
铀对2种小球藻生长和光合作用的影响
2024-03-08 12:32王宇涵唐运来
西北植物学报 2024年1期
王 璇 ,王宇涵 ,唐运来,2* ,陈 梅,2
(1 西南科技大学 生命科学与工程学院,四川绵阳 621010;2 西南科技大学 核废物与环境安全国防重点学科实验室 四川绵阳 621010)
目前,核技术广泛应用于工业、农业、军事、医疗及能源等行业领域,虽为人类带来了便利及巨大经济效益,但也导致大量放射性核素释放到环境中。环境中的放射性核素会破坏生态平衡,最终会随着食物链富集至人体内,危害人体健康。2011年,日本福岛第一核电厂发生核泄漏事故,导致大量放射性核素释放到太平洋,海洋环境的放射性污染问题成为世人关注的焦点[1-2]。日本福岛核事故后,监测数据表明在2011至2012年从北太平洋公海渔场采集的所有鱼类样品中均含有放射性核素137Cs[3],中国也已将U、137Cs、90Sr等核素列为海洋放射性污染的主要监测对象[4]。最近,日本政府计划在2023年内把福岛核事故产生的130万t核废水排入太平洋,此举必将对海洋生态系统的稳定性和海洋生物多样性产生重大影响[5]。因此,开展水体放射性核素的生物学效应及生态风险评估研究,筛选放射性污染的指示生物和生物标志物,建立放射性污染生物监测技术体系具有重要的科学意义和实践应用价值[6-7]。
藻类和水生植物是水生态系统的重要组成部分,在水体放射性污染监测技术体系中具有重要作用和地位。目前国际上有一些将藻类和水生植物用于水体放射性污染生物监测的研究报道,如Herlory等[8]报道铀矿开采会引起周边水生态系统中硅藻群落的结构变化,硅藻可以作为水体铀污染的指示生物。Gosselin 等[9]研究发现波西多尼亚海草(Posidonia oceanica)可以作为水体重金属及核素污染生物监测的指示生物,其在数月内可以精准地反映当前环境中核素及重金属的变化情况,但反映的精确性在较长时间后下降。而国内对于水体核素污染的风险评估与生物监测方面的研究非常少。
目前藻类生物监测技术主要通过筛选指示生物以及测定藻类的生长抑制率、生物种类多样性指数、硅藻生物指数等指标进行水质污染程度的评价[10-11],具有测定周期较长、操作繁琐复杂、灵敏度差等缺陷。叶绿素荧光技术是近年来发展起来的一种可以应用于评价各种逆境胁迫对植物、藻类生物光合作用影响的新技术,具有无损检测和灵敏快速等优点[12],将藻类生物监测技术与叶绿素荧光动力学技术相结合,可大大节省测定时间,提高生物监测的灵敏性[13]。因此,本试验选取模式微藻普通小球藻(Cholorella vulgaris)和从生态脆弱地区四川黄龙国家自然保护区采集的普通小球藻作为研究对象,分析不同浓度铀处理对2种微藻生长以及光合作用参数的影响,比较2种微藻对水体铀污染的响应差异,期望筛选出一些比较灵敏的基于光合作用的尤其是叶绿素荧光参数的水体铀污染生物监测指标,可以为水体铀污染的风险评价与生物监测技术提供一些新的理论依据和技术途径。
