
沉香属植物内生真菌多样性及功能研究进展
2024-03-02 08:09:08黄颖,孟慧,何欣,杨云*1
中草药 2024年4期
黄 颖 ,孟 慧 ,何 欣 ,杨 云 *1
1. 中国医学科学院北京协和医学院 药用植物研究所,中草药物质基础与资源利用教育部重点实验室/濒危药材繁育国家工程实验室,北京 100193
2. 中国医学科学院北京协和医学院 药用植物研究所海南分所,海南省南药资源保护与开发重点实验室/国家中医药管理局沉香可持续利用重点研究室,海南 海口 570311
沉香是瑞香科(Thymelaeaceae)沉香属AquilariaLam. 植物含树脂的木材[1],广泛用于香料、医药、文玩收藏领域。沉香具行气止痛、温中止呕、纳气平喘功效[2],现代药理学研究表明沉香具有镇痛、镇静、抗菌、抗炎、改善心肌缺血、抗肿瘤等广泛药理作用[3]。沉香属现有的21 个种主要分布于东南亚地区,其中白木香Aquilaria sinensis(Lour.) Gilg.和云南沉香A.yunnanensisS. C.Huang 是中国本土沉香属植物,主要分布于广东、广西、海南、云南等地[4]。健康的沉香属植物不产生沉香,只有受到雷击、断枝等伤害时才分泌次生代谢物,经多年累积形成沉香。因沉香属植物资源濒危、沉香形成时间漫长,导致沉香产量低,严重不满足市场需求。常用的人工结香方法主要分为砍伤、凿洞、打钉等物理伤害法,注入植物激素、盐类等化学诱导法和人工接菌法[5]。物理伤害法结香时间长、产量低,化学诱导法结香快、但存在化学试剂污染及毒性的问题,人工接菌法一般以内生真菌为诱导剂,结香时间较短,内生真菌相比化学试剂更安全环保,且所诱导的沉香质量与天然沉香相似[6]。
内生真菌是指其生活史中某一阶段或整个阶段生活在健康植物组织或器官内,并对宿主植物没有引起明显病害症状的一类微生物群,具有促进宿主植物生长发育、提高抗逆能力、促进次生代谢物累积等功能[7]。近年来,许多研究者通过组织分离法从沉香属植物中分离出多种内生真菌,并对其多样性和功能进行研究。内生真菌在诱导沉香属植物产生沉香、提高抗胁迫能力等方面具有重要的应用价值和前景。本文重点对沉香属植物内生真菌的多样性及其诱导结香、提高生态适应性等功能进行综述,为沉香属植物内生菌的深入挖掘和利用提供参考。
1 沉香属植物内生真菌的多样性
沉香属植物的内生真菌集中分布在植物的茎部,具有丰富的种类多样性[8-9],已报道从沉香属植物中分离鉴定的内生真菌有56 属(目),见表1,其中镰刀菌属Fusariumsp.占分离总菌株的19%、毛色二孢属Lasiodiplodiasp.占17.26%、刺盘孢属Colletotrichumsp.占4.68%、木霉属Trichodermasp.占3.24%。

表1 沉香属植物内生真菌分类Table 1 Classification of endophytic fungi in Aquilaria genus
1.1 沉香属植物内生真菌的丰度与宿主植物的部位有关
