
三峡蓄水以来葛洲坝下中华鲟产卵场河床质特征变化规律
2024-03-02 06:38刘文成李君轶李鹏程吴金明辉沈刘志刚
水生生物学报 2024年4期
刘文成 李君轶 李鹏程 班 璇 吴金明 张 辉沈 丽 刘志刚 杜 浩
(1. 上海海洋大学海洋科学学院, 上海 201306; 2. 中国水产科学研究院长江水产研究所农业农村部淡水生物多样性保护重点实验室, 武汉 430223; 3. 中国科学院精密测量科学与技术创新研究院环境与灾害监测评估湖北省重点实验室, 武汉 430077)
对众多河流生物来说, 河床质是其栖息地重要物理组成部分[1]。了解并研究河床质性状及变化情况是保护河流生物的重要前提, 尤其是对水体底层生活及产粘性卵的鱼类, 产卵河床底质是其种群延续的关键因子。水声学技术最早被用来探测潜艇和测量水深, 近期用于探测鱼群较多[2]。与通常的评估方法相比, 水声学方法具有高效、调查区域广、不损坏生物资源、提供可持续的数据等优点[3]。作为水声学方法主要工具的回声探测仪在其中起着关键作用, 回声探测仪发展至今, 目前主要应用有评估鱼类资源量、监测鱼类行为、鉴别鱼类种类、评估其他水生生物、监测水质和水生生态系统、探测水深和底质类别等[4]。
由于水下的特性, 河床质通常使用回声探测仪进行研究, 不同底质类型对声波反射特性有差异,回声探测仪可凭借这一特征来观测河床底质类型以及底部沉淀物[5]。声学特性包括第一个底部回波的第一部分和第二部分被用来区分不同硬度、粗糙度的沉积物[6]。Heald和Pace[7]提供了两个回波的能量特征与海底参数之间关系的理论背景。此后Magorrian等[8]、Kaiser等[9]、Bax等[10]相继使用该声学技术绘制海底生物群落的生物资源。Daniel等[11]利用当时较新的Biosonics回声探测系统对巴德河的整个底质生境进行了描述, 提取粗糙度和硬度数据, 并与底质粒径制定底质分类散点图;Anderson等[12]发现回声探测系统获取的特征参数与底质的化学元素含量有直接关系, 并且探测频率不同, 研究结果可靠性也不同。基于水声学方法探测底质的特点, 结合一定的传统取样或光学观察,能够迅速且可靠地对河床沉积物进行分类[3]。
中华鲟(Acipenser sinensis)是国家一级重点保护水生动物, 国际自然保护联盟(IUCN)极危级(CR)物种, 国际濒危动植物种贸易公约(CITES)附录Ⅱ保护物种[13]。在20世纪后期, 由于水利工程建设、过度捕捞、水体污染和航运等诸多因素, 其野生种群资源量不断下降[14]。据2017—2020年进行的中华鲟自然繁殖生态调查报告, 葛洲坝下近坝江段已连续几年未观测到中华鲟的自然繁殖活动[15],这可能与该区域的非生物环境变化有较大关系。在目前中华鲟历史产卵场无法发挥作用, 而葛洲坝下现有产卵场中华鲟自然繁殖活动持续中断的情况下, 中华鲟自然繁殖所需的非生物环境变化研究亟须开展。
中华鲟历史产卵场河床质特征常被描述为“卵石河床质”, 中国科技工作者认为底质必须具备岩石或卵石中华鲟才会开始产卵[16]。另外河床质粒径、组成和布局等都会对自然繁殖产生影响, 一般认为清洁的河床质对于多数鲟鱼类的自然产卵有利[17]。常剑波和曹文宣[18]认为, 刺激中华鲟产卵的外界因素主要是河床的底质状况。因此众多学者对葛洲坝下中华鲟产卵场的底部环境进行研究, 杜浩等[19]在2008年通过水下视频观测根据卵石大小、排列方式及是否含沙等状况, 将河床质表层结构划分为6种类型。