
海域环境对阿根廷滑柔鱼雌性个体繁殖力特性的影响
2024-03-06 01:45吴海珠陈昭凯林东明
南方水产科学 2024年1期
吴海珠,陈昭凯,林东明,
1. 上海海洋大学 海洋科学学院,上海 201306
2. 大洋渔业资源可持续开发省部共建教育部重点实验室,上海 201306
3. 国家远洋渔业工程技术研究中心,上海 201306
4. 农业农村部大洋渔业开发重点实验室,上海 201306
繁殖力是指生物个体在繁殖期间所产生的配子数量,也称之为生殖力[1-2]。繁殖力作为物种生存和繁衍能力的重要参考指标,是个体及种群繁殖潜能及补充资源量评估的基础[3],与个体的体型、年龄、食物资源、栖息环境等密切相关[2,4]。头足类是世界海洋渔业的重要经济种类,近十年来其年均渔获量占全球海洋渔业捕捞量的5.2%左右[5]。然而,头足类生命周期短,终生一次产卵,产卵结束后不久便死去[6],资源补充量与亲体的繁殖潜能紧密相关[7]。因此,掌握这些种类的繁殖力特性,是促进其可持续开发利用的重要保证[8]。
阿根廷滑柔鱼 (Illexargentinus) 是典型的大洋性头足类,分布在22°S—54°S 的西南大西洋大陆架和大陆坡海域[9],其寿命在1 年左右,性腺发育成熟至繁殖产卵结束的时长占生命周期的10%~20%[10]。阿根廷滑柔鱼群体结构较为复杂,根据栖息海域、产卵季节、成熟体型等划分为秋冬季产卵的布宜诺斯艾利斯-巴塔哥尼亚北部群体 (Bonaerensis-North Patagonic stock, BNS) 和南部巴塔哥尼亚群体 (South Patagonic stock, SPS),以及夏季产卵群体 (Summer spawning stock, SSS) 和春季产卵群体(Spring spawning stock, SPSS) 4 个群体[11-12]。已有研究表明,阿根廷滑柔鱼的最大潜在繁殖力为75 万粒卵母细胞,实际排卵数量占70%左右[13]。该种类的生殖投入类型为收入-资本混合型,即生殖能量主要来源于性腺发育和配子发生时即时的食物摄食转化,以肌肉存储能量的转化投入为辅[14-15]。2023 年,Zang 等[16]报道阿根廷滑柔鱼成熟配子的能量积累稳定,不受个体摄食营养物质变化的影响。然而,类似于其他头足类,阿根廷滑柔鱼对栖息海域环境变化的敏感性高[17],能量积累及对生殖的投入分配容易受到海洋环境因子波动的影响[18]。Barneche 等[19]研究发现,海洋生物种类的生殖投入决定繁殖力大小,进而影响繁殖产出。本研究将测定分析阿根廷滑柔鱼雌性成熟个体的有效繁殖力、成熟卵子大小及其能量密度,并利用混合模型研究栖息海域环境因子对这三者的效应关系,旨在掌握海洋环境对该种类繁殖力特性的影响,为其资源的科学开发与管理提供理论支持。
1 材料方法
1.1 样本来源
阿根廷滑柔鱼实验样本采集自我国远洋鱿钓作业渔船在西南大西洋海域作业期间的渔获物。采样时间为2021 年2—4 月,采样海域为63.60°W—57.53°W、47.13°S—41.58°S (图1)。样本随机采集后,经冷冻保存运回岸上实验室进行分析。

