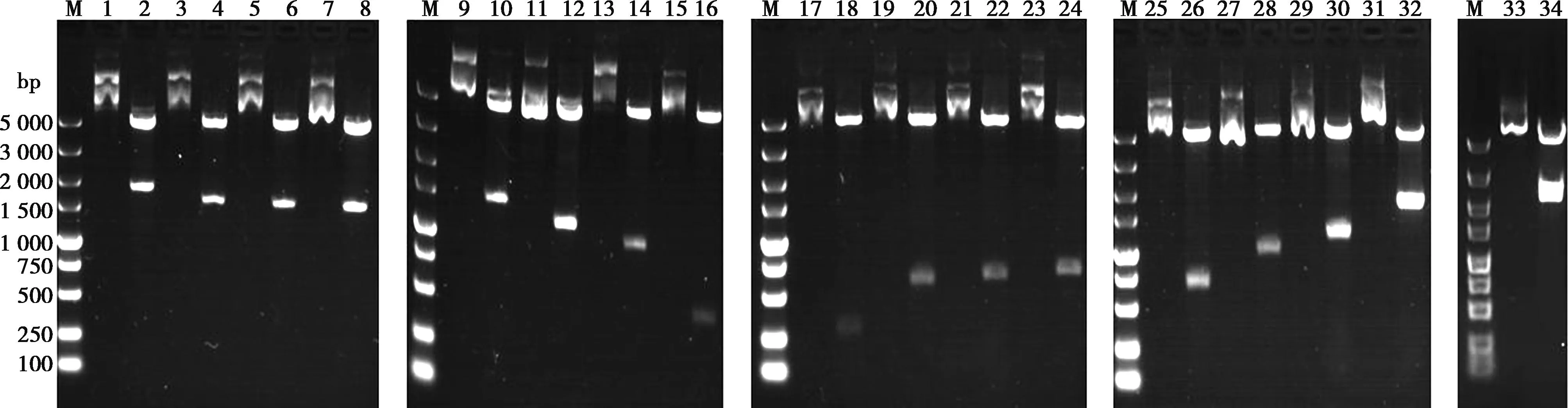
经PCR扩增,得到cELF4基因的13个分子量分别为1 992、1 734、1 476、1 443、1 386、1 119、441、258、516、549、606、873和1 551 bp的核酸片段(图2);克隆至pEGFP-C1载体中构建13个重组真核表达质粒,双酶切鉴定结果显示,13个真核质粒均为阳性(图3)。经PCR扩增得到sELF4基因的17个片段,分子量分别为1 989、1 731、1 473、1 440、1 404、1 383、1 116、948、438、258、516、549、585、606、873、1 041和1 551 bp(图4);克隆至pEGFP-C1载体中构建17个重组真核表达质粒,经双酶切鉴定均为阳性(图5)。

M. DL2000 Marker;1~13. 分别为cELF4-1-1992、cELF4-259-1992、cELF4-517-1992、cELF4-550-1992、cELF4-607-1992、cELF4-874-1992、cELF4-1552-1992、cELF4-1-258、cELF4-1-516、cELF4-1-549、cELF4-1-606、cELF4-1-873和cELF4-1-1551的PCR产物。

M. DL5000 Marker;1. 重组质粒pEGFP-C1-cELF4-1-1992的双酶切产物;2. 重组质粒pEGFP-C1-cELF4-1-1992;3. 重组质粒pEGFP-C1-cELF4-259-1992的双酶切产物;4. 重组质粒pEGFP-C1-cELF4-259-1992;5. 重组质粒pEGFP-C1-cELF4-517-1992的双酶切产物;6. 重组质粒pEGFP-C1-cELF4-517-1992;7. 重组质粒pEGFP-C1-cELF4-550-1992的双酶切产物;8. 重组质粒pEGFP-C1-cELF4-550-1992;9. 重组质粒pEGFP-C1-cELF4-607-1992的双酶切产物;10. 重组质粒pEGFP-C1-cELF4-607-1992;11. 重组质粒pEGFP-C1-cELF4-874-1992的双酶切产物;12. 重组质粒pEGFP-C1-cELF4-874-1992;13. 重组质粒pEGFP-C1-cELF4-1552-1992的双酶切产物;14. 重组质粒pEGFP-C1-cELF4-1552-1992;15. 重组质粒pEGFP-C1-cELF4-1-258的双酶切产物;16. 重组质粒pEGFP-C1-cELF4-1-258;17. 重组质粒pEGFP-C1-cELF4-1-516的双酶切产物;18. 重组质粒pEGFP-C1-cELF4-1-516;19. 重组质粒pEGFP-C1-cELF4-1-549的双酶切产物;20. 重组质粒pEGFP-C1-cELF4-1-549;21. 重组质粒pEGFP-C1-cELF4-1-606的双酶切产物;22. 重组质粒pEGFP-C1-cELF4-1-606;23. 重组质粒pEGFP-C1-cELF4-1-873的双酶切产物;24. 重组质粒pEGFP-C1-cELF4-1-873;25. 重组质粒pEGFP-C1-cELF4-1-1551的双酶切产物;26. 重组质粒pEGFP-C1-cELF4-1-1551。