1 材料和方法
1.1 试验材料
试验材料为2种来自于不同生境的普通小球藻(C.vulgaris),以及蓝藻PCC6803 和莱茵衣藻。其中,一种普通小球藻、蓝藻PCC6803和莱茵衣藻(Chlamydomonas reinhardtii)来自中国科学院水生生物研究所,另外一种普通小球藻由课题组从四川黄龙国家自然保护区采集的藻泥中分离纯化而来[14],经过初步鉴定为一种普通小球藻,为了便于区分在本文中将其命名为黄龙小球藻。
1.2 试验设计
将分离纯化后的4种微藻在光照恒温摇床中进行扩增培养,普通小球藻、蓝藻PCC6803、莱茵衣藻的培养温度为25℃,黄龙小球藻的培养温度为15℃,光照度为4 000 lx,光/暗比为12 h/12 h,摇床转速为90 r/min。本试验设置0,0.5,1,5,10,20 mg/L U6+6个铀处理浓度。用培养基将醋酸双氧铀UO2(CH3COO)2·2H2O 制成100 mg/L U6+的母液,根据微藻的生长曲线选取生长状态良好一致的藻液10 m L,普通小球藻、黄龙小球藻、莱茵藻的藻液用TAP 培养基与铀母液进行定容,蓝藻PCC6803藻液采用BG-11培养基与铀母液进行定容。使各处理最终体积均为50 m L。每种铀处理浓度均设置5次重复,接种后按照每种藻设定的温度、光照度和光暗比放入光照恒温摇床中培养。在处理后的第3,5,7,10,14天取样测定各项指标。
1.3 测定指标及方法
1.3.1 藻类相对生长速率
分光光度法是测定藻细胞密度最常用且技术最成熟的方法之一,多种微藻的藻细胞密度在波长680 nm 下与光密度呈直线关系[15],相关研究发现普通小球藻等绿藻的光吸收峰在680 nm 处,蓝藻PCC6803的光吸收峰波长为720~730 nm 处[16]。根据文献及预实验确定普通小球藻、莱茵藻、黄龙小球藻光吸收峰波长也为680 nm,蓝藻PCC6803的光吸峰波长为720 nm。取3 m L 藻液加入比色皿中,测定普通小球藻、黄龙小球藻、莱茵藻在680 nm处的吸光度A680,及蓝藻PCC6803在720 nm 处的吸光度A720,根据A680和A720值绘制4种藻类相对生长速率的曲线图。
1.3.2 藻类光合放氧速率
采用Chlorolab2 液相氧电极(Hansatech,英国)测定藻类的光合放氧速率。测试前,将样品藻液置于饱和光强下适应30 min。反应杯中测试样品藻液体积为2 m L,测量光照强度为800μmol/(m2·s)。
1.3.3 藻类叶绿素含量
