
仿刺参南移养殖温度耐受性相关基因的差异表达分析
2024-03-05 07:00:04朱志煌吴建绍杨求华
渔业研究 2024年1期
朱志煌,林 琪,吴建绍,杨求华
(福建省水产研究所,福建省海洋生物增养殖与高值化利用重点实验室,福建 厦门 361013)
仿刺参(Apostichopusjaponicus)是棘皮动物门海参纲刺参科动物的统称,具有较高的营养、药用和经济价值。仿刺参属于温带种类,自然分布区域北起俄罗斯远东地区,经朝鲜、韩国、日本,南至中国北部,虽然在福建的自然海区无分布,但其所具有的巨大经济价值推动该品种养殖业从北往南扩张[1]。自2005年以来,福建省从辽宁等地引进仿刺参,并大力开展“北参南养”实验。2022年福建海参产量达到45 636t,占全国总产量的18.36%,稳居全国第三位。
仿刺参为浅海底栖生物。水温是影响仿刺参生长、生理性能和免疫反应等功能的重要环境因素,并且在水温升高到一定范围后,其立即进入摄食停止、活动减少的“夏眠”状态。福建地区夏季的高温会导致养殖仿刺参发生大规模死亡,这给养殖业造成巨大的损失。目前,缺乏仿刺参耐高温良种是制约该产业进一步发展的重要因素。选择育种是获得水生动物新品系的重要方法[2-4]。研究证明,热胁迫刺激能够一定程度地提高动物耐高温的能力[5-7]。热休克蛋白(Heat shock proteins,HSPs)和小热休克蛋白(Small heat shock proteins,sHSPs)作为细胞内的分子伴侣,能够防止蛋白质在细胞内聚集,提高细胞的耐热能力[8-13]。以往研究表明,HSPs是诱导耐热的主要介质,其表达模式与生物体的耐热水平直接相关[14-18]。此外,致死必需蛋白 [Protein lethal (2)essential for life,Protein l(2)efl]基因也是一种小热休克蛋白的同源基因,其表现出与sHSPs类似的功能,主要与生物的耐热能力有关,并在一定程度上保护细胞免于凋亡。HSP26、gp96、HSP70、HSP90a和proteinl(2)efl基因均属于热休克蛋白家族的成员,其表达的水平可用于评估高温诱导下仿刺参的热耐受力。
目前,国内外对北方仿刺参的研究较为系统,众多学者在组织形态学、生态学、生理学、免疫学及分子生物学等方面展开大量研究并取得了丰硕的研究成果[19-22]。有研究指出,在高温胁迫下,仿刺参的HSP70、proteinl(2)efl、HSP90和HSP26等热休克蛋白基因的表达量会上调[23-24]。针对北方仿刺参夏眠前、后展开转录组测序的分析研究,发现温度波动会对其热休克蛋白的表达产生显著的影响[25]。还有研究表明,仿刺参的耐热性是可塑的,预先的热冲击实验能够提高其耐热能力[26-27]。因此,将北方仿刺参转移到南方来进行养殖适应,能在一定程度上提高其耐热性能。
本研究从山东青岛购入仿刺参幼苗,将其饲养于温度较高的福建漳州户外浅池塘中,使其经历长期的高温海水刺激,以此作为南移福建养殖的仿刺参实验组,并以北方相同苗龄的仿刺参作为对照组,比较两组间的抗热性能,以期为仿刺参耐高温品系的培育提供理论参考。