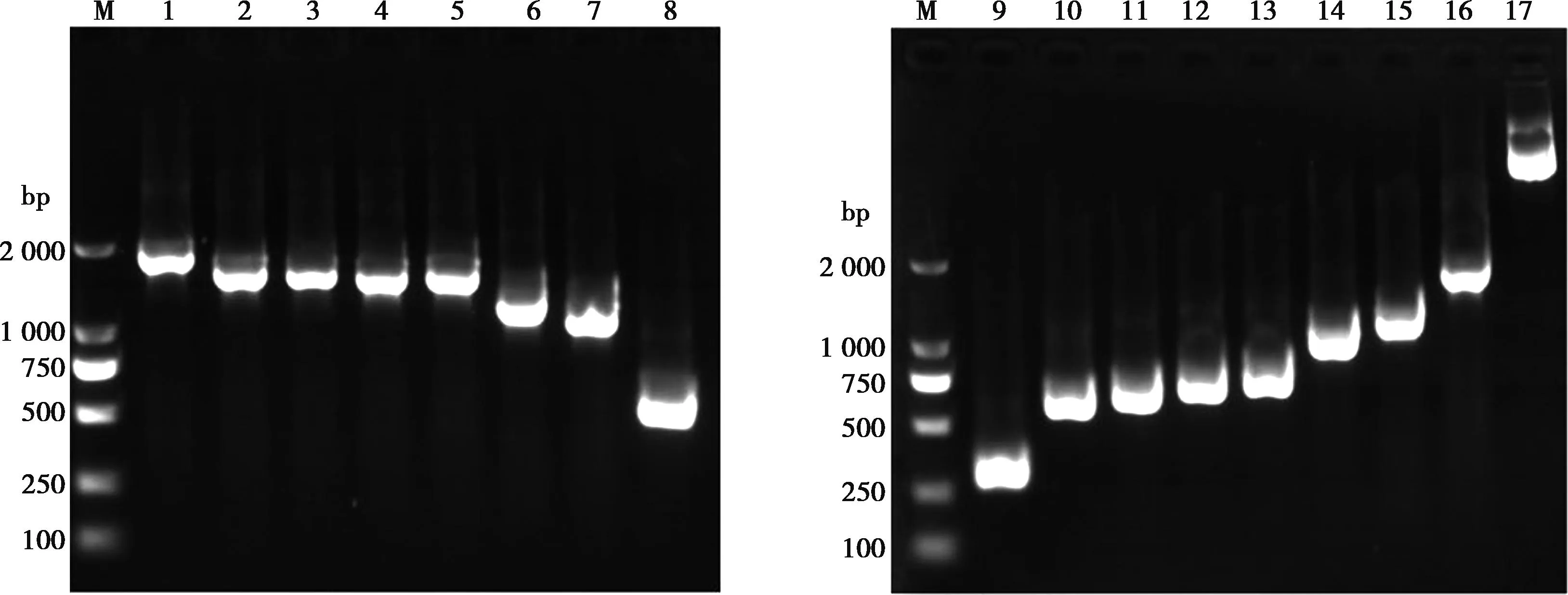
M. DL2000 Marker;1~16. 分别为sELF4-259-1989、sELF4-517-1989、sELF4-550-1989、sELF4-586-1989、sELF4-607-1989、sELF4-874-1989、sELF4-1042-1989、sELF4-1552-1989、sELF4-1-258、sELF4-1-516、sELF4-1-549、sELF4-1-585、sELF4-1-606、sELF4-1-873、sELF4-1-1041和sELF4-1-1551的PCR产物;17. pEGFP-C1-sELE4-1-1989质粒。
M. DL5000 Marker;1. 重组质粒pEGFP-C1-sELF4-259-1989;2. 重组质粒pEGFP-C1-sELF4-259-1989的双酶切产物;3. 重组质粒pEGFP-C1-sELF4-517-1989;4. 重组质粒pEGFP-C1-sELF4-517-1989的双酶切产物;5. 重组质粒pEGFP-C1-sELF4-550-1989;6. 重组质粒pEGFP-C1-sELF4-550-1989的双酶切产物;7. 重组质粒pEGFP-C1-sELF4-586-1989;8. 重组质粒pEGFP-C1-sELF4-586-1989的双酶切产物;9. 重组质粒pEGFP-C1-sELF4-607-1989;10. 重组质粒pEGFP-C1-sELF4-607-1989的双酶切产物;11. 重组质粒pEGFP-C1-sELF4-874-1989;12. 重组质粒pEGFP-C1-sELF4-874-1989的双酶切产物;13. 重组质粒pEGFP-C1-sELF4-1042-1989;14. 重组质粒pEGFP-C1-sELF4-1042-1989的双酶切产物;15. 重组质粒pEGFP-C1-sELF4-1552-1989;16. 重组质粒pEGFP-C1-sELF4-1552-1989的双酶切产物;17. 重组质粒pEGFP-C1-sELF4-1-258;18. 重组质粒pEGFP-C1-sELF4-1-258的双酶切产物;19. 重组质粒pEGFP-C1-sELF4-1-516;20. 重组质粒pEGFP-C1-sELF4-1-516的双酶切产物;21. 重组质粒pEGFP-C1-sELF4-1-549;22. 重组质粒pEGFP-C1-sELF4-1-549的双酶切产物;23. 