采用热乙醇提取法提取藻类叶绿素a、叶绿素b,并采用分光光度法测定含量。取3 m L藻液于10 m L离心管中,12 000 r/min下离心5 min,去掉上清液后,加入5 m L 95%乙醇重新悬浮藻饼,并用锡纸包裹离心管后置于4℃冰箱冷藏过夜。次日将离心管置于55℃水浴锅中水浴加热60 min,后于12 000 r/min下离心5 min,用95%乙醇定容至5 m L,用移液枪吸取3 m L叶绿体色素提取液于比色皿内,以95%乙醇为空白,分别在波长665 nm、649 nm 下测定吸光度值A665和A649值。根据公式(1)、(2)分别计算普通小球藻、黄龙小球藻的叶绿素a(Ca,mg/L)和叶绿素b (Cb,mg/L)含量。
1.3.4 藻类叶绿素荧光参数
采用Dual Pam-100双通道调制叶绿素荧光仪(WALZ,德国)测定藻类的叶绿素荧光参数。测量前先进行30 min暗适应,采用Induction Curve测定程序,测量光照强度为229μmol/(m2·s),作用光照强度为855μmol/(m2·s),饱和脉冲光照强度为5 000μmol/(m2·s)。分别测定F0、Fm、F、F0′和Fm′等参数并计算得到光系统Ⅱ最大光化学量子产率(Fv/Fm)、实际光化学量子产率[Y(Ⅱ)]和光系统Ⅱ相对电子传递速率(r ETR)等叶绿素荧光动力学参数[17-18]。
1.4 数据分析
采用Excel软件对数据进行整理,使用Origin 2021 软件作图并进行趋势分析,所有数据使用SPSS27.0软件进行显著性分析和多因素非线性回归分析并构建回归方程。
2 结果与分析
2.1 不同浓度铀对藻类相对生长速率的影响
如图1所示,普通小球藻 (图1,A)、黄龙小球藻(图1,B)、蓝藻PCC6803(图1,C)、莱茵衣藻(图1,D)4种微藻相对生长速率在不同浓度铀处理下表现明显不同,黄龙小球藻的生长速率在处理浓度间差异均较大,其他3种微藻生长速率则均明显分为两组,在低浓度(对照、0.5 mg/L、1.0 mg/L)处理下明显较高,在高浓度(5~20 mg/L)处理明显较低。后续仅分析2种小球藻的生长速率和光合作用差异。
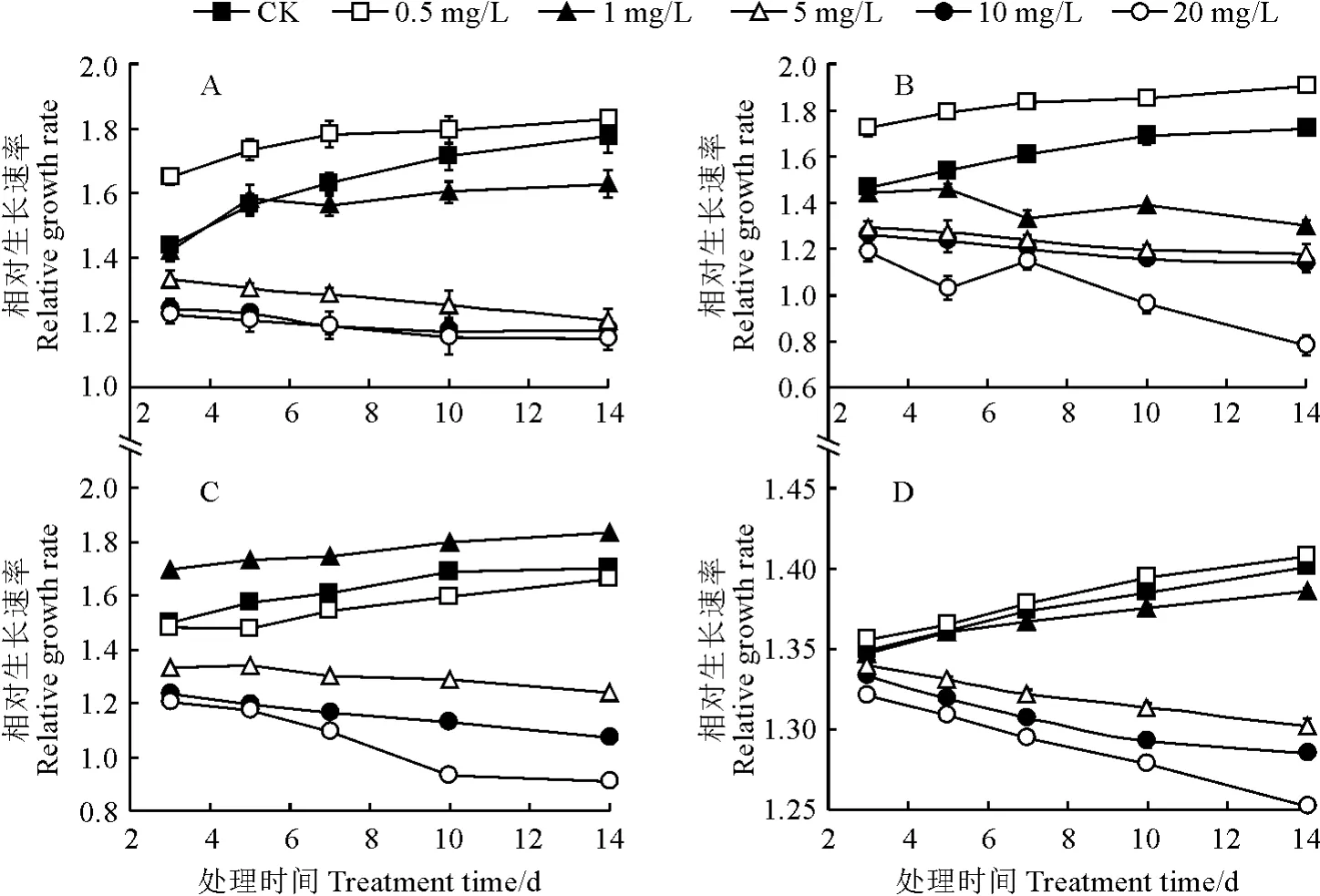
图1 不同浓度铀处理下普通小球藻(A)、黄龙小球藻(B)、蓝藻PCC6803(C)和莱茵衣藻(D)相对生长速率的变化Fig.1 The relative growth rate of chlorella(A),Huanglong chlorella(B),synechocystis PCC6803(C),and C.reinhardtii(D)under different uranium concentrations