在道路桥梁路线设计阶段,设计单位应该综合自身掌握的地形地质条件、周边自然环境资料等对工程投入运营后可能会出现的问题进行分析预测,在此基础上制定科学合理的维护养护和检测维护计划,促进道路桥梁工程使用寿命的提升。
1 材料与方法
1.1 实验动物和驯化
2016年冬季从山东省青岛市即墨养殖场(36°30′43.5″N、120°50′59.0″E)购买仿刺参幼苗,并将其分至两地养殖,一部分放在原养殖场养殖,另一部分转移到福建省漳州市东山养殖场室外饲养浅池塘进行养殖(23°34′34.1″N、117°23′28.8″E),两地养殖的仿刺参均在自然光周期和水温下饲养一年。海水经砂滤过滤,盐度约为30,以流水模式进行养殖。仿刺参幼苗每天投喂饲料一次,每次喂足人工海参饲料(金派生物,中国威海),并定期对池塘吸污。同时,每天记录水温情况,每月测定仿刺参幼苗的生长情况。2017年冬季,分别将青岛市即墨养殖场和漳州市东山养殖场养殖的仿刺参运至厦门市进行实验,此时仿刺参均为20月龄(600日龄)。为了方便后续区分,将两地运回的仿刺参分别简易命名为青岛本地群体(Qingdao local sea cucumber populations,Qls)和南方养殖群体(South aquaculture sea cucumbers populations,Sas)。实验开始前,将两地运来的仿刺参分别移入温度为16 °C的养殖水槽中,暂养14 d,每天换掉1/3~1/2的等温海水,其他饲养条件均相同。
1.2 实验设计
1.2.1 仿刺参温度耐受范围实验
为研究给定处理时间的仿刺参温度耐受范围,急性温度实验设计的刺激温度超过仿刺参养殖过程中海水的自然温度。将不同仿刺参群体分别放入预先调好海水温度的40 L透明塑料桶,每个塑料桶放入10头仿刺参,每个实验温度设置3个重复。将两个群体的仿刺参分别同时放入水温为28、29、30、31、32、33、34 °C的塑料桶中,经过2 h的温度刺激后,转移至16 °C养殖水槽中暂养7 d,观察记录其存活情况,以确定两个仿刺参群体的亚致死温度、致死温度和半致死温度。实验水温由温控器控制调节,实验期间每2 min用精度为±0.2 °C的水银温度计监测水温情况。在实验期间,仿刺参的饲养条件与驯化期间相似。
1.2.2 仿刺参温度耐受性相关基因的差异表达实验
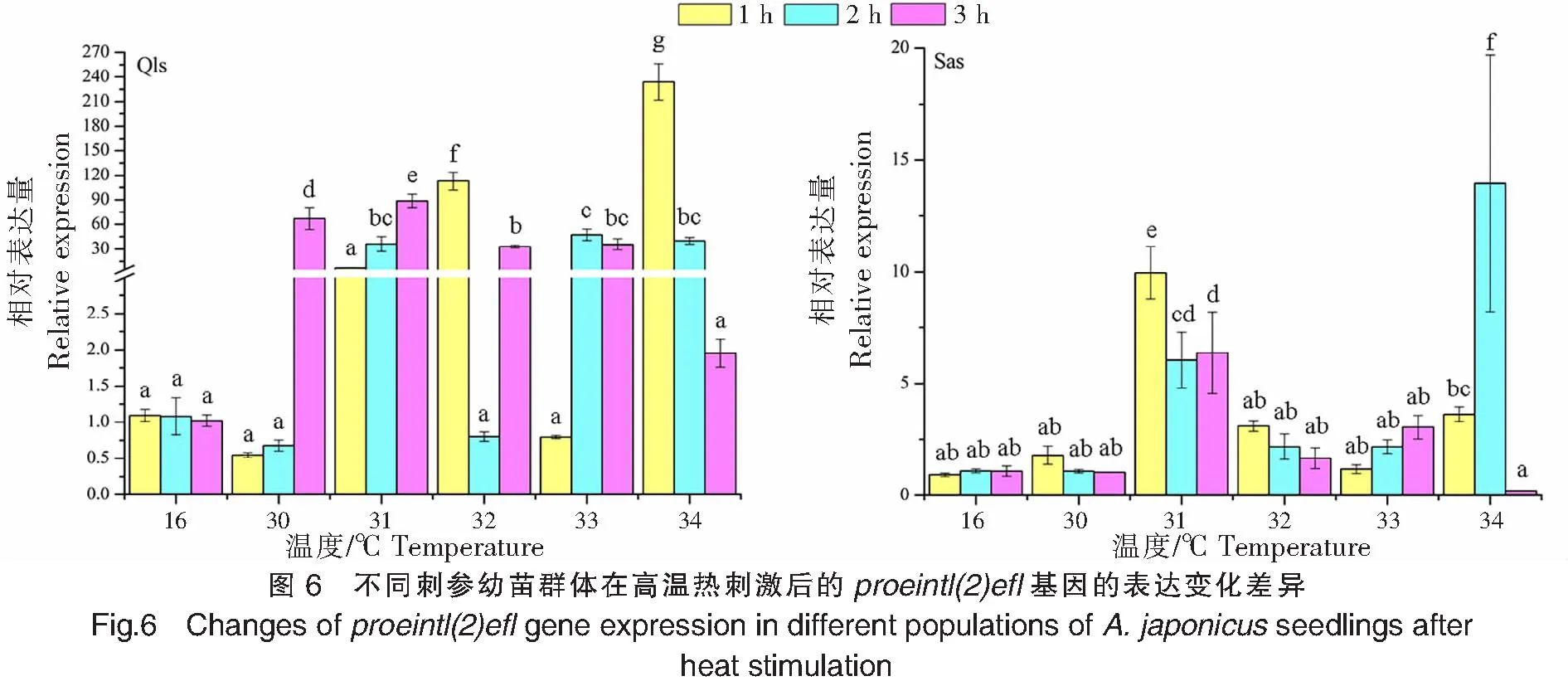
不同刺参幼苗群体在高温热刺激后的proeintl(2)efl基因的表达变化差异如图6所示。青岛本地群体经过30、31 ℃的高温刺激后,proeintl(2)efl基因的表达量峰值出现在3 h处,分别为(67.230±13.512)、(88.897±8.242);在32、34 ℃高温刺激下,proeintl(2)efl基因的表达量峰值均出现在1 h处,表达量的最高值出现在34 ℃刺激1 h处(234.167±22.130);而在33 ℃高温刺激下,proeintl(2)efl基因的表达量峰值出现在高温刺激2 h处。