重组质粒pEGFP-C1-sELF4-1-585;24. 重组质粒pEGFP-C1-sELF4-1-585的双酶切产物;25. 重组质粒pEGFP-C1-sELF4-1-606;26. 重组质粒pEGFP-C1-sELF4-1-606的双酶切产物;27. 重组质粒pEGFP-C1-sELF4-1-873;28. 重组质粒pEGFP-C1-sELF4-1-873的双酶切产物;29. 重组质粒pEGFP-C1-sELF4-1-1041;30. 重组质粒pEGFP-C1-sELF4-1-1041的双酶切产物;31. 重组质粒pEGFP-C1-sELF4-1-1551;32. 重组质粒pEGFP-C1-sELF4-1-1551的双酶切产物;33. 重组质粒pEGFP-C1-sELF4-1-1989;34. 重组质粒pEGFP-C1-sELF4-1-1989的双酶切产物。
2.2 sELF4和cELF4的亚细胞定位
亚细胞定位结果显示,cELF4的1~86 aa(1~258 bp)、292~663 aa(874~1 992 bp)、518~663 aa(1 552~1 992 bp)荧光分布在整个细胞;1~172 aa(1~516 bp)、1~183 aa(1~549 bp)、1~202 aa(1~606 bp)荧光分布在细胞质中;1~291 aa(1~873 bp)、1~517 aa(1~1 551 bp)、1~663 aa(1~1 992 bp)、87~663 aa(259~1 992 bp)、173~663 aa(517~1 992 bp)、184~663 aa(550~1 992 bp)、203~663 aa(607~1 992 bp)荧光分布在细胞核中,1~291 aa(1~873 bp)、203~663 aa(607~1 992 bp)均定位在细胞核,cELF4的核定位信号肽分布在203~291 aa,292~663 aa (874~1 992 bp)定位在细胞质和细胞核中均有分布,且荧光蛋白发生凝聚现象(如图6)。

图6 cELF4不同截短片段与EGFP融合表达的亚细胞定位观察(标尺=100 μm)
sELF4的1~86 aa(1~258 bp)、1~202 aa(1~606 bp)、292~662 aa(874~1 989 bp)、348~662 aa(1 042~1 989 bp)、518~662 aa(1 552~1 989 bp)荧光分布在整个细胞,1~172 aa(1~516 bp)、1~183 aa(1~549 bp)、1~195 aa(1~585 bp)荧光分布在细胞质中,1~291 aa(1~873 bp)、1~347 aa(1~1 041 bp)、1~517 aa(1~1 551 bp)、1~662 aa(1~1 989 bp)、87~662 aa(259~1 989 bp)、173~662 aa(517~1 989 bp)、184~662 aa(550~1 989 bp)、196~662 aa(586~1 989 bp)、203~662 aa(607~1 989 bp)荧光分布在细胞核中,1~291 aa(1~873 bp)、203~662 aa(607~1 989 bp)均定位在细胞核,sELF4的核定位信号肽分布也在203~291 aa(607~873 bp)(如图7)。