杜浩等[20,21]系统观察了中华鲟受精卵的分布状态, 提出由细砂决定的河床质填塞度是决定中华鲟自然繁殖发生位点和繁殖效果的重要因素。在国内使用回声探测仪, 主要是对葛洲坝下中华鲟产卵场底部环境的硬度和粗糙度进行研究。张辉[3]首次研究了葛洲坝下中华鲟产卵场底质表观特征和声学特征的相关性, 2014年结合水下视频获得的研究资料确定了该江段的底质特征关系式[22]。杜浩等[21]研究了2004—2012年葛洲坝下中华鲟产卵场河床质的粗糙度和硬度变化, 探究三峡大坝蓄水运行对河床质的影响。蔺丹清[23]在2016年对整个长江湖北宜昌中华鲟自然保护区进行了声学采样, 确定了相应底质特征的关系式, 与之前相比有一定的区别。虽然部分学者提出了葛洲坝下中华鲟产卵场底质声学特征和表观特征的相关公式, 但其判别底质效果较差, 原因可能在于未使用与Biosonic探测系统配套的声学后处理系统, 使用的声学处理软件未对回波进行特殊处理。
水利工程的建设运行、人类活动及长江流域的洪涝灾害等均会导致河床质及其构建的微生境变化, 从而影响中华鲟自然繁殖行为和早期仔幼鱼的存活, 这直接关系到野生中华鲟种群的补充群体数量。因此有必要对葛洲坝下中华鲟产卵场的河床质进行更深入的研究, 结合水声学方法和水下视频技术, 重新构建硬度、粗糙度与底质表观特征的相关性, 补充分形维数与底质的相关性, 并据此对葛洲坝下中华鲟产卵场的底质演变过程进行描述,结合长江流域发生的水沙事件揭示底质变化规律,并解释中华鲟自然繁殖状况的变化。这有助于推测未来葛洲坝下中华鲟产卵场的变化趋势, 为将来中华鲟人工模拟产卵场的构建、中华鲟潜在产卵场的筛选和葛洲坝下中华鲟产卵场的恢复与重建提供科学依据。
1 材料与方法
1.1 研究区域
研究区域位于紧邻宜昌葛洲坝下游约4 km的江段范围内, 涵盖中华鲟产卵场。位于湖北省宜昌市的葛洲坝属于低水头径流式水电站, 库区长度180 km, 坝前水位27 m, 总库容 1.58×109m3[24]。在葛洲坝建成后, 中华鲟野生种群近三十年在此产卵场进行产卵活动[14]。葛洲坝下中华鲟产卵场河床底质主要由大型卵石、中型卵石、小型卵石、沙石和黏土组成, 另外还存在部分人工护底河床, 在历年调查中未观察到泥类型的底质[19,25]。产卵场依照从上至下的顺序划分为Ⅰ—Ⅴ五个区域, Ⅰ和Ⅱ区域从左至右依次划分为A—D四个区域, 其余区域划分为A—C三个区域(图1)。

图1 葛洲坝下中华鲟产卵场及分区Fig. 1 Spawning grounds and zoning of Chinese sturgeon under Gezhouba Dam
1.2 产卵场水声学调查
河床底质水声学数据采集水声学数据主要来源于长江水产研究所在2004—2020年开展的中华鲟自然繁殖生态调查期间进行的水声学探测活动。探测使用的水声学仪器为美国BioSonics公司DT-X系列数字科学回声系统, 系统参数设置参照张辉等[22]使用的方法。采用科学回声仪配套软件Visual Acquisition 5.0.3实时接收和存储所获取的水声学数据。探测时采用“z”字型路线以8 km/h的速度进行。探测范围从葛洲坝下至下游大约4 km。