南方养殖群体经过30、31、32℃高温刺激后,proeintl(2)efl基因的表达峰值均出现在1 h处;而33、34 ℃高温刺激下,proeintl(2)efl基因的表达量峰值则分别出现在3、2 h处,表达量的最高值出现在34 ℃刺激2 h处(13.948±5.740)。青岛本地群体在高温刺激1、2、3 h后,proeintl(2)efl基因的表达峰值分别出现在34、33、31 ℃;而南方养殖群体则分别出现在31、34、31℃。
1.3 RNA提取和cDNA的合成
将整头仿刺参置于装有液氮的研钵中碾碎。取约80 mg样品,放入1 mL Trizol试剂(Qiagen)中进行匀浆,再使用 Invitrogen 公司的 Trizol 试剂盒,按照说明书提取样本组织的总RNA。利用QuantScript RT Kit,按照说明书合成cDNA,提取的样品保存在-80 ℃冰箱中,备用待测。
1.4 引物设计和荧光定量实验
根据NCBI(http://www.ncbi.nlm.nih.gov/Genbank/)网站上已公布的HSP90a、HSP26、HSP70、gp96和proeintl(2)efl基因为模板设计荧光定量引物(表1)。采用荧光定量 PCR的方法,检测5个温度耐受性相关基因在温度刺激后的不同仿刺参幼苗群体中的表达情况。以β-actin的表达量为标准值,将对照组和实验组个体在不同时间点的表达量进行校正,以消除不同个体对实验结果的影响[26-27]。再将这些校正后的表达量与对照组(16 °C)进行比较,分析各基因在对照组和实验组中的倍数变化,从而发现基因在高温刺激后的表达规律。荧光定量PCR的反应条件为95 ℃预变性10 s、40个循环中95 ℃变性5 s、60 ℃退火34 s。
绘画审美上,中国画追求诗意的传达。苏轼认为王维“诗中有画,画中有诗”,这从本质上体现出诗意精神是中国画的内在特质之一。清代画家恽南田追求简淡清远的审美意趣,体现在绘画上就是“画以简贵为尚,简之入微,则洗尽尘滓,独存孤迥,烟鬟翠黛敛容而退矣。”[12]画家的审美意趣会在绘画中有充分的展现。所以,在中国画教学上,应注重培养学生的审美境界,提升其绘画的追求品味。
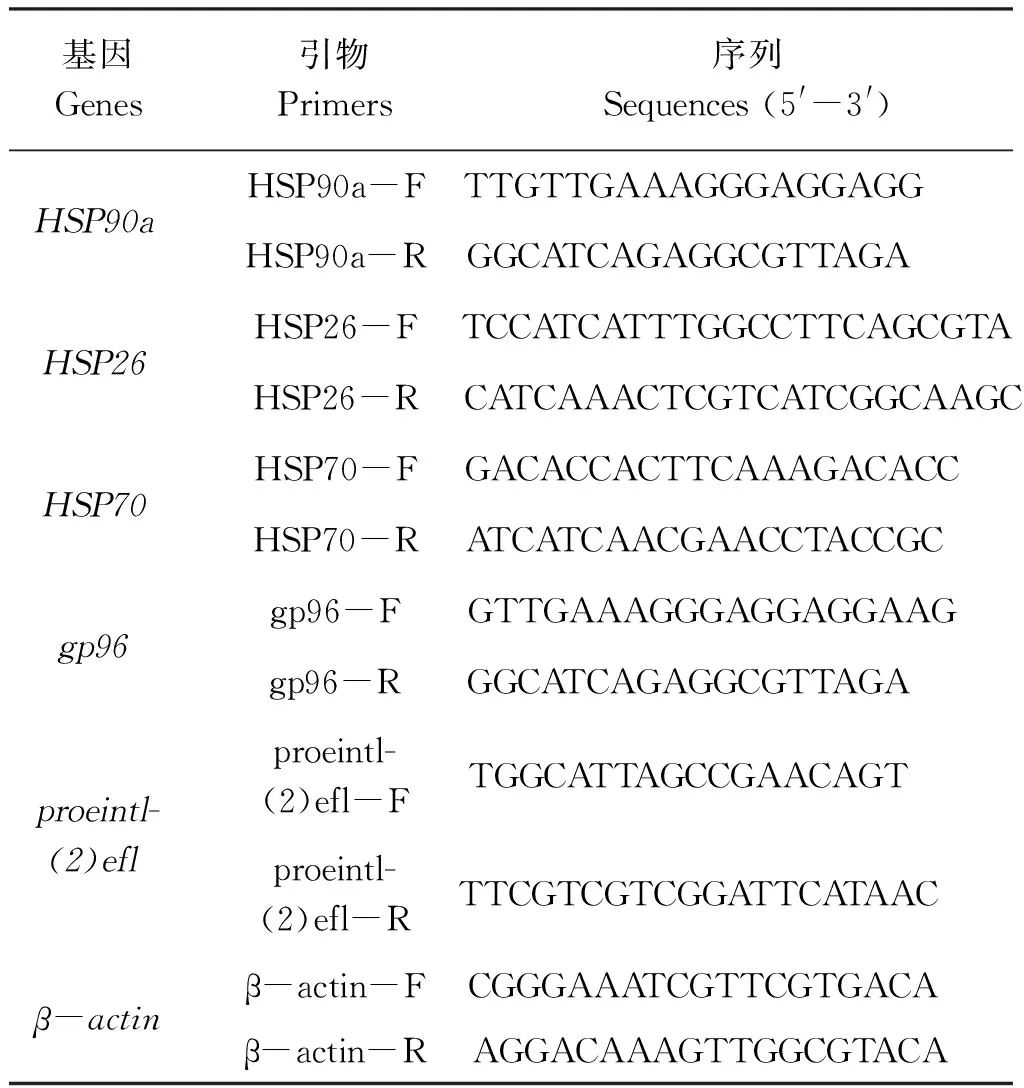
表1 本研究中所使用的引物Tab.1 The primer used in the study
1.5 统计学分析
实验数据采用SPSS 21.0软件进行统计分析,采用单因素方差分析(One-way ANOVA),以最小显著极差法(LSD)比较数据间的显著性[28-29]。数据结果以平均值±标准差表示,P<0.05为差异显著。半致死温度通过SPSS 21.0软件中的概率单位 (Probit)回归法[30-32]计算得出。