图7 sELF4不同截短片段与EGFP融合表达的亚细胞定位观察(标尺=100 μm)
通过对比发现,sELF4的1~202 aa(1~606 bp)分布在整个细胞中,在sELF4-585-606之间还存在一段不完全的定位信号肽,在sELF4 196~291 aa(586~873 bp)之间有可能存在两个核定位信号肽。cELF4 1~202 aa(1~606 bp)仅仅分布在细胞质中,在cELF4 516~606 aa之间还存在一段细胞质信号肽。 sELF4 292~662 aa和cELF4 292~663 aa在细胞质和细胞核中均有分布,且部分荧光蛋白发生凝聚现象。
2.3 sELF4和cELF4基因序列比较分析
氨基酸序列对比分析结果显示(图8),cELF4蛋白和sELF4蛋白的氨基酸序列中第169~303 aa序列完全一致,说明该区段十分保守。第 320~400 aa序列中差异位点相对较多,共有20个变异位点;cELF4在第357 aa缺失H,在第70~160 aa共有13个差异位点,sELF4在第116、117位缺失S和A。

红色背景区代表氨基酸差异区域;无背景色区表示氨基酸保守区域。
3 讨论
hELF4是近年来新发现的一个在肿瘤发生、发展及肿瘤免疫中至关重要的癌基因[2]。研究表明,hELF4参与细胞周期的调节[5],介导成脂成骨细胞的分化[6],参与DNA损伤修复[3]。DNA损伤会破坏代谢稳态并引发炎症反应,Shi等[17]研究发现,hELF4的敲低增加了4细胞胚胎的DNA损伤,这表明hELF4通过控制基因组完整性进而影响猪的早期胚胎发育。Kosti等[18]的研究结果表明hELF4是miRNA-转录因子网络的关键组成部分,是胶质母细胞瘤受体信号通路和脂质动态的桥梁调节因子。卫国红等[19]研究发现,hELF4能促进胰岛素瘤细胞增殖并抑制细胞凋亡,将其与其他类型肿瘤相比,发现ELF4的功能具有一定的多样性,在不同类型肿瘤中,由于hELF4定位在细胞质或细胞核中而发挥功能的差异性,可能激活不同的信号转导通路,而其原因可能是在细胞内相互结合的关键蛋白因子不同。HeLa细胞是一种宫颈癌细胞,具有无限增殖潜能,且繁殖速度快、可连续传代,被广泛应用于肿瘤研究、细胞培养以及生物学试验等。由于其稳定且增殖活性好、外源基因的蛋白表达量高、便于蛋白质的亚细胞定位观察,常用于各类蛋白质定位研究的模式细胞[19]。
