
太行山猕猴的交配行为参数与模式
2024-02-26 10:36:16何民强田军东李文韬路纪琪
兽类学报 2024年1期
何民强 田军东 李文韬 路纪琪*
(1 郑州大学生命科学学院,郑州 450001)(2 郑州大学生物多样性与生态学研究所,郑州 450001)
生存与繁衍是动物的本能,尽管多样的无性繁殖方式产生“新个体”效率明显较高,但有性繁殖被认为必然具有重要的进化意义和巨大的现实意义 (Crow, 1994; Lively and Morra, 2014; Cohen and Marron, 2023)。在营体内受精的有性繁殖动物,交配是雄性使雌性成功受孕的前提条件 (Brennan and Orbach, 2020)。但是,在自然条件下,由于难以直接观察、易受观察者出现的影响、交配持续时间通常较短以及难以与某些社会行为相区分等原因,对非模式脊椎动物的交配行为往往知之甚少 (Bernsteinet al., 1980; Brennan and Orbach,2020)。为此,开展野生动物交配行为研究,不仅是认识动物繁殖生物学的重要环节,也是探究自然选择和性选择如何作用于交配行为的基础,亦有助于理解动物有性繁殖演化这一进化生物学问题 (Lively and Morra, 2014; Cohen and Marron,2023)。
动物的交配行为是性选择的基础,涉及雌雄双方,而自然选择和性选择则均可作用于交配行为 (Brennan and Orbach, 2020; Dixson, 2021; Faselet al., 2023)。根据交配行为的发生进程,可将动物的交配划分为交配前、交配过程和交配后,其中交配过程涉及雄性将生殖细胞输送至雌性体内的过程,交配前则主要涉及配偶选择以使双方常相伴随但尚无交配现象,而交配后则指雌雄双方完成交配且常彼此在空间上呈分离状态 (Opieet al., 2012;尚玉昌,2014; Firmanet al., 2017; Richards, 2017; Dixson, 2021; Petersen, 2022; Cezar and da Silva Evangelista Junior, 2023)。有关动物交配前和交配后的性内和性间配偶选择,已有较多的研究报道 (尚玉昌,2014; Brennan and Orbach, 2020;Petersen, 2022; Sentenskáet al., 2023)。但是,有关动物交配过程的量化分析却较为匮乏,以致尚未充分理解交配模式在有性繁殖动物中如何演化的问题 (Dewsbury and Pierce Jr, 1989;贾志云等,1999; Dixson, 2021)。
交配模式存在种间变异性和种内相对稳定性,影响交配模式的因素有雄性生殖器结构、婚配制度、社会组织、有效性比和性状的波动非对称性等 (Dixson, 2012)。在哺乳动物的交配过程中,依据有无锁结 (genital lock)、 有无抽动 (pelvic thrust)、是否多次插入 (multiple intromission)、是否多次射精 (multiple ejaculation) 或是否长时插入(prolonged intromission),可将雄性哺乳动物的交配模式划分出16 个类型 (Dewsbury, 1972; Dixson,2021)。其中,非人灵长类动物的交配模式可归于第3、10、11 和12 类型 (Dewsbury, 1972;贾志云等,1999; Dixson, 2021)。猕猴属 (Macaca) 已命名物种25 种 (Strier, 2021; Ghoshet al., 2022),为分布最广泛的非人灵长类类群,具有多雌多雄式的社会结构特征,雌性多留居于出生群而雄性常于性成熟期前后迁出出生群,交配模式有单次爬跨射精 (single-mount-to-ejaculation, SME) 和多次爬跨射精 (multiple-mount-to-ejaculation, MME) 2 种类型 (Thierry, 2011; Dixson, 2021)。例如隶属fascicularis种组的日本猴 (M.fuscata) 被归为无锁结、有抽动、多次爬跨射精型 (熊成培和王岐山,1991),隶属sinica-arctoides种组的红面猴 (M.arctoides) 则表现为有锁结、有抽动、单次长时插入跨爬射精型 (Dixson, 2012)。