青岛本地群体和南方养殖群体在高温刺激后的存活率如图1所示。青岛本地群体在28~30 ℃高温刺激2 h后均没有出现死亡,而南方养殖群体在28~32 ℃高温刺激2 h后没有出现死亡,但两个群体在经过34 ℃高温刺激2 h后均全部死亡。在31~33 ℃高温刺激组中,南方养殖群体的存活率均高于青岛本地群体,其中青岛本地群体在31、32、33 ℃温度下刺激2 h后的存活率分别为76.7%±5.8%、50.0%±0.0%、13.3%±5.8%,而南方养殖群体分别为100.0%±0.0%、100.0%±0.0%、56.7%±5.8%,两个群体的存活率差异显著(P<0.05)。因此,高温刺激2 h后,青岛本地群体和南方养殖群体的亚致死温度分别为30、32 ℃,半致死温度(ULTs50)分别为31.9 ℃ (31.4~32.4 ℃)和33.1 ℃ (32.6~33.5 ℃),两个群体的致死温度均为34 ℃(图1)。
2 结果与分析
2.1 不同仿刺参幼苗群体的温度耐受性
因此,虽然中国专利法的第69条(一)条款在文义解释上能够支持这种区别适用者的理论,但这种解释会导致该条的目的落空,不应该固执地坚持。
不同仿刺参幼苗群体在高温热刺激后的HSP26基因的表达变化差异如图5所示。青岛本地群体经过30 ℃高温刺激,HSP26基因的表达量在1 h处出现明显的降低,然后在2 h处继续降低,但在3 h处达到峰值(34.787±4.978);在31 ℃高温刺激下,HSP26基因的表达量在1 h处出现显著上升,之后随着刺激时间的延长而继续上调;在32 ℃高温刺激下,HSP26基因的表达量在1 h处达到高峰,在2 h处降低到正常水平,之后在3 h处又出现升高;在33 ℃高温刺激下,HSP26基因的表达量在1 h处出现显著下调,之后在2 h处出现大幅增加并达到峰值,之后在3 h处出现迅速降低;在34 ℃高温刺激下,HSP26基因的表达量在1 h处迅速上调并达到该群体表达量的最高值(65.127±4.054),之后逐渐降低到正常水平。南方养殖群体的HSP26基因的表达量在30、31 ℃高温刺激下出现显著的上调,但是表达量峰值不同,最高值出现在30 ℃刺激1 h处,为(93.710±5.674);在33 ℃高温刺激下,HSP26基因的表达量随着刺激时间的延长而增加,在3 h处出现表达量峰值;HSP26基因的表达在32、34 ℃高温刺激下的表达模式相近,即高温刺激1 h处的表达没有显著差异,而在2 h处的表达量出现快速上调并达到最高峰,之后恢复正常水平。青岛本地群体在高温刺激1、2、3 h后,HSP26基因的表达峰值分别出现在34、33、30 ℃;而南方养殖群体则分别出现在30、34、31 ℃。
2.2 HSP90a基因的表达情况
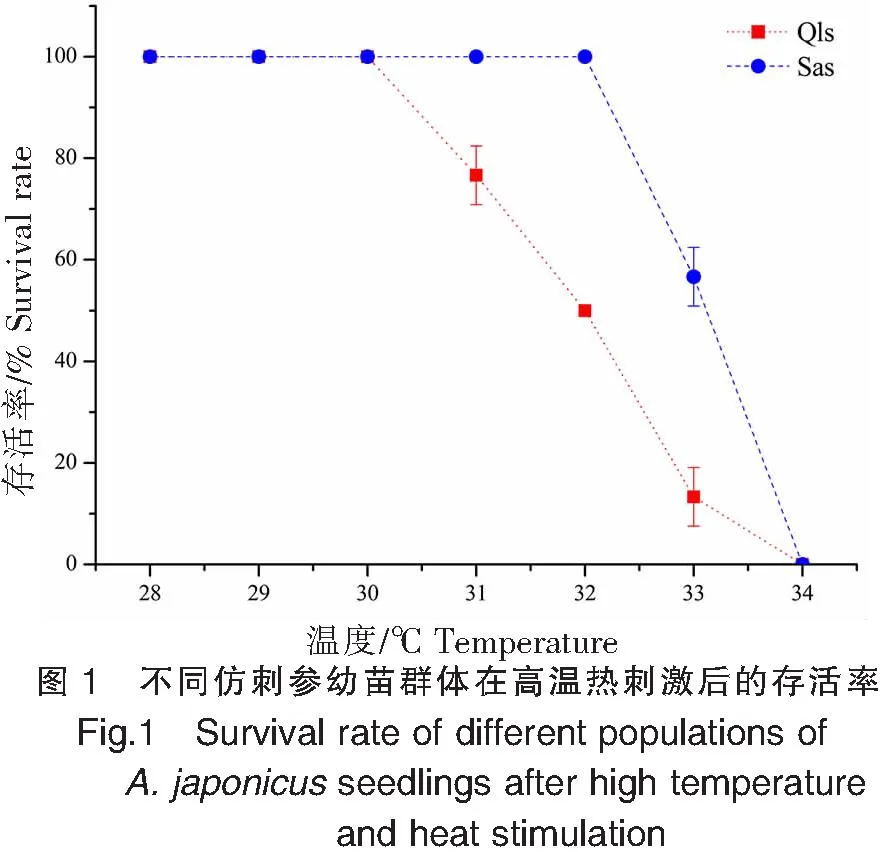
不同仿刺参幼苗群体在高温热刺激后的HSP90a基因的表达变化差异如图2所示。青岛本地群体经30、31、32、33 ℃高温刺激后,HSP90a基因的表达量峰值出现在温度刺激3 h处,分别为(9.801±1.303)、(24.133±2.232)、(21.683±3.082)、(25.473±3.402);而在34 ℃高温刺激后,HSP90a基因的表达量高峰发生在1 h处,此时达到青岛本地群体表达量的最高值(43.367±3.137),随后表达量随着温度刺激时间的延迟而出现降低。