结香部位分离的真菌种类及数量比白木多[10],茎部分离得到的内生真菌种类及数量多于叶部和根部[11],并且同一茎部不同高度的结香部分也存在内生真菌的丰度差异。Chen 等[1]将白木香结香部位按照从上至下分为5 层,分离内生真菌发现:最靠近结香部位的第1 层内生真菌多样性最高,接近白木的第5 层内生真菌多样性最低。环境中90%以上的微生物无法培养[12],仅通过组织分离法不能完全反映内生菌的群落结构和多样性。Liu 等[13]基于高通量测序技术分析白木香不同结香层之间的内生真菌群落多样性,共检测到247 种内生真菌,分属毛色二孢属Lasiodiplodiasp.、木霉属Trichodermasp.、褐顶孢属Phaeoacremoniumsp.等,通过对比丰富度指数和多样性指数发现腐烂层真菌多样性最高。Liu等[14]对离地0.2~2 m 的白木香茎部由下至上取不同高度结香部位样品,经高通量测序发现不同高度样品的真菌群落丰度有明显差异,且沉香品质也不同。
1.2 沉香属植物内生真菌的种类多样性因宿主所处地域或品种不同而有差异
Cui 等[15]采用组织分离法对云南和海南的白木香内生真菌进行分离,共得到28 株内生真菌,分别归属14 个属,其中毕赤酵母属仅在海南白木香中发现,枝孢属等9 个属仅见于云南白木香中。Mohamed等[16]、Chhipa 等[17]、Ramli 等[18]分别对种植在马来西亚和印度的马来沉香A.malaccensis内生真菌进行分离,得到不同种类的内生真菌。其中,位于马来西亚的马来沉香优势菌为镰刀菌属,但位于印度的马来沉香优势菌为木霉属。宋杰等[19]对茂名和东莞的5 种白木香进行高通量测序,结果表明不同区域环境中,同种白木香结香部位的真菌群落具有显著差异,且同一区域不同品种白木香结香部位的真菌群落也具有显著差异,如易结香品种中‘西瓜叶’的优势属为镰刀菌属Fusariumsp.,‘虎斑棋’的优势属为Hypomontagnellasp.。
1.3 沉香结香前后的内生真菌群落差异明显
马华明等[20]研究发现,创伤初期白木香内生真菌种类增多,始结香和已结香的真菌种类相对减少。邹欣涛等[21]基于变性梯度凝胶电泳(denaturing gradient gel elec-trophoresis,PCR-DGGE)技术分析白木香结香前后内生真菌的变化,发现结香后内生真菌在丰富度及多样性上均比未结香白木香更高。Mohamed 等[22]通过实时荧光定量PCR 证明了马来沉香也有类似的差异变化。推测创伤时大量病菌侵染加速结香,结香后期产生的沉香具有抑菌作用,真菌种类逐渐减少。说明沉香结香过程与内生真菌密切相关,且可能是多种真菌共同作用的结果。
2 内生真菌的功能
2.1 内生真菌促进沉香属植物次生代谢物积累
大量研究表明真菌可有效诱导沉香结香,并且所产生的化合物类型与用于诱导的真菌菌株种类密切相关[23]。如在白木香中接种硬孔菌Rigidoporus vinctus[1]、Phaeocremoniumrubrigenum[13]、Paraconiothyriumvariabile[24]、 可可色二孢菌Lasiodiplodiatheobromae、尖镰刀菌Fusarium oxysporum[25]等内生真菌均能有效诱导沉香形成,其中R.vinctus可在2 个月结出浸出物含量为38.9%的沉香,远超《中国药典》2020 年版药用要求,且倍半萜相对含量高达22.76%[1]。Faizal 等[26]成功通过腐皮镰刀菌Fusariumsolani诱导得到高品质沉香。此外,内生真菌还能通过离体组织共培养诱导次生代谢物产生,如Phaeocremoniumrubrigenum侵染白木香愈伤组织后能明显促进倍半萜成分的积累[13];Lasiodiplodiatheobromae代谢物处理白木香愈伤组织24 h 后检测到α-愈创木烯(5.93%)、δ-愈创木烯(1.89%)和α-蛇麻烯(26.20%)3 种倍半萜成分[27];黄绿墨耳真菌提取物可诱导白木香离体侧根组织产生4 种色酮化合物[28]。