水下视频影像数据采集长江水产研究所在2016年1月期间对长江湖北宜昌中华鲟自然保护区范围内河床底部的200个采样点进行样品验证数据采集, 采样范围从葛洲坝下至松滋口大约80 km。采样目的是研究河床底质的表观特性与其水声学特征值的相关性。因此在随机采样点进行声学数据采集的同时, 对声学系统探测区域的底部进行视频数据采集。在随机采样点进行水声学数据采集与水声学调查使用的仪器参数设置是一致的, 在采集之前用锚等固定用物体将船体固定在采样点, 使用Dell Latitude D810笔记本电脑通过Visual Acquisition 5.0.3软件接收和记录声学数据。视频数据采集使用Canon G16光学相机与防水密封护套和灯(RJW7100, 中国)固定在自行设计的不锈钢笼子上,使用蔺丹清[23]的方法进行水下视频观察, 在存储卡容量范围内连续拍摄分辨率为1980×720 dpi的影像资料[23]。使用手持GPS定位系统记录采样位点的经纬度信息, 及时更新采样点位置。
1.3 数据处理
河床底质特征参数提取使用声学后处理系统Visual Bottom Typer对水声学数据进行分析,设置时变增益40 logR用于补偿回波的距离相关性。平均进行3次ping, 以最小化底部的ping到ping变化。硬度、粗糙度和分形维数都是使用该软件对相应声学回波进行归一化处理的参数。一般研究认为硬度和表观充塞度存在相关关系[21], 由第一个底部回波的第一部分(从底部回波开始到回波峰值)评估; 粗糙度和河床底质的粒径存在相关关系,由第一底部回波的第二部分(从回波峰值到末端)评估[26]; 分形维数是量化声学回波信号包络形状的特征参数[28], 被认为与底质的基本元素含量相关, 由整个第一底部回波来评估[27]。
河床底质表观特征提取在2016年采集的200个采样点中有123个采样点部分底质粒径明显大于水下视频采集视野中的范围, 因此只选用77个采样点的水声学特征及表观特征数据。通过视觉检查水下视频, 使用美国渔业协会推荐的方法确定河床底质的充塞度和粒径[22]。充塞度是单位视野面积中河床质(粒径较大的砾石或卵石)的缝隙被河床推移质(粗砂和细砂)填塞的面积百分比[21], 通过视觉检查和相应软件辅助确定底质充塞度大小。使用Bain等对主要底质类型描述方法和底质观测结果[28], 将77个视频观测点的底质主要组成划分为中型卵石、小型卵石、沙石、黏土及人工护底河床共5种类型。
统计分析和空间分布分析采样点的水声学特征与河床底质充塞度、粒径的相关性通过Origin 2020分析。河床底质的声学特征参数数据连同位置信息导入ArcGIS 10.6, 使用反距离权重法构建网格大小为10 m的栅格图像。将各栅格图像转换为点数据, 导入Origin中绘制相应箱线图, 获取除去异常点后的极值和平均值。结合粒径-粗糙度拟合方程将粗糙度转换为粒径, 构建粒径空间分布图。通过均值变化找到河床质演变的关键时间节点, 统计计算关键年份产卵场分区的特征参数统计值并进行单因素方差分析, 使用ArcGIS中的重分类工具按对应图例进行分类并计算所占比例。
相关性结果用于底质类型的判别分析, 水声学特征参数栅格图像结果用于底质类型的空间分布分析, 重分类结果和箱线图结果用于分析葛洲坝下中华鲟产卵场的底质演变。
2 结果
2.1 底质声学特征与表观特征的相关性
通过水下视频视觉检查发现, 在所有采样点中,河床质主要组成大都为卵石和沙的非均质混合物,纯沙和纯卵石仅在部分区域可见。在所有样本中,充塞度为0—100%, 底质粒径为0—200 mm。声学分析发现, 硬度基本在0—0.05, 粗糙度为0—0.007,分形维数为0—0.20。进一步的回归分析发现, 充塞度(E)和硬度(H)之间的关系可以用Boltzmann模型进行拟合, 方程为E=3+95/{1+exp[(H-0.00679)/0.00232]}(R2=0.74; 图2a), 中 等 粒 径(D)和 粗 糙 度(R)之间的关系可以用logistic模型进行拟合, 方程表示 为D=285-285/(1+R/0.000824) (R2=0.87; 图2b)。另外分形维数(FD)的数值大小可以很好地区分河床底质的主要组成成分(图3)。