其中,普通小球藻和黄龙小球藻的相对生长速率在0.5 mg/L U6+处理下始终显著高于同期对照(P<0.05),在1 mg/L U6+处理下第7天起均显著低于对照,而且黄龙小球藻对铀处理的响应比普通小球藻更敏感,受到促进或者抑制的程度更高,此时基本上停止生长。同时,2种小球藻的生长速率在高浓度铀(5~20 mg/L U6+)处理下始终低于同期对照,均随着处理时间延长逐渐降低,受到显著抑制,且浓度越高受到的抑制作用越大,黄龙小球藻受抑制更严重。可见,铀对2种小球藻生长的影响存在‘低促高抑’的浓度效应,在高于1 mg/L U6+处理下就受到显著抑制,且黄龙小球藻更易受到影响,受影响的程度也更大。
2.2 不同浓度铀藻类光合放氧速率的影响
如图2所示,普通小球藻、黄龙小球藻的光合放氧速率在不同浓度铀处理下的变化趋势相似,即在CK、0.5 mg/L和1.0 mg/L U6+处理下均随着处理时间延长呈上升趋势,而在其余浓度铀处理下却呈逐渐下降趋势;同时,2种小球藻的光合放氧速率在0.5 mg/L U6+处理下均始终显著高于同期CK(P<0.05),受到明显促进,但在5 mg/L 及以上浓度铀处理下均不同程度低于同期CK,均受到显著抑制作用,且随着铀处理浓度的增加抑制程度越显著;但在相同铀浓度处理下,2种藻类之间光合放氧速率无显著差异(P>0.05)。
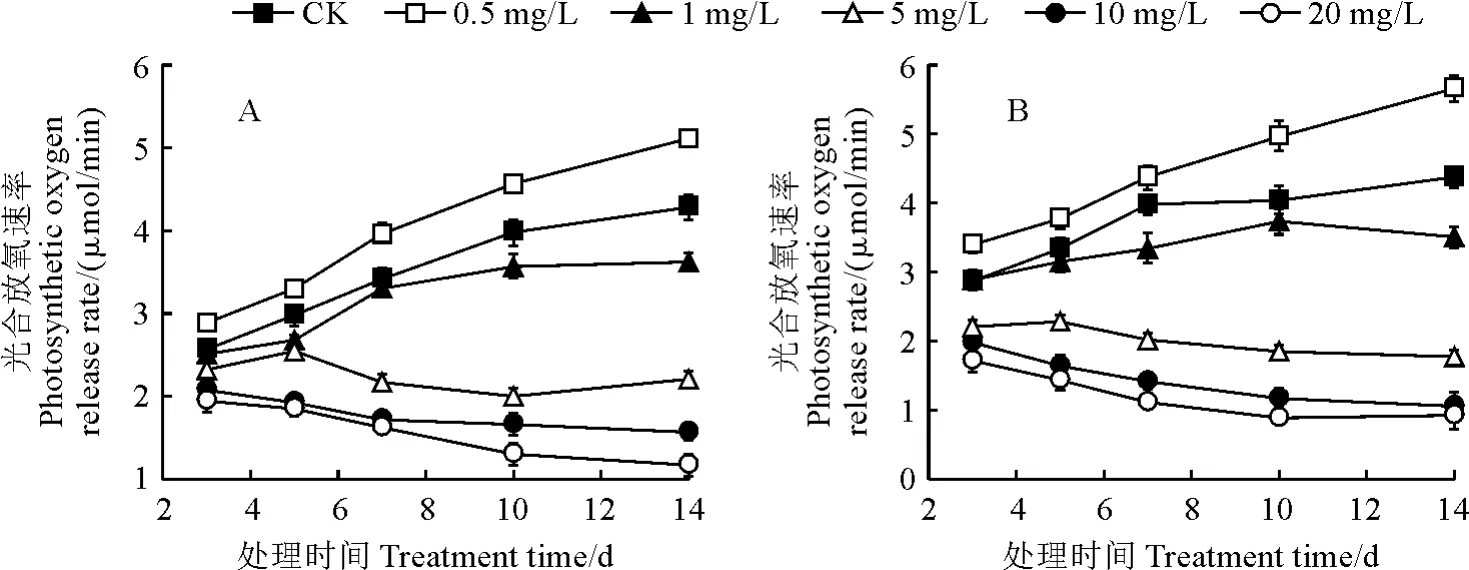
图2 不同浓度铀处理下普通小球藻(A)和黄龙小球藻(B)光合放氧速率的变化Fig.2 The photosynthetic oxygen release rate of chlorella(A)and Huanglong chlorella(B)under different uranium concentrations
2.3 不同浓度铀对藻类叶绿素含量的影响
普通小球藻叶绿素a和叶绿素b含量均随着处理时间增加呈上升趋势,但在各浓度铀处理第14天时均不同程度低于相应对照,且此时各浓度处理叶绿素b含量均与CK 差异显著(图3,A、B)。黄龙小球藻叶绿素a和叶绿素b含量均随着处理时间的增加均呈先升后降的变化趋势(图3,C、D)。其中,黄龙小球藻叶绿素a含量在0.5~5 mg/L U6+处理后3~7 d时比CK 略高,在处理10~14 d时则显著高于CK(P<0.05),但在20 mg/L U6+处理不同时期均显著低于CK(图3,C);黄龙小球藻叶绿素b含量在各浓度铀处理第5天时均显著低于对照,在0.5~5 mg/L U6+处理第10天起均显著高于同期CK(图3,D)。另外,在处理第14天时,2种微藻间叶绿素a含量在低浓度铀(0.5 mg/L U6+)处理下无显著差异(P>0.05),但在高浓度铀(10~20 mg/L)处理下黄龙小球藻叶绿素a含量下降幅度较普通小球藻更大(图3,A、C);在处理第14天时,黄龙小球藻叶绿素b含量在低浓度铀处理(0.5 mg/L U6+)下较普通小球藻上升幅度更大,在高度铀(20 mg/L U6+)处理下较普通小球藻下降幅度更大(图3,B、D)。说明黄龙小球藻对铀胁迫的反应比普通小球藻更敏感,中低浓度(0.5~5 mg/L U6+)铀处理有利于其叶绿素合成。