沉香属植物接种内生真菌后能产生含有倍半萜、色酮、芳香族化合物、烷烃和脂肪酸等成分的沉香,与天然沉香主要化学成分一致。倍半萜、色酮、芳香族化合物是沉香特征成分,与沉香品质的高低有关。
2.1.1 内生真菌诱导沉香倍半萜类成分 倍半萜类化合物是沉香特征成分和主要活性成分,总含量可以反映沉香的品质。根据其基本结构可分为桉烷型、艾里莫芬烷型、螺旋烷型、呋喃型、愈创木烷型、杜松烷型、前香草烷型、蛇麻烷型和其他骨架类型[29-30],具有调节中枢神经系统、抗炎、抗肿瘤、抗菌、抗氧化等活性。Zhang 等[25]在白木香上接种尖孢镰刀菌,所结沉香的精油中含76.77%的成分为倍半萜,主要为α-桉叶醇(8.37%)、马兜铃酮(8.14%)、cis-Z-α-甜没药烯环氧化物(7.68%)、2,2,6-三甲基-1-[(1E)-3-甲基-1,3-丁二烯基]-5-亚甲基-7-氧杂双环[4.1.0]庚烷(5.21%)等倍半萜化合物。郑科等[31]在腐皮镰孢菌和可可毛色二孢菌诱导结香实验中检测到沉香螺旋醇的生成。Chhipa 等[17]在马来沉香中接种Penicilliumpolonicum发现3 个月内可诱导沉香螺醇生成。大多数真菌诱导结香的沉香倍半萜相对含量为10%~15%[32],而镰刀菌、硬孔菌诱导的沉香中含环氧异香橙烯、氧化香橙烯- (1)、沉香螺醇、愈创木醇、吉马酮、白千层醇等倍半萜化合物,总倍半萜相对含量可高达55.70%[1,33]。
2.1.2 内生真菌诱导沉香色酮类成分 色酮类化合物是沉香主要活性成分,具有抗菌、抗炎、抑制α-葡萄糖苷酶、抑制乙酰胆碱酯酶等活性[34]。Cui 等[35]将炭角菌属Xylariasp.接种到白木香中8 个月后检测到4-苯基-2-丁酮、4-(4-甲氧基苯基)-2-丁酮等色酮类化合物。Chen 等[36]在可可色二孢菌、镰刀菌诱导的沉香中检测到6,7-二甲氧基-2-(2-苯乙基)色酮。
2.1.3 内生真菌诱导沉香芳香族化合物产生 苄基丙酮等低分子量芳香族化合物是沉香主要成分之一,对小鼠具有镇静作用[37]。研究发现,可可毛色二孢菌诱导形成的沉香中可检测到苄基丙酮[36];镰刀菌诱导形成的沉香中含苯乙醛、1,2-苯二甲酸等成分[25]。白木香内生真菌YNAS04、YNAS06 和YNAS08 能大量诱导多种萘、酚等芳香性化合物的合成[38]。
2.1.4 内生真菌产生与沉香特征成分相同或类似的代谢产物 Monggoot 等[39]发现Arthriniumsp.菌丝的乙酸乙酯提取物中含β-二氢沉香呋喃、α-沉香呋喃,间座壳属和刺盘孢属中除上述2 种化合物外,还检测到δ-榄香烯、β-榄香烯、cis-二氢沉香呋喃,3 种菌株的提取物中均检测到高含量的氧化沉香螺醇,且均具有抗氧化活性。葡萄座腔菌A13 接种于白木香木屑中进行发酵,发酵产物中分离得到7 个2-(2-苯乙基)色酮类似物[40]。炭垫菌属中的挥发性成分中检测到α-芹子烯等5 种倍半萜化合物[31],毛壳菌属Chaetomiumsp.的发酵产物中分离到倍半萜烯等化合物,这些化合物具有抗菌活性[41]。由此可见,内生真菌能产生与沉香特征成分相同或相似的代谢产物,可促进沉香积累形成,并具有不受植物资源的限制的优势,有望作为沉香特征成分的替代来源。
2.2 内生真菌提高沉香属植物抵御生物胁迫的能力