图2 后向散射参数和底质表观特征之间的关系Fig. 2 Relationship between backscattering parameters and substrate phenology

图3 分形维数与河床质主要组成的关系Fig. 3 Relationship between fractal dimension and major composition of riverbed mass
2.2 底质水声学特征变化
葛洲坝下中华鲟产卵场河床质水声学特征均值在2004—2020年发生了明显的变化, 图4—6显示该区域的3种水声学特征参数变化趋势是一致的,3种特征参数在2004—2008上升, 在2008—2012年下降, 在2012—2019年持续上升, 在2019—2020年下降, 但是变化幅度有所区别。硬度均值在2004—2012年的变化不明显, 在0.0046—0.0054波动, 其在2012—2015年增加78.37%, 在2015—2019年增加115.56%, 在2019—2020年减少52.18%。粗糙度均值波动较为明显, 自2005年增长到0.001后保持稳定, 从2008年开始减少, 在2008—2012年减少22.21%,在2015—2019年增加75.13%, 在2019—2020年减少68.88%。分形维数均值从2004—2005年增长116.91%后, 在2005—2012年保持在0.108—0.123内波动,2015—2019年维持在0.127—0.138波动, 2020年下降到0.12。可以发现2008年、2012年、2015年、2019年和2020年是底质特征变化的关键时间节点。

图4 葛洲坝下中华鲟产卵场硬度年际变化Fig. 4 Interannual variation in hardness of Chinese sturgeon spawning grounds under Gezhouba Dam
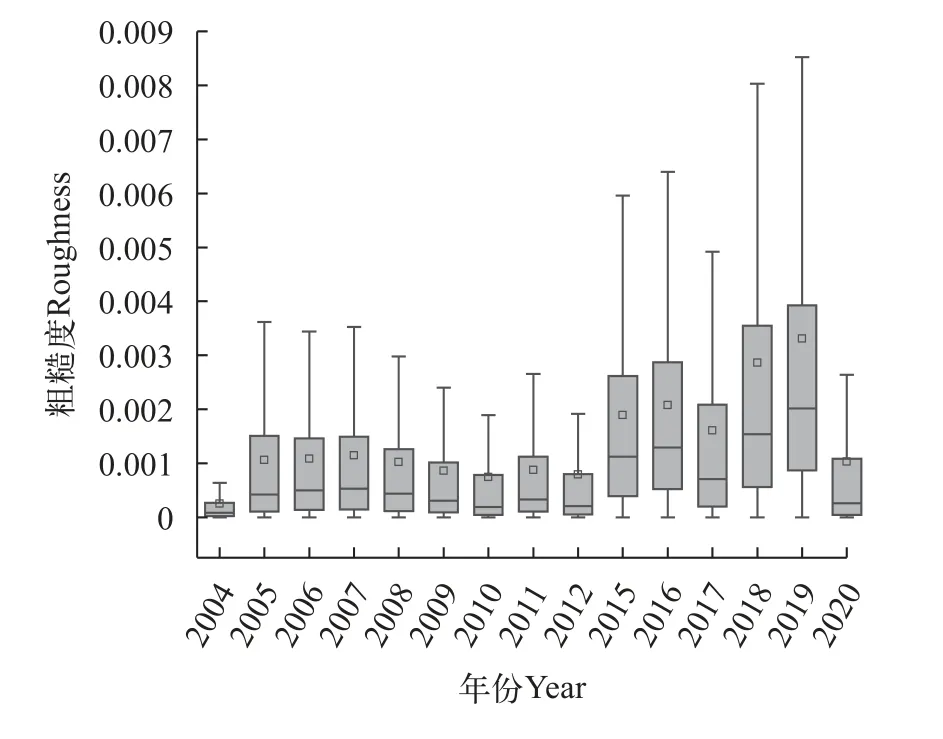
图5 葛洲坝下中华鲟产卵场粗糙度年际变化Fig. 5 Interannual variation in roughness of Chinese sturgeon spawning grounds under Gezhouba Dam

图6 葛洲坝下中华鲟产卵场分形维数年际变化Fig. 6 Interannual variation of fractal dimension of Chinese sturgeon spawning grounds under Gezhouba Dam
2.3 河床质特征演变过程
稳定产卵区底质变化的相应关键时间节点的特征空间分布图如下(图7—10)。结果显示在ⅡA区域三种特征参数一般较小, 且常年维持在一定的水平。ⅠB、ⅡB、ⅡC、ⅢB、ⅤA和ⅤB区域的底质较为稳定, 声学参数特征一般较大, 其他区域每年都有较大的变动。

图7 葛洲坝下中华鲟产卵场硬度空间分布Fig. 7 Spatial distribution of hardness of Chinese sturgeon spawning grounds under Gezhouba Dam