图3 不同浓度铀处理下普通小球藻(A,B)和黄龙小球藻(C,D)叶绿素含量的变化Fig.3 The chlorophyll content of chlorella(A,B)and Huanglong chlorella(C,D)under different uranium concentrations
2.4 不同浓度铀对藻类叶绿素荧光参数的影响
2.4.1 最大光化学量子产率
铀对2种小球藻光系统Ⅱ最大光化学量子产率(Fv/Fm)值的影响呈明显的双重作用,即在CK、0.5 mg/L U6+处理下均随着处理时间延长呈上升趋势,而在其余浓度铀处理下呈逐渐下降趋势(图4)。
其中,2种普通小球藻的Fv/Fm在0.5 mg/L U6+处理下均显著高于同期CK(P<0.05),并随处理时间增加均呈上升趋势,自处理第7天起,黄龙小球藻上升幅度较普通小球藻更大;2 种小球藻的Fv/Fm在1 mg/L U6+处理下就受到显著影响,并在处理后期均较CK 组显著降低(P<0.05);在5~20 mg/L高浓度铀处理7 d后,2种小球藻的Fv/Fm均较同期CK 显著降低,且降幅随处理时间而增加,黄龙小球藻比普通小球藻下降的幅度更大。
2.4.2 实际光化学量子产率
图5显示,2种小球藻光系统Ⅱ实际光化学量子产率[Y(Ⅱ)]在0.5 mg/L U6+处理下均受到显著促进,并随着铀处理时间延长而增加,但处理第14天时2种藻类Y(Ⅱ)无显著差异(P>0.05);在1 mg/L U6+处理下,普通小球藻Y(Ⅱ)在处理3~10 d时仍显著高于同期CK,但黄龙小球藻Y(Ⅱ)在处理第7天起较CK 显著降低(P<0.05),且随处理时间增加抑制程度越严重,即在低浓度铀处理下,黄龙小球藻Y(Ⅱ)对铀处理比普通小球藻更敏感;2种小球藻的Y(Ⅱ)在5~20 mg/L U6+处理7 d后均较同期CK 显著下降,在处理第14 天时,10~20 mg/L U6+处理黄龙小球藻Y(Ⅱ)受抑制程度比普通小球藻更大。

图5 不同浓度铀处理下普通小球藻(A)和黄龙小球藻(B)Y(Ⅱ)的变化Fig.5 Y(Ⅱ)of chlorella(A)and Huanglong chlorella(B)under different uranium concentrations
2.4.3 相对电子传递速率