在野外环境下,内生真菌可触发沉香属植物的防御机制,使植物产生具有抗菌作用的次生代谢物,保护植物免受进一步伤害。沉香树脂形成后,内生真菌的种类和数量开始减少[20]。沉香中分离的2-(2-苯乙基)色酮类化合物具有抗菌活性[42]。此外,沉香精油具有抗金黄色葡萄球菌、白色念珠菌、热带假丝酵母菌、枯草芽孢杆菌活性[43],精油中分离的异石竹烯也具有抗菌活性[44]。由此可见,沉香属植物可在内生真菌作用下,产生次生代谢物以提高对病菌的抗胁迫能力。
内生真菌代谢产物也具有抗菌活性,进一步帮助植物抵御外源性病原体入侵、提高应对生物胁迫的能力,保持植物的健康生长。沉香属植物内生真菌可产生4-羟基苯乙酸等具有抗菌活性的代谢产物[41]。内生真菌C.gloeosporioides、C.perangustum、Diaporthesp.、F.tricinctum、P.macrospinosa、Aspergillussp. A14 的液体培养粗提物也具有抗菌活性[45-46]。内生真菌可间接提高沉香属植物应对生物胁迫能力,作用途径见图1。

图1 内生真菌提高沉香属植物应对生物胁迫能力的途径Fig. 1 Way of endophytic fungi improving ability of Aquilaria to cope with biotic stress
2.3 内生真菌提高沉香精油质量
内生真菌在精油提取的发酵过程中具有重要作用,可提高精油质量。一方面,植物细胞壁的主要为纤维素、果胶和木质素等成分,需要不同功能的酶共同作用才能分解[47]。微生物分泌纤维素酶、半纤维素酶和果胶酶等酶类,促进植物细胞壁中多糖的糖苷键水解,导致细胞破裂,促使胞内物质释放[48];另一方面,微生物可作为生物转化剂,通过微生物细胞的生物转化作用将植物体内的前体物质转化为挥发性化合物,改善精油质量,提高精油提取率[49]。
发酵是沉香精油提取的重要步骤,发酵后沉香精油的提取率提高,且精油中挥发性成分种类增多。Sen 等[50]的研究表明沉香经发酵后精油的挥发性成分发生改变,沉香经镰刀菌发酵45 d 后增加了γ-桉叶醇、沉香螺醇、马兜铃烯等91 种挥发性成分[51]。Islam 等[52]将3 种级别的沉香发酵21 d,发现发酵过程中所有样品的真菌数量不断增加,A 级沉香精油含有更多挥发性成分、质量最佳,发酵液中的微生物数量和种类也最多,推测微生物在发酵过程中有助于提升沉香挥发性成分的含量。沉香精油整个生产过程涉及的相互关联的生化过程还没有被深入研究,进一步了解参与沉香发酵的微生物的作用,有助于优化发酵过程,从而提高沉香精油的质量。
3 内生真菌作用机制和接菌方法
3.1 内生真菌诱导沉香形成的机制
在自然界中植物与内生真菌形成互利共生关系,内生真菌可提高宿主植物抗逆能力、促进次生代谢物合成[53]。倍半萜类化合物和2-(2-苯乙基)色酮类化合物是沉香主要成分和特征成分[5]。内生真菌通过调节植物体内的激素水平[54],甲羟戊酸(mevalonic acid,MVA)途径[55]促进植物自身合成倍半萜类物质。