本研究以GFP为报告基因,将重组质粒转染HeLa细胞,观察目的蛋白的亚细胞定位,确定了sELF4全长蛋白和cELF4全长蛋白均定位在细胞核中,这与人ELF4蛋白定位一致。cELF4和sELF4的核定位信号分布在203~291 aa(607~873 bp),而人ELF4包含两个核定位信号分别位于173~183 aa和196~202 aa[14],这说明不同种属来源的ELF4蛋白的核定位信号肽分布有差异。一般来说,信号肽大概为16~26个氨基酸,而本研究发现信号肽分布在203~291 aa,推测cELF4和sELF4可能存在多个核定位信号;cELF4和sELF4的亚细胞定位存在差异,在cELF4-516-606之间还存在1段细胞质信号肽,而sELF4-516-585之间存在1段细胞质信号肽。此外,发现去除了N端核定位信号肽的sELF4 292~662 aa和cELF4 292~663 aa在细胞质和细胞核中均有分布,且部分荧光蛋白发生凝聚现象,而这一现象在全长蛋白的表达中并未观察到,推测这种现象可能与蛋白质翻译后折叠、修饰或亲水性发生改变有关。Hamdan等[20]研究发现,内质网应激会导致胞质蛋白稳态缺陷和蛋白质聚集体的形成。蛋白质构象异常或折叠错误可导致其结合成更大的、不可溶的结构,蛋白质发生聚集[21],这与应激或衰老条件下的翻译错误、突变或缺乏寡聚组装伴侣有关[22]。Jamar等[23]研究结果发现,在缺乏无义介导的mRNA降解、翻译停滞降解和无终止降解的突变体中,mRNA监测途径的缺失会导致蛋白质聚集增加。喻娇等[16]研究发现cELF4基因(GenBank No. MZ198105)与sELF4基因(GenBank No.KU097322)的氨基酸相似性为91.8%。本研究利用DNAStar 7.0 软件对cELF4基因编码的氨基酸序列与sELF4基因编码的氨基酸序列进行对比分析,发现二者在某特定区域(169~303 aa)保守性极高,亚细胞定位试验显示cELF4和sELF4的核定位信号均分布在203~291 aa,表明来源于猪和犬的ELF4蛋白核定位信号肽高度保守。
综上所述,本研究明确了cELF4和sELF4基因的信号肽定位序列分布特征,为后续开展基因功能研究提供了依据。
猜你喜欢
河北科技大学学报(2023年5期)2023-11-09 01:44:44
食品科学(2018年10期)2018-05-23 01:27:28
山西医科大学学报(2017年11期)2017-12-01 07:22:37
浙江农业学报(2017年3期)2017-04-08 02:39:02
食品研究与开发(2016年9期)2016-06-13 08:26:16
哈尔滨商业大学学报(自然科学版)(2016年1期)2016-04-22 07:04:42
华东理工大学学报(自然科学版)(2015年4期)2015-12-01 04:00:23
华东理工大学学报(自然科学版)(2015年4期)2015-12-01 04:00:23
西南医科大学学报(2015年1期)2015-08-22 13:01:46
农村百事通(2015年8期)2015-05-19 08:07:27