图1 西南大西洋阿根廷滑柔鱼采样海域和采样站点Fig. 1 Sampling sea area and sampling stations for I. argentinus in southwest Atlantic Ocean
1.2 样品处理与卵径的测量
实验室室温解冻之后,测定阿根廷滑柔鱼样本的胴长 (Mantle length, ML)、体质量 (Body mass,BM)、性别、性腺发育成熟度等。胴长测定精确至1 mm,体质量测定精确至1 g。性腺发育成熟度参照林东明等[20]的标准,划分为I、II、III、IV、V、VI、VII、VIII 等8 个时期,其中I—II 期性腺未发育、III 期生理性发育、IV—V 期生理性成熟、VI 期功能性成熟、VII 期繁殖期、VIII 期繁殖后期 (或衰败期)。性腺发育成熟度判定之后,摘取并称量雌性样本每个性腺组织的质量,包括卵巢、缠卵腺、输卵管、输卵管腺。性腺组织质量测定精确至1 mg。
为分析阿根廷滑柔鱼雌性个体的繁殖力特性,本研究共选取IV—VII 期的雌性样本61 尾 (2 月21 尾,3 月21 尾,4 月19 尾),进行卵巢卵母细胞计数、输卵管成熟卵子计数、成熟卵子大小和成熟卵子能量密度等的分析研究。其中,IV 期17 尾、V 期14 尾、VI 期17 尾、VII 期13 尾 (表1)。卵巢卵母细胞计数、输卵管成熟卵子计数均采用质量计数法,具体测定方法参照林东明等[21],即:1) 卵巢的卵母细胞计数,于卵巢的前、中、后3 个部位分别切取0.3~0.5 g 组织样品,计数每份组织样品卵径>0.4 mm 的卵母细胞数量;2) 输卵管的成熟卵子计数,称取左、右输卵管卵子总质量,于左输卵管和右输卵管中部各挑取0.3~0.5 g 的卵子样品并计数卵子数量。已有研究表明,大部分卵黄早期的卵母细胞 (卵径≥0.40 mm) 都将发育成熟[22],并在繁殖期间成功排放受精[23-24];这部分卵母细胞与输卵管卵子的合计数量可定义为有效繁殖力[2]。对卵巢组织样品或输卵管卵子样品计数之后,进行卵巢卵母细胞数量和输卵管成熟卵子数量的计算,计算公式为:
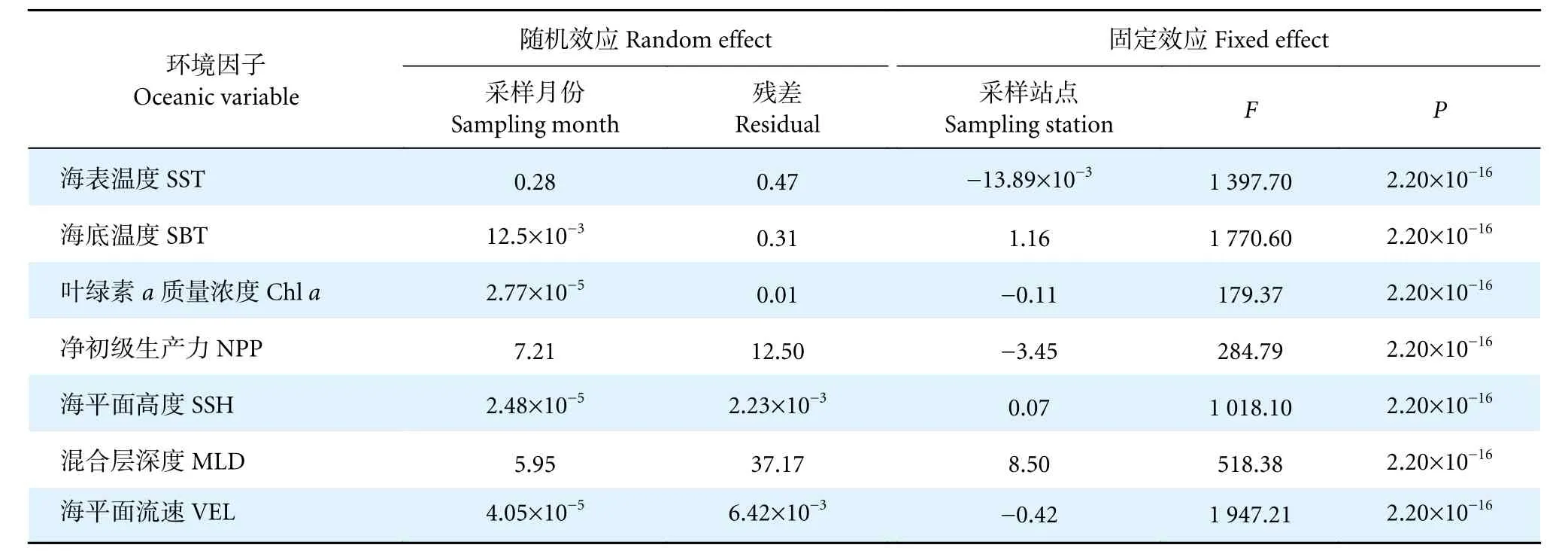
表1 阿根廷滑柔鱼采样站点海洋环境因子与采样站点线性混合效应模型分析结果Table 1 LMM fitting results of oceanic variables and sampling stations for I. argentinus
式中:N为卵巢的卵母细胞数量或输卵管的成熟卵子数量 (粒);m为切取的卵巢组织样品质量或者选取的输卵管部分卵子质量 (mg);e为切取的卵巢组织样品中计数的卵母细胞数量或选取的输卵管部分卵子样品所计数的卵子数量 (粒);M为卵巢总质量或者输卵管卵子总质量 (mg)。
输卵管成熟卵子样品计数后,随机选取80~100 粒卵子置于体式显微镜下拍照,利用Image Pro-Plus 软件测定卵子长径。共测定了8 827 粒成熟卵子长径,测量精度为0.01 mm。
1.3 卵子能量密度