猕猴 (M.mulatta) 是自然界中分布最为广泛的非人灵长类物种,其典型社群由多个成年雄性和多个成年雌性以及未成年个体组成,雌性终身留居于出生群,雄性则多在性成熟前后离开出生群而扩散至其他群体,从而常形成以母系关系为纽带的社群特征,并且群内成年个体间具有严格的线性等级关系,成年雄性间等级关系更加严 格(Thierry, 2011;田军东等,2011a)。猕猴与日本猴、食蟹猴 (M.fascicularis) 和台湾猴 (M.cyclopis) 被划为fascicularis种组,该种组的交配模式被归于第10 类型,即无锁结、有抽动、多次插入和无长时插入 (Dixson, 2012)。然而,以往对猕猴交配模式的研究多在圈养或半圈养条件下实施,甚至未观察到交配中出现抽动、多次插入等现象(Dewsbury and Pierce, 1989)。猕猴华北亚种 (M.m.tcheliensis) 现仅自然分布于太行山南端,故而被称为“太行山猕猴”,是中国现生最北端的野生猕猴 (路纪琪,2022)。迄今为止,对太行山猕猴的繁殖生态已有较多认识,但对其交配模式的研究仅限于圈养群体 (侯进怀等,1998) 以及野外群体的初步观察 (李德森等,1990),关于野外条件下交配过程的量化分析尚属空白,遑论与近缘物种的比较研究。为此,本研究基于个体识别,对太行山猕猴国家级自然保护区的济源五龙口地区太行山猕猴的交配前、交配过程和交配后的行为进行了观察和量化分析,以期了解太行山猕猴的交配行为参数和交配模式,深化对猕猴属物种繁殖生态学的认识。
1 研究方法
1.1 研究地点
本研究地点为河南太行山猕猴国家级自然保护区的济源五龙口地区 (北纬35°12′49″,东经120°41′25″)。该保护区位于河南与山西交界的太行山南端,为中国地形第二阶梯向第三阶梯过渡区,略呈东西走向、中间略南凸的狭长片区,区内山势陡峭,海拔600 ~ 1 200 m;气候属大陆性季风气候,四季分明,冬冷夏热,年均气温14.3 ℃,年均降水量675 mm,雨水偏于夏季;植被类型以暖温带落叶阔叶林为主,兼具部分针叶混交林,森林覆盖率达92%以上 (宋朝枢和瞿文元,1996;胡玉梅等,2004)。
经过30 余年的保护,太行山南端太行山猕猴数量由20 世纪90 年代的约2 000 只增至约4 000 只(路纪琪,2022)。济源五龙口地区由20 世纪80 年代初的1 群50 余只发展至多群数百只,目前该区域核心区的太行山猕猴群体主要包括WKL-1A、WKL-1B、WKL-2 和WKL-3 群,除每天有指定人员定时、定点进行补投食物 (玉米、花生等) 之外,猕猴仍可取食周边可利用自然食物资源,如野葡萄、壳斗科植物种子、荆条嫩茎、君迁子果实等植物以及部分动物 (以节肢动物幼虫及成虫为主)(宋朝枢和瞿文元,1996; Luet al., 2007)。
1.2 研究对象
本研究以WKL-1A 群为目标群,该群由原WLK-1 群 (180 余只) 于2017 年分群形成,继承了WLK-1 群的主要成年雄性和雌性个体,并且分群时该群体数量 (约120 只) 较WLK-1B 群 (约50 只)大。基于体型、毛色、面部、伤痕等特征,对群内猕猴进行个体识别并命名。参考刘振河等(1986) 和田军东等 (2011a),对群内个体进行性别-年龄组划分。2018 年11 月至2019 年10 月,该群包括92 只猕猴。其中成年雌性31 只,成年雄性10 只,亚成年个体11 只,青少年个体28 只,2018年春夏季出生婴猴19 只,新生婴猴死亡7 只,成年雄性JB于2018年12月迁出本群,此外无其他迁入迁出或个体死亡现象。
交配需要雌雄双方参与,雌性猕猴具有月经周期但可受孕状态信号并不明显 (Thierry, 2011),从而难以判定雌性是否因处于可受孕状态而参与交配,而成年雄性个体在交配期则可能随时参与交配,因而以成年雄性为对象可更好地进行行为数据采集。此外,猕猴社群内成年雄性数量相对较少(成年雄雌性比约为1∶2.9) (Thierry, 2011),选择以成年雄性为对象则可在相对较短时间内,对群内所有成年雄性进行观察与行为记录,从而可更有效地采集数据。由于可能存在雌性个体与群外雄性交配的现象,以群内雄性个体为对象可能错失部分数据,但群外交配现象相对较少 (Ruiz-Lambideset al., 2017),故而本研究以WLK-1A 群内成年雄性为对象。