从图6可知,随着铀处理时间的增加,不同浓度铀处理普通小球藻、黄龙小球藻的光系统Ⅱ相对电子传递速率(r ETR)的变化趋势与Y(Ⅱ)表现基本一致,均表现为低浓度(0.5 mg/L)促进,高浓度(5~20 mg/L)抑制,且同样在1 mg/L U6+处理7 d后2种微藻r ETR 差异更明显。
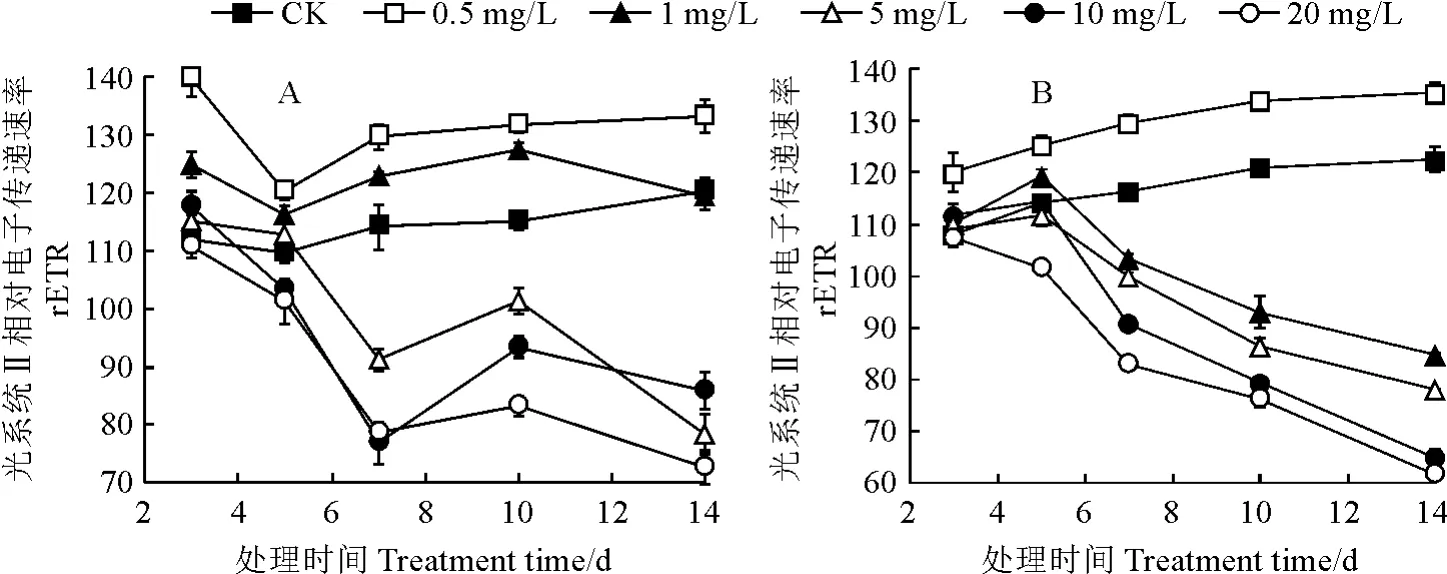
图6 不同浓度铀处理下普通小球藻(A)和黄龙小球藻(B)r ETR 的变化Fig.6 r ETR of chlorella(A)and Huanglong chlorella(B)under different uranium concentrations
2.5 不同参数对铀处理浓度响应差异的回归分析
黄龙小球藻比普通小球藻对铀处理更敏感,相对生长速率在1 mg/L U6+处理下就表现出显著的抑制效应(图1)。此外,黄龙小球藻的光合放氧速率、叶绿素含量、叶绿素荧光参数均在1 mg/L U6+处理下表现出不同程度的抑制效应,因此,选择黄龙小球藻来评价藻类生长及光合参数对铀胁迫浓度的响应差异,为此建立相关回归模型。
由以上试验结果可知,铀浓度对黄龙小球藻7种参数指标均表现为低促高抑,且铀浓度拐点在0~1 mg/L区间,通过0~1 mg/L U6+处理3~14 d的所有试验数据构建回归方程,发现该浓度下数据与二次函数拟合度最高,且存在最大值,根据拟合后7种参数的最大值即可判断各项指标开始下降时的铀浓度,继而找出对铀浓度变化敏感的参数。进一步将试验数据进行显著性分析可得,黄龙小球藻光系统Ⅱ最大光化学量子产率(Fv/Fm)、实际光化学量子产率[Y(Ⅱ)]、相对电子传递速率(r ETR)、相对生长速率(A680)、光合放氧速率、叶绿素a含量均与铀浓度存在极显著关系(P<0.01),叶绿素b含量与铀浓度存在显著关系(P<0.05)。令Fv/Fm、Y(Ⅱ)、r ETR、相对生长速率、光合放氧速率、叶绿素a含量、叶绿素b含量指标分别为y1~y7,铀浓度指标为x1,处理时间为x2,则铀浓度在0~1 mg/L范围,处理3~14 d时与7种参数有显著性回归方程(表1)。

表1 7种参数与铀浓度在0~1 mg/L区间内的回归方程Table 1 Regression equations between seven factors and uranium concentrations in the range of 0-1 mg/L
根据回归方程(表1)可知,光系统Ⅱ最大光化学量子产率(Fv/Fm)、实际光化学量子产率[Y(Ⅱ)]、相对电子传递速率(r ETR)、相对生长速率(A680)、光合放氧速率、叶绿素a含量、叶绿素b含量7种参数到达最大值时铀浓度分别为0.48,0.41,0.41,0.42,0.45,0.43,0.46 mg/L,即大于此浓度时,7种参数指标开始下降;为保证监测的便捷性与时效性,本试验时间维度最大值为14 d,当自变量为时间时,3~14 d范围内7个参数指标均未出现拐点,最大值均出现在第14天。根据不同参数的铀浓度拐点可知,实际光化学量子产率[Y(Ⅱ)]、相对电子传递速率(r ETR)较其他5种参数对铀浓度处理更敏感。