关键年份的三种声学参数的显著性差异结果显示, 葛洲坝下中华鲟产卵场底质在2004年的硬度与2012年无显著差异(P≥0.05), 在2008年的粗糙度与2020年无显著差异(P≥0.05), 在2008年的分形维数与2020年无显著差异(P≥0.05), 除此之外硬度、分形维数各年均有显著性差异(P<0.01; 表1)。

表1 关键年份葛洲坝下中华鲟产卵场底质声学参数统计值Tab. 1 Statistical values of substrate acoustic parameters of Chinese sturgeon spawning grounds under Gezhouba Dam in key years
栅格图像重分类的统计结果显示在2004年、2008年和2012年硬度小于0.007的部分占据整个研究区域的85%以上, 到了2015年其所占比例锐减至52.22%, 2019年就只有23.59%, 2020年回升至57.42%(表2)。粗糙度和分形维数也都有相似的变化, 在2004年、2008年和2012年, 粗糙度和分形维数较小的区域占据的比例均较大, 2012—2019年期间占据比例减少较多, 2020年才开始增加(表3和表4)。

表2 中华鲟产卵场底质硬度占比Tab. 2 Percentage of substrate hardness in Chinese sturgeon spawning grounds (%)

表3 中华鲟产卵场底质粗糙度占比Tab. 3 Substrate roughness ratio of Chinese sturgeon spawning grounds (%)

表4 中华鲟产卵场底质分形维数占比Tab. 4 Substrate fractal dimension ratio of Chinese sturgeon spawning grounds (%)
表5显示2004年整个产卵场基本上是以卵石和沙的混合底质为主, 2008年卵石就占据成为产卵场的主要底质, 黏土几乎消失, 2012年沙和卵石的混合底质类型增加, 2015年河床质基本以卵石为主,2019年含沙的底质比例进一步减少, 直至2020年,含沙底质的比例开始回升。

表5 中华鲟产卵场底质占比Tab. 5 Percentage of substrate in spawning grounds of Chinese sturgeon (%)
葛洲坝下中华鲟产卵场的底质变化主要是推移质和卵石的运动, 而卵石底质在一般情况下推移距离较短, 因此主要是推移质量的变化。在2004年至2020年整个时间段, 底质组成经历从含大量沙、含适量沙到含极少量沙, 最后沙量回升的过程。
3 讨论
3.1 河床质特征的水声学参数研究
早在1984年, 研究学者就确认了河床质可以被声学特征所区分[6], Daniel等[11]结合一定的光学观测建立了声学特征和底质类型之间的关系, 并以此对巴德河的底部进行分类。2011年杜浩等[20]发现中华鲟产卵场的硬度和粗糙度的大小明显与底质类型相关, 但是其后使用Sonar 5 Pro获取的声学特征判别底质类型存在一定的困难, 原因可能是缺乏归一化处理声学回波的步骤[22,23], 研究获取的声学特征与底质特征相关关系区分底质效果较差。