南方养殖群体经30 ℃高温刺激1 h后,HSP90a基因的表达量已达到(42.000±8.798);在30、31、33 ℃高温刺激下,HSP90a基因的表达峰值出现在温度刺激3 h处,而32 ℃高温刺激的最高表达值出现在2 h,此时达到南方养殖群体表达量的最高值(222.150±4.980)。在34 ℃的高温刺激下,南方养殖群体和青岛本地群体的表达模式相近,HSP90a基因的表达高峰均发生在1 h处。青岛本地群体在高温刺激1、2、3 h后,HSP90a基因的表达峰值分别出现在34、34、33 ℃;而南方养殖群体则分别出现在32、32、30 ℃。

注:标有不同小写字母表示差异显著(P<0.05),标有相同小写字母表示差异不显著(P>0.05)。以下同此。Notes:Different lowercase letters indicated significant difference (P<0.05),while the same lowercase letter meant no significant difference (P>0.05).The same as below.
2.3 gp96基因的表达情况
不同仿刺参幼苗群体在高温热刺激后的gp96基因的表达变化差异如图3所示。青岛本地群体在30 ℃高温刺激下1、2 h处,gp96基因的表达量显著低于对照组(16 ℃),而在刺激3 h后,表达量才上调并达到峰值(2.508±0.910);在31 ℃高温刺激下,gp96基因的表达量在1、2、3 h处均出现显著上调,并在3 h处出现表达量峰值;在32 ℃高温刺激下,gp96基因的表达量在温度刺激1 h后出现大幅增加,并达到峰值,随后在2、3 h处表达量分别出现明显的下调和上调;在33 ℃高温刺激下,gp96基因的表达量随着高温刺激时间的延长而增加,在3 h处表达量达到青岛本地群体的最高值(353.833±21.586);在34 ℃高温刺激下,gp96基因的表达量随着温度刺激时间的延长而降低。
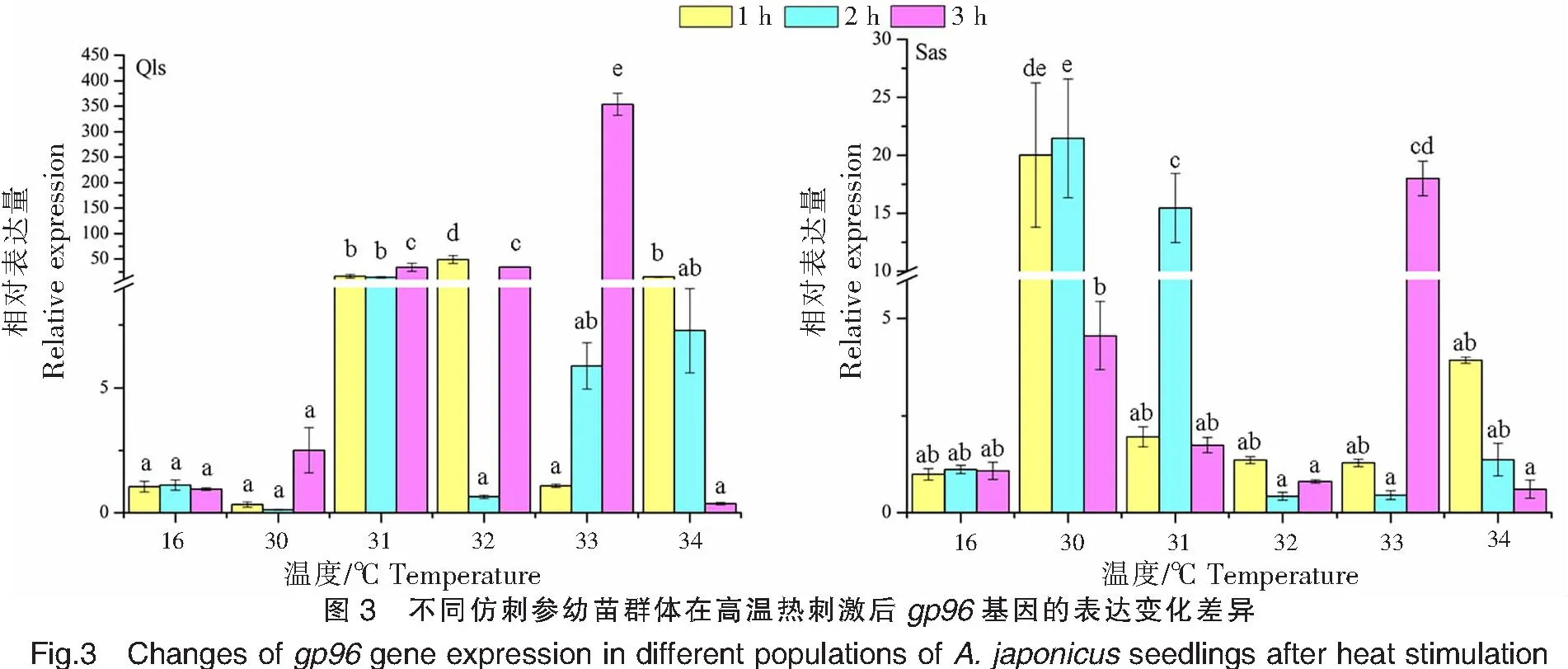
青岛本地群体在高温刺激1、2、3 h后,gp96基因的表达量峰值分别出现在32、31、33 ℃;而南方养殖群体则分别出现在30、30、33 ℃。
南方养殖群体在30 ℃高温刺激1 h后,gp96基因的表达量已达到(20.019±6.224),并在2 h处达到峰值,此时也是南方养殖群体表达量的最高值(21.467±5.131),最后在3 h处出现下降;在31 ℃高温刺激下,gp96基因的表达模式与30 ℃高温刺激下的相同,呈现先升高后下降的趋势;在32 ℃高温刺激下,gp96基因的表达量在1 h处出现上调,但在2~3 h间出现上下浮动;在33 ℃高温刺激下,gp96基因的表达量在1~3 h间来回波动,最高和最低表达值分别出现在2、3 h处;在34 ℃高温刺激下,gp96基因的表达量在1 h处达到最高,之后表达量随着高温刺激时间的延长而逐渐降低。