在白木香中,内生真菌促进树体生成游离脂肪酸,引发脂肪酸氧化级联反应,导致水杨酸、茉莉酸产生,水杨酸和茉莉酸又可诱导白木香倍半萜合酶基因(sesquiterpene synthase gene,ses TPS)的表达促进倍半萜类化合物合成[56-57],从而累积形成沉香。张鹏等[58]发现白木香受到真菌感染时,抗氧化酶活性增强,茉莉酸、水杨酸、乙烯前体含量在感染初期显著升高,促进萜类和色酮类物质的合成;Sen 等[51]在镰孢菌-马来沉香的相互作用中发现植物受到真菌刺激后,脂氧合酶(lipoxygenase,LOX)通路激活生成游离脂肪酸,促进茉莉酮酸酯等植物激素的产生。茉莉酸作为诱导倍半萜形成的关键信号,诱导倍半萜合酶基因ASS1的表达促进倍半萜合成[59]。内生真菌还可通过诱导转录因子-甲羟戊酸(transcription factors-mevalonic acid,TFs-MVA)通路磷酸化而促进沉香倍半萜的累积。植物中的倍半萜来源于异戊烯基二磷酸( isopentenyl pyrophosphate,IPP),IPP 可通过植物细胞中的MVA途径合成[60]。IPP 通过缩合反应形成倍半萜的前体物质-法尼基二磷酸(farnesyl diphosphate,FPP),FPP 经倍半萜环化酶形成倍半萜类化合物[61]。Liu等[12]研究发现在受到P.rubrigenum感染的白木香愈伤组织中检测到MVA 相关酶显著上调,下游法尼基焦磷酸合酶(farnesyl diphosphate synthase,FPS)基因、Ses TPS1 和Ses TPS2 显著上调,表明真菌诱导的倍半萜生物合成主要通过MVA 途径增加。在倍半萜生物合成过程中,52.9%TFs 的蛋白磷酸化水平发生了显著变化,磷酸化MYB(V-myb avian myeloblastosis viral oncogene homolog)转录因子、碱性亮氨酸拉链(basic leucine zipper,bZIP)转录因子和WRKY 转录因子与倍半萜生物合成途径酶高度相关。另外,在P.rubrigenum诱导3 或18 d 后,转录因子ARR2、ALR2和NAC17表达上调,推测P.rubrigenum促进倍半萜生物合成可能存在另一种调控机制。
此外,色酮类化合物是沉香主要活性成分之一,真菌侵染白木香过程中通过调节查耳酮合酶(chalconesynthase,CHS)基因表达促进色酮的积累。研究发现,白木香在腐皮镰刀菌的刺激下,2 个月后CHS基因表达上调,随后检测到色酮累积,说明沉香中色酮的积累与白木香受到真菌刺激后CHS基因表达上调相关[62]。内生真菌诱导色酮类化合物形成的过程尚未进行深入研究,进一步研究诱导机制和代谢途径可为内生真菌的应用提供参考。
3.2 沉香属植物的接菌方法
自20 世纪90 年代起,内生真菌就作为接种剂用于人工诱导结香[63]。并非所有的内生真菌都可用于诱导结香,表2 列出了具有诱导能力的沉香属植物内生真菌。
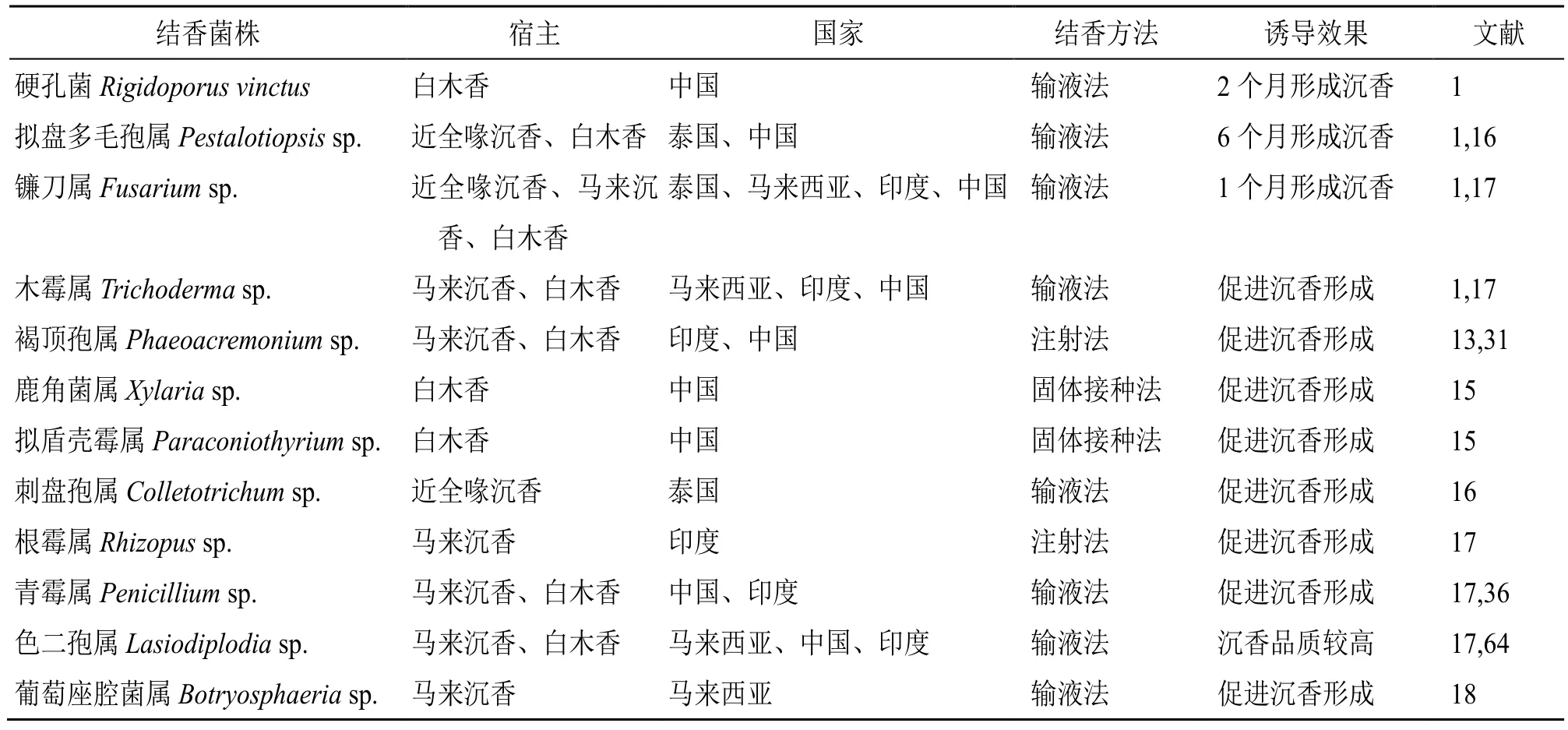
表2 具有诱导结香能力的沉香属植物内生真菌Table 2 Endophytic fungi of Aquilaria with ability to induce agarwood formation
沉香属植物的接种方法主要有固体接种法和液体接种法2 种。固体接种法将含菌丝的固体培养基填入树孔中,随后孔洞附近产生褐色树脂[24,35]。此法操作简单,但产量较低。液体接种法有输液法、注射法以及喷施法等,其中以输液法为主。输液法利用植物自身的蒸腾作用把输液装置中的菌液输送到植物内部[36],此法的沉香产量较高,但存在菌丝体堵塞输液管的问题。注射法即通过注射器将少量菌液注入孔中,等待菌株侵染后结香,但由于操作较繁琐,结香产量低等推广较少。喷洒菌液法是在白木香树干上锯出缺口并喷洒菌液,以塑料薄膜包扎,一段时间后缺口附近产生褐色树脂[65]。该方法操作简单,但结香效率偏低,且树木锯口后易受台风雨水的影响。