输卵管成熟卵子计数测定之后,将余下的成熟卵子样品于−50 ℃冷冻干燥至恒质量,并称量干质量 (Dry mass, DM),测定精确至0.001 g。随后,将干燥的输卵管卵子样品置于研磨机中研磨粉碎,利用氧弹热量仪测定能量密度值 (kJ·g−1)。
1.4 环境数据下载
已有研究表明,阿根廷滑柔鱼对栖息海域环境的波动变化敏感性高,容易受到水温、海平面高度、海域生产力等的影响[18]。为此,本研究从哥白尼海洋服务 (Copernicus Marine Service;https://marine.copernicus.eu/) 中下载2021 年2—4 月西南大西洋阿根廷滑柔鱼采样海域的环境因子数据,包括海底温度 (Sea bottom temperature, SBT)、净初级生产力 (Net primary production, NPP)、混合层深度(Mix layer depth, MLD)、海表温度 (Sea surface temperature, SST)、叶绿素a质量浓度 (Chlorophyllaconcentration, Chla)、海平面高度 (Sea surface height, SSH)、海表流速 (Sea surface velocity, VEL)。SBT、MLD、SST、SSH、VEL 等环境因子的空间分辨率为0.083°×0.083°,时间分辨率为d;NPP、Chla的空间分辨率为0.25°×0.25°,时间分辨率为d。
1.5 数据分析
阿根廷滑柔鱼采样站点环境因子数据,采用如下匹配处理方法:1) 以采样站点为中心点,排序并筛选距离采样站点最近的4 个环境因子空间位点;2) 匹配采样日期与环境因子的时间,筛选出每个环境因子4 个空间位点的数值;3) 计算每个环境因子在4 个空间位点的数值的平均值,作为采样站点的环境因子数据。利用线性混合效应模型(Linear mixed-effects models, LMM),以采样月份为随机因子,拟合分析每个环境因子的采样站点差异性。
利用单样本Kolmogorov-Smirnoff 分析检验有效繁殖力、卵子大小、能量密度等数据的正态性。若符合正态分布,则利用单因素方差分析 (Oneway ANOVA) 检验各数据分布的月间差异性,并进行Tukey HSD 随后检验;若不符合正态分布,则利用Kruskall-Wallis 非参数检验分析各数据分布的月间差异性,利用Games-Howell HSD 进行随后的显著性检验。
为了掌握海洋环境对阿根廷滑柔鱼雌性个体繁殖力特性的影响,本研究利用广义加性混合效应模型 (Generalized Additive Mixed-Effects Models,GAMM),以采样月份为随机因子,分别拟合雌性成熟个体的有效繁殖力、卵子大小、卵子能量密度与SBT、MLD、SST、SSH、NPP、Chla、VEL 等环境因子的关系。多重共线性分析显示,SBT、NPP 和MLD 存在多重共线性关系 (vif>10)。因此,GAMM 模型中的环境因子为SST、SSH、Chla和VEL。