综上所述,老年衰弱患者进行手术存在发生短期和长期不良结局的高风险,术后不良事件发生率较非衰弱患者显著增加。有必要将衰弱纳入老年患者的术前评估中,以明确是否存在衰弱及老年衰弱患者术中可能出现的问题,并以此为据进行个体化围手术期管理。全过程需要跨学科团队协作。目标是手术可以使老年衰弱患者获益,减少术后不良并发症和失能,最终能够回归社区生活,改善生活质量。
2.4 HSP70基因的表达情况
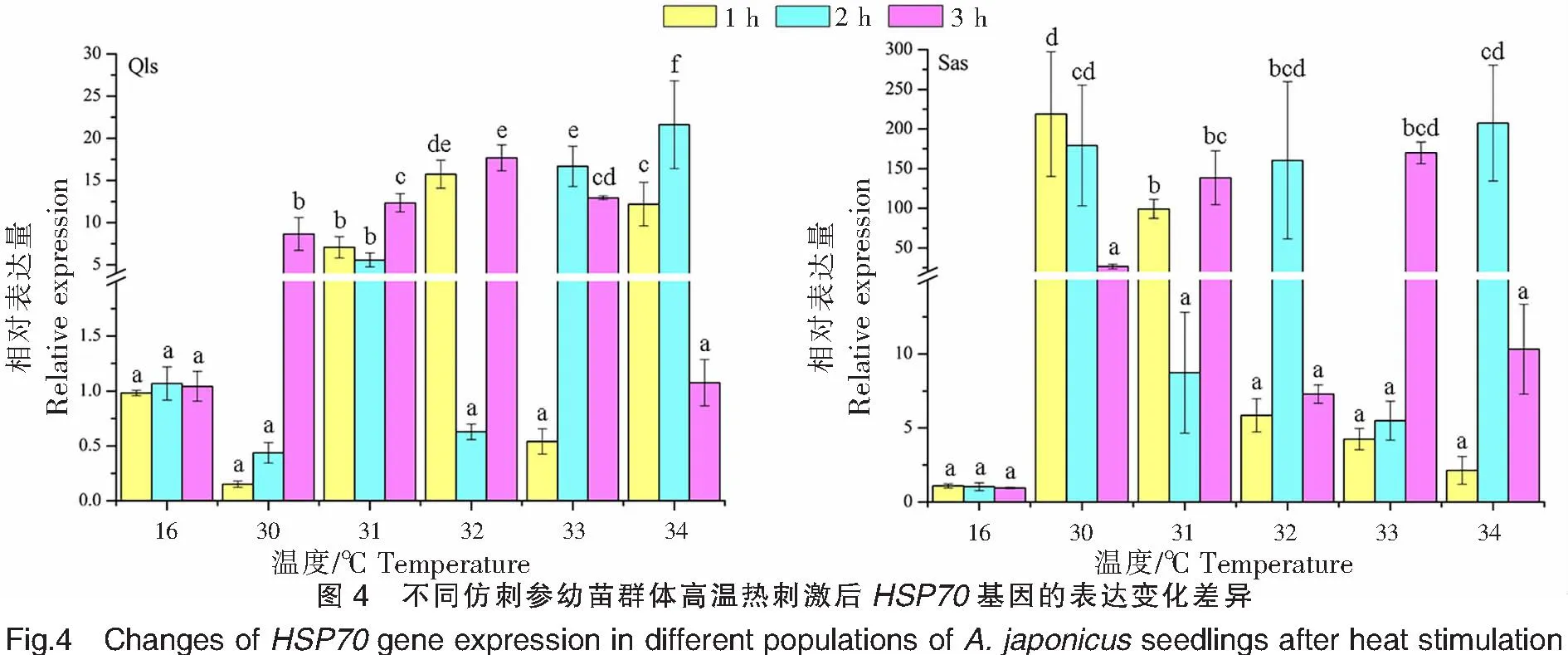
不同仿刺参幼苗群体在高温热刺激后的HSP70基因的表达变化差异如图4所示,在相同的温度刺激下,不同刺激时间的HSP70基因的表达存在差异。青岛本地群体在30、31、32 ℃高温刺激下,HSP70基因的表达量均在3 h处达到峰值,分别为(8.649±1.936)、(12.350±1.076)、(17.680±1.532);而在33、34 ℃高温刺激下,HSP70基因的表达量峰值均出现在2 h处;表达量的最高值出现在34 ℃刺激2 h处,为(21.610±5.208)。南方养殖群体在30 ℃高温刺激下,HSP70基因的表达量在1 h处达到峰值,也是该群体表达量的最高值,为(218.750±78.701);而在31、33 ℃高温刺激下,HSP70基因的表达量峰值均出现在3 h处;在32、34 ℃高温刺激下,HSP70基因的表达量峰值均出现在2 h处。青岛本地群体在高温刺激1、2、3 h后,HSP70基因的表达量峰值分别出现在32、34、32 ℃;而南方养殖群体则分别出现在30、34、33 ℃。
2.5 HSP26基因的表达情况
控制变量。经济发展水平用剔除价格因素的实际人均GDP衡量,地区经济发展水平越高越有利于该地集聚资源,有利于提升企业的创新水平,促进产业升级[16];人力资本水平用各地区平均受教育年限来测量,计算公式为:(小学受教育人数×6+初中受教育人数×9+高中受教育人数×12+大学受教育人数×16)/总受教育人数[17], 人力资本是影响环境规制和技术进步的重要因素;产业结构的计算公式是工业增加值/GDP,产业结构的变动直接推动产业升级变化;技术创新用各年各省的专利授权量表示[18],新技术的发明能够促使企业的生产经营向高新技术方向转变,从而使产业结构向更高端的方向调整,实现产业升级。

2.6 proeintl(2)efl基因的表达情况
为了解不同诱导温度和处理时间下两个仿刺参群体HSPs、sHSPs和proeintl(2)efl基因的表达模式差异,将两个群体分别转入选定的温度中进行热刺激,温度设置为16 (对照组)、30、31、32、33、34 °C,刺激时间分别为1、2、3 h。每个刺激温度和刺激时间均有10头仿刺参,在刺激1、2、3 h后,分别随机采集不同实验组别中的3头仿刺参,然后立即置于液氮中,带回实验室并保存在-80 °C冰箱中,备用待测。
3 讨论
3.1 高温驯化提高仿刺参的耐热性