1.3 行为数据采集
研究期间 (2018 年11 月至2019 年10 月),于每日08: 00—17: 00 (冬季时段,11 月至翌年3 月) 或18: 00 (夏季时段,4—10 月),以WLK-1A 群内的成年雄性为对象,利用焦点动物取样法对成年雄性进行行为数据采集。所采集行为参考太行山猕猴PAE 行为谱 (田军东等,2011b),主要包括社会理毛 (social grooming)、跟随 (following)、挨坐(contact siting)、邀配 (solicitation)、嗅阴 (anus-genital sniffing)、 交配 (copulatory behavior)、 爬跨(mounting) 等行为数据。其中,爬跨是指雄性后肢抓握雌性后肢而前肢常按压在雌性背部但不伴随射精现象,交配则是在爬跨基础上发生射精现象,射精过程常伴随雄性身体短暂性僵持及退下后乳白色精液遗落出并固化。每日开始观察时随机选取1只成年雄性为焦点对象进行观察,当该个体在前2 min 观察时间内与雌性发生爬跨或交配则将其作为有效焦点对象,对其开展10 ~ 20 min 行为取样;若所观察雄性个体未在前2 min 内与雌性发生爬跨或交配行为,则放弃对该雄性个体的观察,进而随机选择其余未观察雄性。为保证数据覆盖度,在取样过程中,确保每2天对群内所有成年雄性个体进行至少1 次的10 ~ 20 min 的观察。此外,为便于后期核查数据,观察过程中使用数码摄像相机 (HDR-AS50, Sony Corporation) 进行摄制录像。采用连续记录法 (continuous recording method)记录数据,包括行为类型、发起者、接受者和持续时间。
研究期间,共采集有效焦点数据935 条,总观察时间15 270 min,共记录到爬跨或交配行为1 049 条。因成年雄性JB 于2018 年12 月底迁出研究群,且爬跨或交配数据仅有5条,故未将其加入数据分析中。
1.4 猕猴属交配行为数据检索
为探究猕猴属物种交配行为参数及交配模式与系统发育关系的相关性,检索并分析了该属物种交配行为参数及模式相关文献。分别在百度学术和中国知网、谷歌学术和Web of Science 中,以关键词“猕猴AND 交配模式”和“Macaca AND(Mating pattern OR Copulation behavior)”分别检索,以获取包含猕猴属物种交配行为参数及模式的文献。共检索到红面猴(Chevalier-Skolnikoff,1975),食蟹猴、猕猴和冠毛猕猴 (M.radiata)(Shivelyet al., 1982),日本猴和藏酋猴 (M.thibetana) (熊成培和王岐山,1991),豚尾猴 (M.nemestrina) (Tokudaet al., 1968) 7 个物种的交配行为参数与模式,包括交配时间、爬跨时间、爬跨次数、抽动次数、射精时长以及交配模式。
1.5 数据处理
所采集数据利用Microsoft Office 2016 进行整理与汇总,数据以平均值 ± 标准差 (mean ± SD) 展示。依据基于全基因组数据所分析灵长类系统进化关系文献 (Kudernaet al., 2023),以几内亚狒狒(Papio papio) 为外群,绘制了猕猴属部分物种的系统进化树 (图1);依据文献 (Thierry, 2011),将猕猴属物种分为3 个种组和4 个社会风格类型,以分析该属物种交配模式与这三者的关系。利用二项式检验对配对行为发起者进行检验,差异的显著性水平设为α = 0.05。
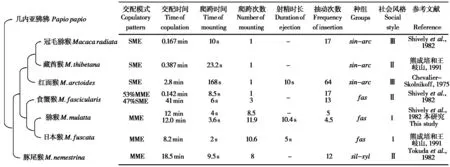
图1 猕猴属7 个物种交配模式和参数与系统发育树、种组、社会风格关系. SME:单次爬跨射精;MME:多次爬跨射精;sin-arc:sinica-arctoides种组;fas:fascicularis种组;sil-syl:silenus-sylvanus种组Fig. 1 Relationship between copulatory pattern and parameters and the phylogenetic tree, group category, social style in seven macaque species.SME: single-mount-to-ejaculation; MME: multiple-mount-to-ejaculation; sin-arc: sinica-arctoides group; fas: fascicularis group; sil-syl: silenus-sylvanus group