所有数据分析在R 平台[25]进行。LMM 和GAMM 模型分别采用 R 平台中的lmerTest 和mgcv 数据包进行处理。统计分析的显著性水平为P<0.05,结果用“平均值±标准差”表示。
2 结果
2.1 栖息海域环境因子
2021 年2—4 月,阿根廷滑柔鱼样本采集区域的海域环境因子热点图分布见图2。其中,采样站点的海表温度为12.31~17.37 ℃,平均值为 (14.74±1.97) ℃ (图2-a);海底温度为4.60~7.81 ℃,平均值为 (6.45±1.12) ℃ (图2-b);海平面高度为−0.17~0.05 m,平均值为 (−0.06±0.05) m (图2-c);混合层深度为10.37~43.80 m,平均值为 (21.13±8.73) m(图2-d);叶绿素a质量浓度为0.37~0.62 mg·m−3,平均值为 (0.42±0.05) mg·m−3(图2-e);净初级生产力为11.76~25.11 mg·m−3·d−1,平均值为 (18.36±5.34) mg·m−3·d−1(图2-f);海表流速为0.04~0.45 m·s−1,平均值为 (0.10±0.07) mg·m−3·d−1(图2-g)。LMM 模型结果显示,这些环境因子在不同采样站点之间均存在显著性差异 (P<0.05,表1)。
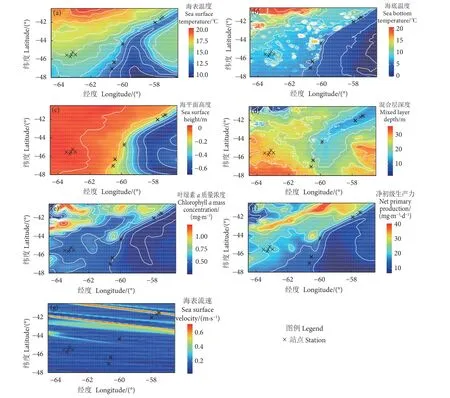
图2 阿根廷滑柔鱼采样海域环境因子分布Fig. 2 Distribution of oceanic variables in sampling sea area of I. argentinus
2.2 有效繁殖力及与环境因子关系
阿根廷滑柔鱼雌性个体的有效繁殖力为22 996~161 797 粒卵母细胞,平均值为67 061.72±24 147.00;60%雌性个体的有效繁殖力在40 000~80 000 粒卵母细胞之间 (图3-a)。在不同月份之间,雌性个体的有效繁殖力无显著性差异 (F=0.47,P=0.50) (图3-b)。
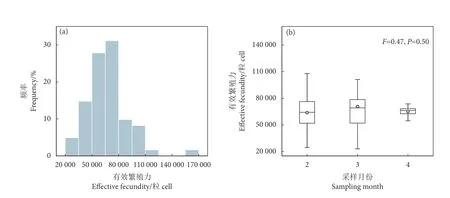
图3 阿根廷滑柔鱼雌性成熟个体有效繁殖力频率分布及其月份分布Fig. 3 Frequency distribution and monthly distribution of effective fecundity for mature female I. argentinus
GAMM 模型结果显示,阿根廷滑柔鱼雌性个体的有效繁殖力与叶绿素a质量浓度呈显著相关关系 (F=5.57,P<0.05),而与海表温度 (F=0.74,P>0.05)、海平面高度 (F=0.16,P>0.05) 和海表流速(F=2.35,P>0.05) 等无显著效应关系 (表2)。随叶绿素a质量浓度的增加,阿根廷滑柔鱼有效繁殖力呈先升高后降低的变化趋势,在叶绿素a质量浓度为0.40~0.45 mg·m−3时达到较大值 (图4)。
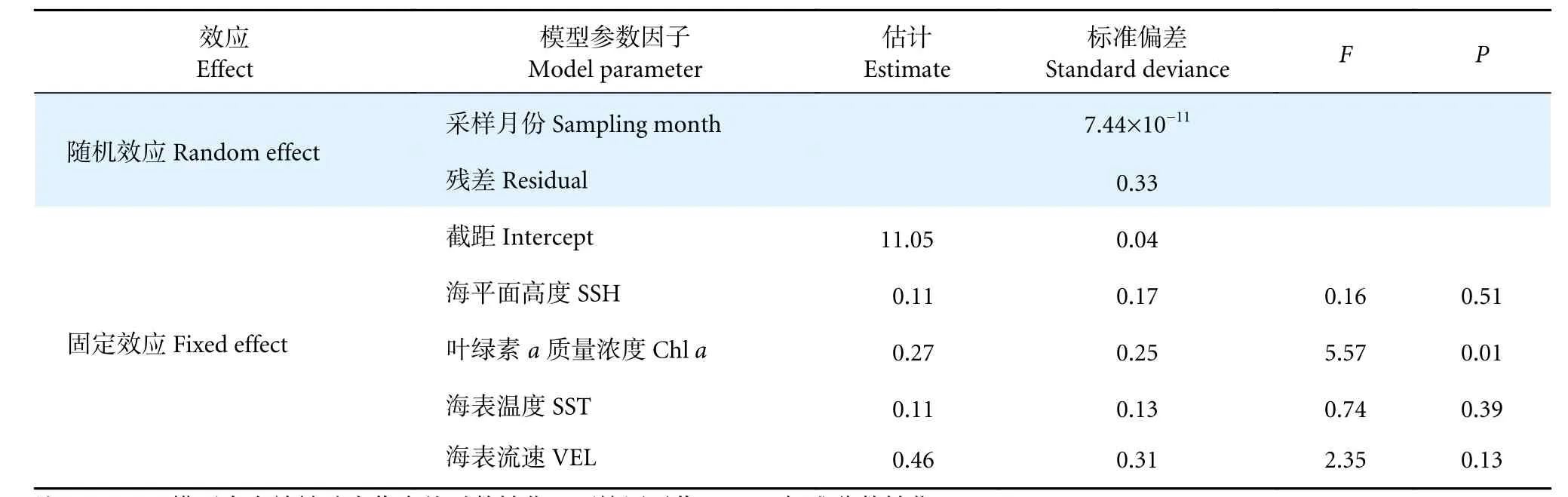