4 存在问题与展望
沉香属植物内生真菌具有丰富的多样性,其功能及代谢产物蕴含巨大的应用价值。内生真菌不仅能促进沉香属植物产生沉香,其种类还影响次生代谢物的类型,导致实际生产中沉香品质发生变化。因此,研究沉香属植物内生真菌及其功能对实际生产具有重要意义。
目前,内生真菌分离鉴定的方法较为成熟,已报道的沉香属植物内生真菌种类较多,但存在以下问题:(1)沉香内生菌的分离鉴定方法多采用传统培养方法,对于难以培养的内生真菌,分离鉴定仍是难题;(2)沉香属植物内生菌大量研究集中在诱导结香的效果上,对机制的研究较少,内生真菌和沉香属植物的互作机制、代谢途径和关键酶的发掘尚且不足;(3)目前尚未发现可广泛用于沉香实际生产的内生真菌,实际应用中缺乏高效、便捷的接菌技术;(4)内生真菌具有促进植物生长、增强抗逆能力等能力,大部分研究只关注沉香属植物内生真菌诱导结香,缺乏与促生、抗逆及发酵产物等其他功能研究;(5)对沉香属植物内生真菌代谢产物的种类和活性研究较浅,其生物合成途径和作用机制尚不明确。
针对这些问题,今后对于沉香属植物内生真菌的研究可包括以下几个方面:(1)借助高通量测序技术对沉香属植物内生真菌进行深入挖掘,或以培养组学方法进行内生真菌培养,辅以基质辅助激光解析电离飞行质谱MALDI-TOF-MS、内转录间隔区(internal transcribed spacer,ITS)测序技术等技术建立新型的内生真菌分离鉴定方法,充分挖掘沉香内生菌资源,为筛选高效结香菌株提供材料;(2)结合蛋白组学、转录组学及代谢组学探究内生真菌-沉香应激响应与代谢共调,挖掘内生真菌诱导倍半萜或色酮生物合成的关键通路与转录因子,为真菌诱导沉香提供参考;(3)挖掘优质促结香菌株、改进现有接种技术,利用内生真菌定向增加沉香中某类特征成分的含量,在较短时间内促进高品质沉香生产;(4)探讨沉香属植物和内生真菌共生关系,挖掘内生真菌促进沉香属植物的生长和生物防治功能;(5)筛选内生真菌次生代谢物中具有药用价值的活性物质,结合发酵技术大规模生产,进一步探索其生物合成途径。
综上,随着沉香产业发展,内生真菌相关研究也在不断深入。开发绿色、安全、高效、环保的内生真菌沉香结香技术,拓展其在促生、抗逆、发酵、生物合成等方面的应用,可以为沉香属植物内生真菌及其代谢物研究和沉香产业发展提供参考。
利益冲突所有作者均声明不存在利益冲突
猜你喜欢
中草药(2023年14期)2023-07-21 07:02:02
CHINA TODAY(2022年8期)2022-08-03 07:41:34
小小说月刊(2022年14期)2022-07-18 07:37:02
趣味(作文与阅读)(2021年3期)2021-07-19 06:10:34
世界科学技术-中医药现代化(2021年12期)2021-04-19 12:31:28
中成药(2018年1期)2018-02-02 07:20:11
中成药(2017年10期)2017-11-16 00:50:02
中国现代中药(2017年8期)2017-09-21 08:32:42
现代装饰(2017年11期)2017-05-25 02:15:19
金色少年(奇趣科普)(2017年1期)2017-03-03 07:05:33