2 结果
2.1 交配前行为
社会理毛是太行山猕猴交配前的主要行为(49.8 %,n= 3166),雌性个体较雄性发起较多的行为为社会理毛 (32.8%和16.9%)、跟随 (14.5%和7.5%) 和挨坐 (11.2%和7.9%),而雄性较雌性发起较多的行为为邀配 (5.2% 和3.2%) 和嗅阴(0.8%和0%)。此外,交配前行为主要由雌性个体发起 (61.8%) (二项式检验,P< 0.05)。
2.2 交配过程
雌雄个体配对成功 (初次观察到成年雌性与雄性发生爬跨行为并伴有射精现象) 后,在主动或因受到外界刺激 (其他个体突然吼叫、跑跳等) 而发起交配时,雄性常立即爬跨雌性,以前肢按压雌性背部,后肢抓握雌性后肢下部或着地,雌性多呈四肢着地站立。雄性猕猴的阴茎可在爬跨前或爬跨过程中勃起,随后插入雌性阴道。在插入期间,雄性个体常快速抽动阴茎,随即从雌性个体身上退下,然后两者挨坐或进行社会理毛,直至下一次爬跨开始,此过程称为一个爬跨回合 (图2)。通常情况下,爬跨回合可重复十余次,平均 (11.9 ±1.3) 次,随后雄性常表现出身体痉挛,伴有张嘴、露齿及发生鸣叫等现象,尤其是有射精现象 (所排出精液常迅速固化并呈凝胶状)。交配后雄性个体通常会取食 (92.5%) 固化的精液,雌性亦有取食。在最后一个爬跨回合中,雌性多回视雄性并伴咧嘴、呜鸣等,偶尔会用前肢抓拽雄性。此后,雌性个体会给雄性个体理毛,二者随后分离。由12 次爬跨回合以及1次射精,构成一次完整的交配过程,而此配对关系可维持约2.1 d (标准方差为2.03 d,n= 212,范围:1 ~ 11 d)。因此,太行山猕猴的交配模式属于多次爬跨射精型。

图2 太行山猕猴一次完整交配过程示意图Fig. 2 Schematic diagram of a complete copulation of Taihangshan macaque
研究期间,共记录到1 049 次爬跨或交配行为,其中包含射精的交配有86 次,占总交配次数的8.3%。太行山猕猴一次完整的交配过程持续时间为 (719.4 ± 43.3) s (n= 42,范围:190 ~ 1312 s),平均每次爬跨时长为 (3.6 ± 0.05) s (n= 963,范围:1 ~ 13 s),平均每次爬跨间隔持续 (67.4 ±2.03) s (n= 876,范围:11 ~ 558 s),平均爬跨(11.9 ± 1.3) 次才能到达射精状态,平均每次射精持续 (10.4 ± 0.21) s (n= 86,范围:4 ~ 16 s),平均射精间隔时间为 (588.8 ± 106.6) s (n= 4,范围:333 ~ 768 s)。
2.3 交配后行为
在一次完整的交配过程完成后 (图2),参与双方一般不会立即分离,而是进行社会理毛、挨坐休息等。在所观察的80 次交配后行为中,雌性为雄性理毛占19.4%,而雄性为雌性理毛仅占3.8%;成年雌性和雄性个体休息、移动和跟随的占比分别为18.1%、 5.0%、 3.8% 和19.4%、 5.6%、0.6%;仅有1 例雄性在交配后觅食,1 例雄性攻击雌性。
2.4 猕猴属部分物种交配模式比较
基于系统演化关系、传统分类群组与社会风格,比较了猕猴属部分物种的交配模式及参数。其中,系统演化关系分析结果支持传统分类群组的划分,且各物种的交配模式类型和系统演化关系与传统分类群组均基本吻合,仅食蟹猴呈现出以多次爬跨射精 (MME) 型为主 (53%) 的交配模式。从交配参数来看,物种间变化较大, MME 型的物种爬跨次数为8 ~ 12次。社会风格分组与交配模式类型匹配关系不甚明显,仅提示社会风格偏严格 (Ⅰ和Ⅱ型) 的物种可能倾向于MME型,而偏宽松 (Ⅱ和Ⅲ型) 的物种则倾向于SME型 (图1)。
3 讨论