表2 阿根廷滑柔鱼雌性个体有效繁殖力与环境因子的GAMM 模型拟合结果Table 2 GAMM model fitting results of effective fecundity and oceanic variables for female I. argentinus

图4 阿根廷滑柔鱼雌性成熟个体有效繁殖力与叶绿素a 质量浓度的关系Fig. 4 Relationship between effective fecundity and chlorophyll a mass concentration for mature female I. argentinus
2.3 成熟卵子大小及与环境因子关系
阿根廷滑柔鱼输卵管成熟卵子长径为1.11~1.89 mm,平均值为 (1.41±0.23) mm,但是存在两个峰值区间,分别为1.1~1.2 mm 和1.5~1.6 mm(图5-a)。显著性检验结果显示,阿根廷滑柔鱼成熟卵子长径存在显著的采样月间差异 (F=72.05,P=9.36×10−12),2 月成熟卵子长径最大,平均为(1.60±0.22) mm (图5-b)。
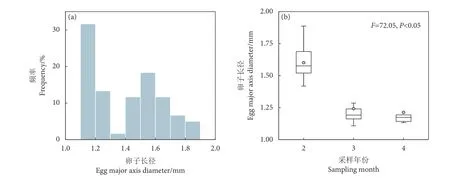
图5 阿根廷滑柔鱼雌性成熟个体输卵管成熟卵子长径频率分布及不同月份间的大小分布Fig. 5 Frequency distribution of oviduct egg diameter and their size distribution in different months for mature female I. argentinus
阿根廷滑柔鱼成熟卵子长径与叶绿素a质量浓度 (F=17.65,P<0.05)、海表温度 (F=27.23,P<0.05)、海平面高度 (F=9.91,P<0.05) 等呈显著相关关系,而与海表流速无显著效应关系 (F=2.38,P>0.05) (表3)。其中,成熟卵子长径在海表温度低于14 ℃时变化平缓,之后随着海表温度升高呈增大趋势 (图6-a);随着叶绿素a质量浓度增加,成熟卵子长径也呈增大趋势,在叶绿素a质量浓度达约0.50 mg·m−3时,卵子大小达到较大值 (图6-b);成熟卵子长径与海平面高度成正相关关系,随着海平面高度的增加而增大 (图6-c)。
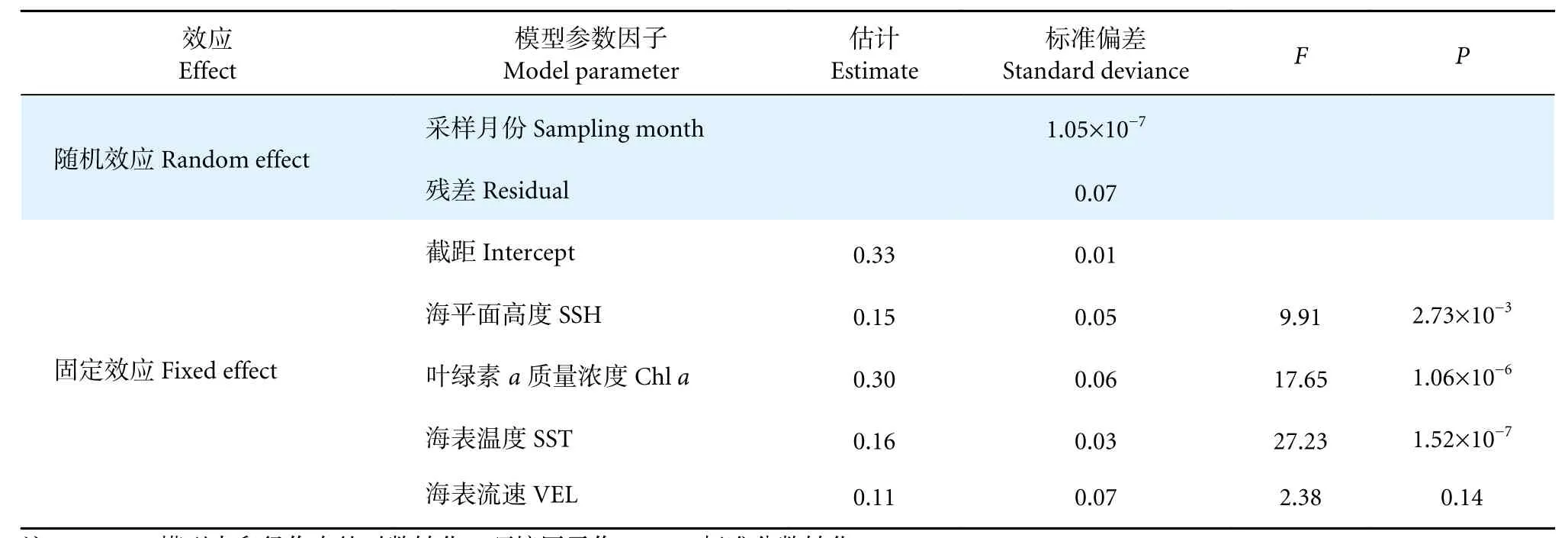
表3 阿根廷滑柔鱼雌性个体卵径与环境因子的GAMM 模型拟合结果Table 3 GAMM model fitting results of oviduct egg diameter and oceanic variables for mature female I. argentinus
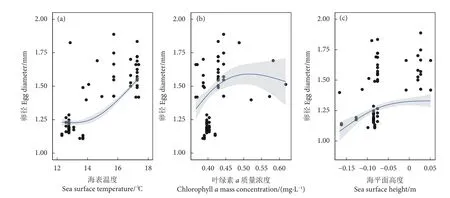
图6 阿根廷滑柔鱼雌性成熟个体输卵管卵子大小与海表温度、叶绿素 a 质量浓度、海平面高度的关系Fig. 6 Relationships between oviduct egg diameter and sea surface temperature, chlorophyll a mass concentration,sea surface height for mature female I. argentinus