3 讨论
近年来,许多研究表明,盐胁迫及重金属胁迫会显著影响植物的光合作用,且吴惠芳等[19]认为Fv/Fm下降幅度可以作为评价植物抗逆性的参考指标。研究发现,谷子光系统Ⅱ和光系统I光化学量子产量在低钾胁迫下降低[20];萝卜叶片光系统Ⅱ的光化学淬灭系数(qP)和电子传递速率(ETR)在50μmol/L铀胁迫下显著降低[21];燕麦的叶片净光合速率(Pn)和qP在镉胁迫下下降最显著[22];高磷浓度胁迫下,螺旋藻的Fv/Fm随着磷浓度增加呈现先升后降的趋势[23];西瓜幼苗的光合作用在高浓度(180~240 mg/L)镉处理下受显著抑制[24];菠菜叶片光合色素含量和Pn在低浓度(20 mg/L)铀处理下升高,其叶绿素荧光动力学参数在高浓度(50 mg/L以上)铀处理下受到显著影响,其中的最大荧光(Fm)、Fv/Fm和光系统Ⅱ潜在活性(Fv/Fo)均显著降低,而初始荧光(Fo)显著升高,其原因可能是高浓度铀破坏了光系统Ⅱ尤其是反应中心的结构和功能[25]。迄今,关于铀对藻类生长和光合作用影响的研究比较少。杨昊等[26]研究发现,当铀浓度为30 mg/L 时,伊乐藻的光合作用受明显抑制,其Fv/Fm较对照组下降了47.7%。唐东山等[27]报道,满江红鱼腥藻及爪哇伪枝藻的丙二醛含量在30 mg/L铀处理下显著增加,抗氧化酶系统受到破坏,生长严重受阻。本试验结果表明,小球藻的生长和光合作用在低浓度铀(0.5 mg/L)处理下显著促进,在高浓度铀(5~20 mg/L)处理下受到显著抑制,表现为相对生长速率(A680)、光合放氧速率、Fv/Fm、Y(Ⅱ)、r ETR 等指标显著下降。尤其是黄龙小球藻,在1 mg/L 低浓度铀处理下各项指标就表现出明显抑制效应,远远低于先前所报道的高等植物和其他藻类受到抑制的铀浓度,说明小球藻的光合作用对铀处理非常敏感。Fv/Fm反映了光系统Ⅱ的原初光能转换效率,其数值的下降表明微藻正受到光抑制,且光系统Ⅱ反应中心受到破坏或可逆失活;Y(Ⅱ)反映微藻用于光合电子传递的能量占所吸收光能的比例,在5~20 mg/L高浓度铀处理下,微藻的光系统Ⅱ光能转化效率显著降低;r ETR 的变化与光系统Ⅱ电子传递能力有关,其值降低说明铀抑制了光系统Ⅱ电子传递活性。以上这些结果表明,高浓度铀处理可能会导致微藻类囊体上的捕光天线、光系统Ⅱ反应中心、放氧复合体、电子传递体等蛋白的结构域发生变化,从而破坏了光系统Ⅱ,进而抑制其功能[25,28]。以后,铀对藻类生长及光合作用的影响机理仍需要进一步探究。