本研究在之前研究的基础上, 使用Biosonics声学后处理系统对数据进行归一化处理, 结合水下视频影像提取的底质表观特征, 分析三种声学参数与底质表观特征的相关性, 并以此对中华鲟产卵场底质进行分类, 绘制该研究区域的底质分类图, 描述中华鲟产卵场的底质演变过程。
由于水下视频设备的限制, 粒径超过200 mm的底质没有得到展示, 图8e显示在分布图中有部分粗糙度的数值是大于200 mm底质对应的粗糙度,实际葛洲坝下中华鲟产卵场粒径大于200 mm的河床质确实占据了一定的比例, 因此粒径-粗糙度拟合曲线对后续的推测趋势应该是正确的, 但具体的验证还需要设备条件的更新。2004—2012年葛洲坝下中华鲟产卵场整体硬度变化不大, 但是实际其底质含沙量明显下降[21], 这能够由充塞度和硬度的相关关系解释。图2a的散点分布显示, 前期充塞度和硬度的相关性较差, 例如在硬度为0.005时, 充塞度可以在20%—80%波动, 因此推断在推移质和卵石底质混合且推移质较多的情况下, 硬度极有可能不仅与推移质的填充量相关, 还与推移质在卵石之间填充的方式、河床质的孔隙度等特征相关。若结合图3对底质进行分类, 那么大部分采样点的底质主要组成就应为人工护底, 但是实际上葛洲坝下江段的人工护底仅占很小的一部分。因此应对底质的分形维数数值范围进行修正, 图9显示2012年后葛洲坝下中华鲟产卵场的分形维数以0.07—0.20为主, 而此时其河床质的主要组成为中型卵石和小型卵石, 主要组成为中型卵石的分形维数范围应该修正为0.07—0.20, 以此修正范围可以更加准确地对底质进行分析。

图8 葛洲坝下中华鲟产卵场粗糙度空间分布Fig. 8 Spatial distribution of roughness of Chinese sturgeon spawning grounds under Gezhouba Dam

图9 葛洲坝下中华鲟产卵场分形维数空间分布Fig. 9 Spatial distribution of fractal dimensions of Chinese sturgeon spawning grounds under Gezhouba Dam

图10 葛洲坝下中华鲟产卵场粒径空间分布Fig. 10 Spatial distribution of grain size of Chinese sturgeon spawning grounds under Gezhouba Dam
3.2 长江水文事件与产卵场河床质变化的相关性
2003年6月, 三峡大坝首次进行蓄水运行, 而后其水位不断抬高, 至2008年9月初步开展175 m蓄水计划[29], 期间蓄水运行导致长江上游转移至中下游的沙等推移质在坝区内淤积, 葛洲坝江段推移质的量不断被冲刷而补充减少, 原本的平衡被打破。2004 年12月开展了葛洲坝下近坝江段的河势调整工程[30], 期间产卵场区域的含沙量受到了一定的影响。因此在2004—2008年, 葛洲坝下中华鲟产卵场河床底质发生了明显的改变, 推移质减少, 充塞度显著降低、卵石暴露。2008年长江中上游干流及多条支流均发生了历史同期最大洪水, 且当时汶川地震造成了大量的河道崩岸, 上游推移质转移较多。在长江宜昌站监测获取的2008—2013年的输沙量, 除2011年较少外, 其他年份均保持在一定的水平(图11), 此期间洪水带来较多的沙可能改变了三峡大坝蓄水造成的清水下泄现象, 期间推移质得以补充, 因此2008—2012年该区域充塞度增加。
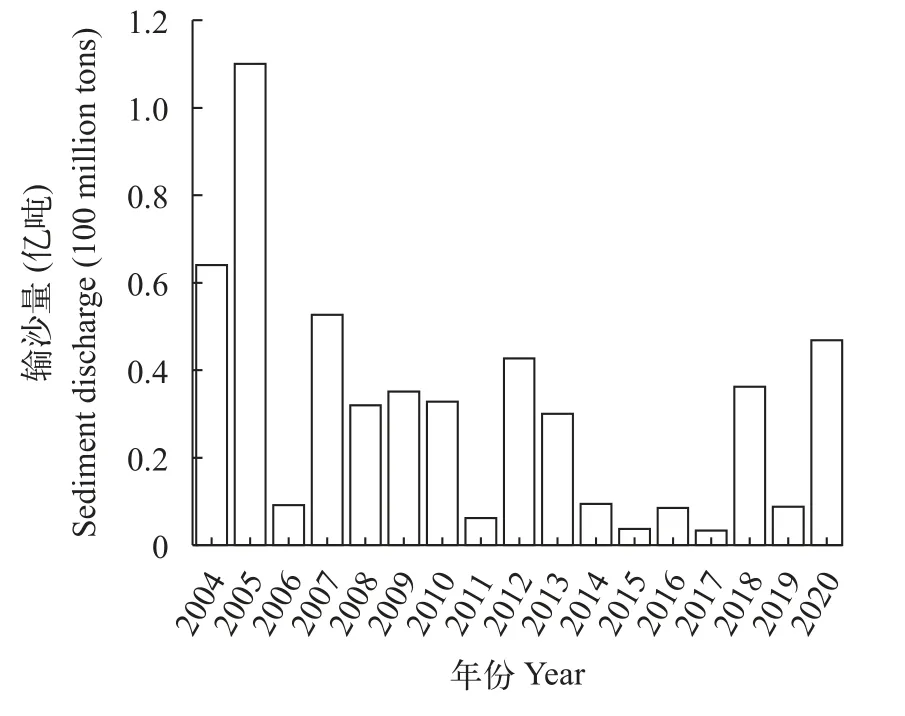