2.4 卵子能量密度及与环境因子关系
输卵管卵子的能量密度为22.80~26.21 kJ·g−1,平均值为 (24.95±0.93) kJ·g−1(图7-a)。不同采样月份之间,输卵管卵子的能量密度值差异显著(F=67.66,P<0.05);其中2 月的卵子能量密度值最大 ,平均值为 (25.72±0.26) kJ·g−1;随着采样月份的推移,卵子能量密度呈下降趋势,但是3 和4 月间的卵子能量密度无显著性差异 (Tukey HSD,P>0.05) (图7-b)。

图7 阿根廷滑柔鱼雌性成熟个体卵子能量密度频率分布及其采样月份分布Fig. 7 Egg energy density frequency distribution and its distribution by sampling months for mature female I. argentinus
GAMM 模型结果显示,阿根廷滑柔鱼雌性成熟个体的输卵管卵子能量密度与海表温度 (F=14.95,P<0.05)、叶绿素a质量浓度 (F=7.62,P<0.05)、海平面高度 (F=14.88,P<0.05) 三者均呈显著的正效应关系,与海表流速无相关性 (F=2.48,P>0.05) (表4)。随着海表温度升高、叶绿素a质量浓度增大、海平面高度升高,成熟卵子能量密度均呈增大趋势 (图8)。

表4 阿根廷滑柔鱼雌性成熟个体卵子能量密度与环境因子的GAMM 模型拟合结果Table 4 GAMM model fitting results of oviduct egg energy density and oceanic variables for mature female I. argentinus
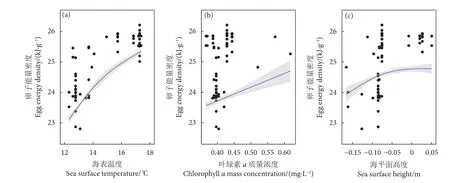
图8 阿根廷滑柔鱼雌性成熟个体卵子能量密度与海表温度、叶绿素a 质量浓度、海平面高度的关系Fig. 8 Relationships between oviduct egg energy density and sea surface temperature, chlorophyll a mass concentration,sea surface height for mature female I. argentinus
3 讨论
3.1 海洋环境对繁殖力大小的影响
繁殖力作为种群繁衍的基础,一定程度上决定着物种后代补充量的大小[26-27]。头足类种群结构复杂,往往存在不同的地理种群,且种群之间繁殖力差异明显[13]。不同地理群体之间的繁殖力差异是对栖息水域的选择适应过程[28-29],也是物种遗传和海域环境共同作用的结果[3]。通常,栖息水域以外海或公海海域为主的柔鱼类的最大潜在繁殖力可达百万粒卵母细胞[30],栖息在沿岸或港湾海域为主的乌贼类的潜在繁殖力在5 000 粒卵母细胞以内[31],而生活在珊瑚礁区域的银磷乌贼 (Sepioteuthissepioidea) 仅有2~6 粒卵母细胞[32]。