目前,水体污染监测方法主要为物理化学方法和生物监测方法,物理化学检测方法虽有操作简单的优点,但其存在着监测不全面、不能体现污染物生物毒性的劣势。生物监测方法普遍耗时较长,但有着可用作长期多样化监测、低成本监测、可富集污染物、可评价污染物毒性等特点[29]。现有的基于指示生物、物种多样性变化以及常规生理生化指标变化的水体生物监测指标体系存在周期长,操作繁琐,灵敏性差等缺陷[11],因此筛选超敏感的指示生物和生物标志物,建立新的操作简单、灵敏性高的监测指标体系是现代水体污染风险评价与生物监测技术的前沿发展方向。本试验采用不同浓度铀分别处理2种来自不同生境的普通小球藻和黄龙小球藻,发现黄龙小球藻的相对生长速率、Y(Ⅱ)、r ETR 等指标在1mg/L低浓度铀处理下就可以做出快速灵敏的响应,敏感程度显著大于普通小球藻,更是显著大于已经报道的其他高等植物和藻类,其原因可能是黄龙小球藻生长在高海拔的黄龙地区,其生长条件对水质有着极高要求,相较于其他通过人工繁殖及其他自然环境下生长的藻种,其对水体污染的敏感性有着得天独厚的优势。因此黄龙小球藻具有成为评价水体铀污染风险的超敏感指示生物的潜力。另一方面,本研究表明不同浓度铀处理下,微藻的叶绿素荧光参数Y(Ⅱ)和r ETR 对铀处理的敏感性大于相对生长速率、光合放氧速率、叶绿素含量等指标。因此,叶绿素荧光参数Y(Ⅱ)和r ETR 可以作为对铀污染处理比较敏感的生物标志物。将黄龙小球藻与叶绿素荧光动力学技术结合起来应用于水体铀污染的风险评价与生物监测,可以建立一个新的生态风险评价与生物监测技术体系,具有广阔的应用前景。此外,叶绿素荧光动力学参数很多,在后续的工作中开展进一步系统研究也非常必要。
4 结论
小球藻的生长和光合作用在低浓度铀处理下得到显著促进,而在高浓度铀处理下受到显著抑制。来自于生态脆弱地区的黄龙小球藻比普通小球藻对铀污染更敏感。不同浓度铀处理下,小球藻的光系统Ⅱ实际光化学效率Y(Ⅱ)和相对电子传递速率r ETR的响应比相对生长速率、光合放氧速率、叶绿素含量等参数更敏感。将黄龙小球藻和叶绿素荧光动力学参数Y(Ⅱ)和r ETR 结合起来应用于水体铀污染的生态风险评价,可以提高水体铀污染生物监测的灵敏度。
猜你喜欢
新高考·高三数学(2022年3期)2022-04-28
当代水产(2021年8期)2021-11-04
军事文摘(2020年20期)2020-11-16
内蒙古民族大学学报(社会科学版)(2020年1期)2020-11-03
陶瓷学报(2019年5期)2019-01-12
今日农业(2019年10期)2019-01-04
文苑(2018年22期)2018-11-15
陕西画报(2016年1期)2016-12-01
环境科技(2016年2期)2016-11-08
电源技术(2016年2期)2016-02-27