图11 长江宜昌站输沙量年际变化Fig. 11 Interannual variation of sand transport at Yichang Station of Yangtze River
从2012—2015年, 粗糙度、硬度和分形维数的均值增加趋势明显, 但由于2013年和2014年进行数据采集是使用其他水声学仪器, 无法提取和其他年份相同处理的水声学特征参数, 因此无法判断水声学参数在2012—2015年是逐步上升还是在某一年突然增加的, 依据宜昌站监测的输沙量变化可能能够解释这一变化: 在2012年长江上游的向家坝和各种新建水坝的相继投入运行后, 2013年开始宜昌站输沙量减少, 2015年到达最小值(图11), 葛洲坝下中华鲟产卵场的推移质被进一步冲刷而得不到补充。由此推测, 2012—2015年该区域含沙量应该是逐渐减少的, 粗糙度和硬度逐年上升。葛洲坝下江段的底质泥沙含量还取决于三峡大坝的运行状态,比如2016—2020年多发的洪水导致三峡大坝库区内泥沙堆积过多, 三峡大坝在2020年8月进行了沙峰排沙调度[31], 葛洲坝下中华鲟产卵场底质含沙量因此在2019—2020年增加, 而 2018年7月虽然也进行了沙峰排沙调度, 但可能由于排沙时间较早和排沙量相对较少, 2018年中华鲟繁殖生态调查期间产卵场泥沙留存量较少, 底质状况并未改善。
通过葛洲坝下中华鲟产卵场底质的演变分析发现, 该江段河床质变动的主要原因有水利工程的建设运行、长江流域的洪水发生以及人类活动。大型水利工程对葛洲坝下中华鲟产卵场河床质的影响是逐年增加的, 尤其是在其上游多个水利工程相继运行后, 中华鲟自然繁殖所需的推移质被冲刷而得不到补充; 挖沙活动和泥沙调度能直接影响到中华鲟产卵场的底质含沙量; 长江流域的大型洪水能够改善河床质状况。若是在排除人类活动影响的前提下, 葛洲坝下中华鲟产卵场底质的推移质含量会持续减少, 中华鲟自然繁殖活动很难恢复, 但是长江流域洪水的发生可能会缓解这种不利状况,其缓解程度取决于洪水发生的位点、规模及三峡大坝的运行状况。
3.3 河床质变化对中华鲟自然繁殖的影响及中华鲟产卵场保护性建议
长江水坝导致的一些制约因素对中华鲟的自然繁殖行为是有显著累积影响的[14], 底质因素就是众多制约因素之一。在三峡蓄水运行初期, 其蓄水程度还没有达到较高水平, 2004—2008年产卵场各个区域的含沙量减少程度有所不同, 但是中华鲟所在的Ⅳ和Ⅴ部分区域的充塞度还维持在一定水平,因此除了在2003年后中华鲟自然繁殖次数由2次转为1次, 中华鲟的自然繁殖活动依然能监测到。2008—2012年中华鲟产卵场的上产卵区(Ⅰ、Ⅱ区), 其河床质的变化较下产卵区(Ⅲ—Ⅴ区)明显,更适合中华鲟自然繁殖, 这可能是中华鲟主要产卵区域迁移的原因之一。2012—2015年中华鲟适宜繁殖底质逐年减少, 这可能是中华鲟自然繁殖首次中断和再次中断的诱因。2015—2019年粗糙度和硬度持续增加使得葛洲坝下的中华鲟产卵场已不适宜繁殖, 2017年中华鲟自然繁殖连续中断至今。中华鲟的自然繁殖活动明显与底质状况相关。
在长江流域三峡大坝和上游向家坝等多个水坝蓄水运行的情况下, 中华鲟产卵场的恢复与重建是困难的, 但是仍然存在部分希望, 建议: (1)通过历史数据筛选中华鲟适宜的产卵条件(地形、底质、水温等), 在长江湖北宜昌中华鲟自然保护区的范围内, 筛选葛洲坝产卵场下游其余区域条件适宜的江段, 进行部分改造使其成为新的中华鲟产卵场;(2)在保证中华鲟繁殖需求的流速和水温的前提下,调整三峡大坝进行泥沙调度活动的时间和规模, 将坝区排沙的开始时间延后至9—11月期间或者在2020年三峡大坝进行泥沙调度规模的基础上增加坝区排沙量, 使葛洲坝下江段在中华鲟繁殖期间得以维持良好的底质状况, 为中华鲟恢复产卵活动提供适宜环境, 实现葛洲坝下中华鲟产卵场的功能修复。