已有研究表明,阿根廷滑柔鱼不同群体的繁殖力差异显著,以秋冬季产卵的南巴塔哥尼亚群的繁殖力最大,可达24.6 万粒卵母细胞[33],夏季产卵群体的繁殖力为2.5 万~13.5 万粒卵母细胞[21]。本研究中,阿根廷滑柔鱼雌性成熟个体的有效繁殖力介于2.3 万~16.2万粒卵母细胞,与夏季产卵群体的繁殖力相近[21]。同时,阿根廷滑柔鱼雌性成熟个体的有效繁殖力在2—4 月间无显著性差异。结合采样海域及采样月份,本研究的阿根廷滑柔鱼雌性成熟个体可能主要来自夏季产卵群体,并有南部巴塔哥尼亚群的个体。
此外,生物个体繁殖力作为种群繁殖力的重要指标,能够反映出繁衍后代的能力及对环境波动变化的响应能力[4]。有研究显示,头足类繁殖力大小容易受到栖息海域生产力波动变化的影响[6]。在海洋环境中,叶绿素a质量浓度较高表征浮游植物生长繁盛,可为生物提供丰富的饵料基础[34-35],进而影响栖息海洋生物个体的时空分布及其繁殖价值[36]。本研究显示,阿根廷滑柔鱼雌性个体的有效繁殖力与栖息海域的叶绿素a质量浓度显著相关,且随叶绿素a质量浓度增大呈增加趋势,而与海表温度、海平面高度和海表流速等环境因子无显著效应关系。可见,栖息海域初级生产力是影响阿根廷滑柔鱼繁殖力的关键环境因子。同时,海域初级生产力是影响生物个体生长发育及其体型的关键因素[37],而体型与个体的繁殖潜能呈线性相关[2]。因此,阿根廷滑柔鱼繁殖力与叶绿素a质量浓度的显著效应关系进一步佐证了头足类繁殖力大小容易受到栖息水域生产力的影响。而且,这种效应关系也可推测阿根廷滑柔鱼在叶绿素a质量浓度较高的海域具有较好的摄食情况以促进个体生长,提高了繁殖价值。
3.2 海洋环境对卵子大小及能量积累的影响
海洋生物的卵子大小容易受到较多生物及生态因素的影响,如物种的繁殖产卵批次、繁殖力大小、产卵策略、栖息海域环境等[13,38]。有研究表明,卵子大小是海洋生物对生境变化的适应性结果,与后代成活率密切相关[39]。本研究发现,阿根廷滑柔鱼雌性成熟个体的成熟卵子长径平均值为1.41 mm,与其他大洋性柔鱼类的卵径大小相近[2]。但是,这些种类均有很大的繁殖力,如茎柔鱼的繁殖力高达320 万粒卵母细胞,卵径则只有1.2 mm 左右[40]。这可能是大洋性柔鱼类对多变的大洋性海洋环境的一种生殖选择策略,通过产生卵径较小、数量较多的配子以维持种群数量稳定[13]。与此相反,生活于近海底栖的乌贼类繁殖力较低、卵子长径也较大[41],与相对稳定的近海底层水域环境条件密切相关[42]。可见,头足类的卵子大小是对其栖息环境的选择适应。在本研究中,阿根廷滑柔鱼雌性个体的成熟卵子大小与栖息海域的叶绿素a质量浓度、海表温度、海平面高度等均呈显著的效应关系,与头足类配子大小与栖息海域环境密切相关的研究结果相一致[41,43]。然而,本研究发现阿根廷滑柔鱼成熟卵子大小分布存在两个峰值区间,且来自2 月的样本卵径显著大于3 和4 月。在西南大西洋,2 月是海域生产力较高、水域温度相对适宜的季节,可为海域内的生物种类提供丰富的饵料生物资源,促进其生长发育[34]。因此,海域生产力、水温的季节差异可能是引起阿根廷滑柔鱼成熟卵子存在两个卵径峰值区间分布的原因。具体原因仍需要后续多个年份实验数据的验证。