本研究发现,太行山猕猴的交配模式为多次爬跨射精型;一次完整的交配过程包括12 个爬跨回合和1次射精,持续时长平均为 (719.4 ± 43.3) s;交配前行为以社会理毛为主,而交配后行为则以社会理毛和休息为主;猕猴属物种的交配参数存在种间差异,交配模式与系统发育关系和传统群组分类基本吻合,但与社会风格相关性不明显。
交配是营有性繁殖动物成功繁衍的前提,亦是自然选择和性选择发挥作用的关键环节之一(Ågmo and Moralí, 2022; Cohen and Marron, 2023)。性选择理论认为,高等动物中因雌性个体繁殖成本常高于雄性,故雌性在繁殖中占主动地位,常被称为挑剔的雌性 (choosy female) (de Vries and Lehtonen, 2023)。本研究发现,雌性太行山猕猴是交配前行为的主要发起者,并且在配对期间常通过社会理毛等行为来维持配对关系,该结果与日本猴和斯里兰卡猕猴 (M.sinica) 的研究结果一致(Tokuda, 1962; Chevalier-Skolnikoff, 1975),但与李德森等 (1990) 对太行山猕猴交配行为的观察结果相反,可能是由于后者不仅样本量小 (7 例),且研究方法欠规范 (仅在拍摄纪录片时进行观察,而未进行基于个体识别的系统观察)。此外,本研究发现,配对个体间社会理毛行为所占比例最高(49.7%),且主要由雌性个体发起,这可能是雌性采取的维持配偶关系的策略,即通过投入理毛行为以获取交配机会,符合生物市场理论认为的“投入-收益”现象 (Noë and Hammerstein, 1995;Sentenskáet al., 2023)。
在自然选择、性选择的作用下,动物演化出具有物种特异的生殖系统特征与交配模式,可根据有无锁结、有无抽动、是否射精前插入以及是否多次射精4 个特征判定动物的交配模式 (Dewsbury, 1972; Dixson, 2021)。依据此标准,太行山猕猴的交配模式可归属于第10 类型,即交配过程中无锁结、有抽动、多次插入和多次射精型——多次爬跨射精,该过程与圈养太行山猕猴的研究结果相一致 (侯进怀等,1998)。
本研究量化了太行山猕猴具体交配过程并进行了属内种间比较分析。首先,本研究发现,成年雄性太行山猕猴平均经历约12 个爬跨回合才完成射精,其交配模式属于多次爬跨射精型,与同属于fascicularis种组的日本猴相似,但有别于sinica-arctoides种组的单次爬跨射精型 (熊成培和王岐山,1991)。前人研究发现,猕猴等哺乳动物存在性高潮现象,而性高潮的演化可能与确保成功受孕相关 (Lodé, 2019; da Silvaet al., 2023)。由此看来,猕猴的多次爬跨射精可能是为提升受孕而演化的交配模式,或者是雌性为了暂时“垄断”所选雄性个体而形成的配偶保护策略 (Dixson, 2021)。其次,频繁爬跨之后才射精,可能与猕猴属物种的社会组织与社会结构密切相关。在Ⅰ类社会风格中,猕猴和日本猴社群内成年个体间具有严格的线性等级结构,而且成年雌雄性比偏向雌性个体,但由于雄性需要“多次爬跨射精”而导致难以实现有效交配。这既表明了雌性在繁殖过程中占主导地位,亦可能使得高等级雄性无法垄断可繁殖雌性资源,而低等级雄性或群外雄性亦可被雌性选择,进而获取一定的繁殖机会,或者低等级雄性及群内出生雄性因高等级雄性的排斥而倾向于离群寻求繁殖机会,从而演化出雌性留群、雄性扩散的社群特征 (Shivelyet al., 1982; Caldecott, 1986; Thierry, 2011)。
致谢:感谢济源五龙口风景管理局对本研究的大力支持;感谢太行山猕猴国家级自然保护区济源管理局对本研究的许可。
猜你喜欢
保健与生活(2024年4期)2024-02-02 15:53:28
小猕猴学习画刊(2022年10期)2022-01-01 04:48:21
青年文学家(2021年29期)2021-11-19 16:11:47
童话世界(2020年32期)2020-12-25 02:59:14
小猕猴智力画刊(2019年4期)2019-05-08 21:56:28
小猕猴智力画刊(2019年3期)2019-04-19 00:02:10
祝您健康(2018年1期)2018-01-10 14:20:58
天津诗人(2017年2期)2017-11-29 01:24:14
中国火炬(2014年8期)2014-07-24 14:30:25
中国火炬(2011年7期)2011-07-25 10:38:31