4 结论
本研究利用丰富的水声学数据, 结合水声学的方法和水下视频技术对葛洲坝下中华鲟产卵场进行了深入的研究, 完善了底质特征之间的相关关系,找到了底质变化的关键时间节点和原因, 并推断了中华鲟自然繁殖中断可能的原因, 研究结果对中华鲟的种群存续问题来说意义重大, 但也存在着部分不足, 比如在底质水声学特征和表观特征的相关性研究中, 粒径大于200 mm的底质未有研究, 相关关系对较大的粒径判别可能有偏差, 需要后续的设备和技术更新; 在葛洲坝下中华鲟产卵场底质演变过程的分析中, 只是解析了推动底质演变的原因, 而没有对其进行量化, 对研究区域后续的底质变化只能大致推测; 本研究只是对葛洲坝下江段的底质变化进行了研究, 但并没有获取中华鲟的适宜底质特征, 下一步工作可以对以往产卵区域的底质水声学特征进行精确提取并判别。这些不足之处都需要科研工作者们去一步步解决, 为下一步筛选中华鲟潜在产卵场、建立人工模拟中华鲟产卵场乃至葛洲坝下中华鲟产卵场的修复与重建等提供科学依据和方案。
本研究的主要结论: (1)粗糙度、硬度、分形维数三种声学特征分别与河床底质的粒径、充塞度和主要组成类型相关, 粗糙度和硬度与相应表观特征能建立相关公式, 在河床质充塞度较大的情况下(20%—80%), 依据硬度-充塞度拟合曲线判别底质效果较差。(2)2008、2012、2015、2019和2020年是葛洲坝下中华鲟产卵场河床质变化的重要时间节点, 研究区域内的主要变化如下: 含沙底质比例在2004—2008年减少33.08%, 在2008—2012年增加12.69%, 在2012—2015年 减 少33.83%, 在2015—2019年减少6.29%, 2020年含沙量回升, 相比2019年增加39.37%。在2004—2020年整个时间段, 底质组成经历一个从含大量沙、含适量沙到含极少量沙,最后沙量回升的过程。(3)结合长江流域发生的水沙变化事件, 发现水利工程的建设运行、长江流域洪水的发生及人类活动都会影响河床质的演变进程: 水利工程蓄水运行造成的清水下泄会导致其下游的推移质持续减少, 长江流域洪水的发生有利于泥沙从上游向中下游转移, 河势调整工程和泥沙调度等人类活动则会直接影响泥沙含量的变化。(4)河床质的演变进程会影响中华鲟的自然繁殖状况: 中华鲟自然繁殖活动在2013年首次中断, 在2015年再次中断, 2017年连续中断至今, 也正是在此期间, 葛洲坝下中华鲟产卵场底质被严重冲刷,推移质急剧减少, 中华鲟的自然繁殖活动明显与河床质状况相关。
猜你喜欢
——奋战在一线的葛洲坝水泥人
工友(2023年5期)2023-05-18
水生生物学报(2023年3期)2023-03-10
——以泉吉河为例
生态学报(2022年22期)2022-12-16
长江科学院院报(2021年8期)2021-04-03
水电与新能源(2020年8期)2020-09-01
农家之友(2020年9期)2020-04-21
中国三峡(2017年4期)2017-06-06
中国三峡(2017年4期)2017-06-06
中国三峡(2017年4期)2017-06-06
中国三峡(2016年5期)2017-01-15