通常,生物个体的卵径越大其卵黄物质积累也越多,可提高胚胎发育的成功率和孵化仔鱼的成活率[44]。本研究发现,阿根廷滑柔鱼成熟卵子能量密度也在2 月达到最大值,与该月卵径最大的情况一致,说明在2 月亲体对单配子的能量投入相对较高。与其他头足类相似,阿根廷滑柔鱼具有较强的环境适应能力[38],在生长和配子发生上可以适时优化能量积累及分配[45],以促进卵母细胞的生长发育[31]。已有研究表明,叶绿素a质量浓度对头足类性腺组织能量密度可产生正效应关系[18],栖息海域水温可直接影响海洋生物个体对营养物质的摄取水平及个体代谢率[46];海平面高度则是许多环境因素的综合作用,如海洋环流、海面风速、上升流等,对营养物质的聚集起着关键作用,为捕食者提供丰富的饵料生物,促进组织的能量积累[47]。在本研究中,阿根廷滑柔鱼雌性个体成熟卵子的能量密度与海表温度、叶绿素a质量浓度和海平面高度均呈显著的正相关关系,也进一步说明了该种类在生殖投入上对栖息海域环境的选择适应特征。此外,3 和4 月相对较低的卵子能量密度也可说明阿根廷滑柔鱼能量积累对配子发生的投入分配是一个连续的过程[22],该过程与栖息海域的环境波动变化密切相关。然而,Zang 等[16]报道阿根廷滑柔鱼成熟卵子具有稳定的营养物质组成,则成熟卵子大小应该也是相对恒定的。为此,后续仍需要增加对卵子大小及能量积累分析的采样时间序列及海域范围,以深入探讨造成卵子大小及能量投入分配月间差异的具体原因。
4 结论
本研究通过对阿根廷滑柔鱼雌性成熟个体繁殖力特性进行分析,同时应用广义加性混合模型分析了繁殖力特性与环境因子之间的效应关系,发现繁殖力大小与叶绿素a质量浓度显著相关,且叶绿素a质量浓度、海表温度、海平面高度对成熟卵子大小及其能量密度产生了显著性影响。这些结果表明,海域生产力及相对适宜的物理环境对阿根廷滑柔鱼繁殖力特性有显著的影响作用。然而,西南大西洋的大陆架宽广、洋流系统显著,且容易受到大陆径流的补充影响,海域中的水温、盐度、溶解氧、营养盐等存在较大的季节性变化,随之影响海域生产力及其物理环境[34]。因此,后续仍需通过延长采样时间及增大采样海域范围,进一步开展阿根廷滑柔鱼繁殖力特性与栖息海域环境的关系研究,以深入探讨该种类繁殖的环境选择适应特性。
猜你喜欢
海洋渔业(2021年1期)2021-03-18
水生生物学报(2021年1期)2021-02-04
海洋信息技术与应用(2020年2期)2020-07-27
热带海洋学报(2020年3期)2020-05-25
海洋通报(2020年6期)2020-03-19
成都信息工程大学学报(2019年5期)2019-05-21
猪业科学(2018年4期)2018-05-19
祝您健康·文摘版(2018年3期)2018-04-06
婚育与健康(2015年12期)2015-07-06
恋爱婚姻家庭·养生版(2015年10期)2015-05-14