仿刺参是典型的温带物种,对外界环境变化具有强敏感性。Meng X等[27]和王青林[32]对北方仿刺参进行高温驯化,结果发现高温驯化后仿刺参的半致死温度为31.8 ℃。本研究将北方仿刺参转移至南方,在自然水温下进行为期一年的养殖,然后与在北方养殖的同批次仿刺参进行耐热性比较,分析了两个群体间的耐热性差异,其中温度耐受性实验得出两个群体的致死温度均为34 ℃;南方养殖群体的亚致死温度和半致死温度分别为32、33.1 ℃,青岛本地群体分别为30、31.9 ℃。本研究发现的青岛本地群体半数致死温度与Meng X等和王青林的结论基本一致。李晓英等[33]比较了北方5个不同地理群体仿刺参的高温耐受力,结果发现胶南群体的半致死温度最高(31.9 ℃),而昌黎群体半致死温度最低(31.5 ℃),分析认为胶南群体分布靠南、纬度最低,经过长时间较高温度的驯化与适应,使其对于高温的耐受力最强,而昌黎群体分布位于较北端、纬度较高,长期处于略微低温驯化中,导致其对高温的耐受能力较弱。本研究发现,南方养殖群体的亚致死温度和半致死温度显著高于青岛本地群体,这与李晓英等[33]的研究一致,说明生长环境中水温的高低能够影响仿刺参的温度耐受性,而受到长期高温驯化的仿刺参具有更强的高温耐受性。养殖在南方自然水温中的仿刺参幼苗比北方苗种经历更强的高温胁迫,因此其相较于北方苗种具有更高的耐热性。今后在仿刺参品种选育工作中可以通过在南方累代定向选育,不断提高其耐热性能,从而迭代选育出耐高温品系。
那个时候,我们的县里,早就有了洋学堂了。小学好几个,大学没有。只有一个男子中学,往往成为谈论的目标。谈论这个,不单是翠姨,外祖母、姑姑、姐姐之类,都愿意讲究这当地中学的学生。因为他们一切洋化,穿着裤子,把裤腿卷起来一寸,一张口,格得毛宁外国话,他们彼此一说话就“答答答”,听说这是什么俄国话。而更奇怪的就是他们见了女人不怕羞。这一点,大家都批评说是不如从前了,从前的书生,一见了女人脸就红。
3.2 南北不同养殖仿刺参群体的热休克蛋白基因的表达差异
当生物体受到外界环境刺激时,包括热休克蛋白在内的一些基因的表达可能被诱导。根据平均分子质量大小,热休克蛋白可分为HSP110、HSP100、HSP90a、HSP70、HSP60和sHSP[15,34-35]。本研究发现,经过高温热刺激后,南方养殖群体和青岛本地群体的HSP90a、gp96、HSP70、HSP26和proeintl(2)efl基因的表达存在差异。经过30 ℃高温刺激的青岛本地群体的HSP90a、gp96、HSP70、HSP26基因的表达量在1、2 h处没有出现显著增加,在3 h处才出现明显上调,而南方养殖群体的基因表达量在30 ℃高温刺激1 h处均出现显著上调。结果表明经过高温热刺激后,南方养殖群体的温度耐受性相关基因的上调时间明显早于青岛本地群体,说明南方养殖群体对高温热刺激的适应能力强于青岛本地群体。
在不同的刺激温度下,青岛本地群体和南方养殖群体的HSP90a、gp96、HSP70、HSP26和proeintl(2)efl基因的表达量峰值发生在不同的高温热刺激时间点,青岛本地群体的表达峰值分别出现在34 °C 1 h处、33 °C 3 h处、34 °C 2 h处、34 °C 1 h处和34 °C 1 h处,而南方养殖群体则分别出现在32 °C 2 h处、30 °C 2 h处、30 °C 1 h处、30 °C 1 h处和34 °C 2 h处。随着热刺激温度的升高或时间的延长,基因表达并非一直上调,这种下降与高温下生物体整体蛋白质合成的减少有关[36]。当外界温度达到生物体的耐受极限,随着温度的进一步升高,机体热损伤蛋白质会不断增加,总蛋白质合成遭到破坏,因此热休克蛋白的表达受到抑制。
对比南方养殖群体和青岛本地群体,HSP90a基因在南方养殖群体的表达量最高值(为对照组的222倍)是青岛本地群体(为对照组的43倍)的5倍,HSP70基因在南方养殖群体的表达量最高值(为对照组的218倍)是青岛本地群体(为对照组的21倍)的10倍,这与王青林[32]研究发现的仿刺参高温驯化组HSP70基因在热刺激诱导温度下的最高表达量明显高于低温驯化组的结果一致,说明经过高温驯化的仿刺参能保持较高水平的HSP70表达,以防止蛋白质变性和聚集。 此外,HSP90a、gp96、HSP70、HSP26和proeintl(2)efl这5个基因在两个仿刺参群体的34 ℃高温刺激3 h处的表达量均下调至对照组水平,说明此时仿刺参HSPs基因的表达已达到上限并开始迅速回落,其已无法通过继续提高HSPs基因的表达水平来抵抗外界的热胁迫,这与本研究两个群体的致死温度均为34 ℃的结果一致。
(1)基于 OMI的汾渭平原 SO2柱浓度值季节变化特征比较明显,由高到低依次为冬季、秋季、春季和夏季。太原、渭南和西安逐月浓度变化总体呈现下降趋势,临汾表现出了弱上升趋势。这4个城市的SO2柱浓度值与地面监测站的SO2浓度值具有较好的相关性,说明基于 OMI的 SO2柱浓度变化一定程度上可反映大气SO2浓度变化。
4 结论
经过高温驯化,仿刺参能够更早地对外界高温做出反应,同时其具有更强的合成热休克蛋白的能力,以避免温度骤升引起的蛋白质变性和聚集,从而提高它们在夏季高温下的耐热能力和存活率。因此,通过将仿刺参幼苗进行“北参南养”,经过几代的高温选育,能够为仿刺参耐高温品系的选育提供良好的实验材料。
猜你喜欢
红蜻蜓·低年级(2021年12期)2022-01-19 05:18:32
红蜻蜓·低年级(2021年12期)2021-12-19 15:06:23
大连海洋大学学报(2020年2期)2020-05-06 02:26:10
福建林业科技(2019年1期)2019-04-10 03:15:44
金桥(2018年7期)2018-09-25 02:28:14
商周刊(2018年10期)2018-06-06 03:04:10
江苏农业科学(2017年10期)2017-07-21 13:45:32
知识经济·中国直销(2016年5期)2016-11-07 09:34:56
江苏农业科学(2015年7期)2015-08-20 11:59:23
——An Idea From "Etudes Metro"—the Work of Pierre Schaeffer
大众文艺(2014年4期)2014-01-28 14:52:00
