
唐古特瑞香开花物候及繁育系统研究
2024-02-01 07:54祝睿彦张春云王海平王勤礼王建东张春梅
草地学报 2024年1期
何 瑞,闫 芳,,4*,祝睿彦,张春云,王海平,王勤礼,王建东,徐 涛,张春梅,陈 叶
(1. 甘肃农业大学园艺学院,甘肃 兰州 730070; 2. 河西学院祖师麻原植物应用技术研究所,甘肃 张掖 734000;3. 甘肃泰康制药有限责任公司,甘肃 武威 733000; 4. 甘肃省河西走廊特色资源利用重点实验室,甘肃 张掖 734000)
唐古特瑞香(DaphnetanguticaMaxim)为瑞香科(Thymelaeaceae)瑞香属(DaphneLinn.)多年生常绿小型灌木,主要分布在我国的温带、热带地区,如陕西、甘肃、青海、四川、云南、西藏等海拔3 000 m左右的山坡灌丛、林缘、沟谷地带[1],因此又称甘青瑞香、甘肃瑞香、陕甘瑞香等。该物种株高0.6~1.0 m左右,分枝多且长势不规则,枝上长有绒毛,单叶互生,呈披针状,叶缘常外卷,花叶同生,雌雄同花,花期大致在5—6月,花型多以数朵呈顶生或腋生头状花序,花色艳丽,花冠外侧为紫色、浅紫色或白色,内侧则以白色常见,花香浓烈,结果期在6—7月,核果多为红色,蚕茧型[2]。该物种在临床医学中应用十分广泛,具有很高的药用价值,其干燥的根皮和茎皮为我国传统中药材祖师麻的重要基源成分之一[3],藏药中则以全株入药,被称为‘基原省相那玛’[4],根皮、茎皮具有消肿止痛、祛风除湿等功效;其叶、果可治疳积等;其花可治肺痈等[5]。近年来,随着该物种药用价值的深度挖掘,市场需求量不断增大,药农为追求利益最大化,对野生唐古特瑞香进行连根采挖,加之其自身野生繁殖量小、坐果率低、自然更新能力弱、生长繁殖缓慢等特性制约,导致野生唐古特瑞香资源逐年匮乏,极大限制了该物种的资源可持续利用。关于唐古特瑞香的研究主要集中在有效成分含量测定[5]、化学成分分析[6-7]、药理作用分析[8]、胁迫生理响应[9]、组织培养[2]等领域,但对该物种开花物候、花器官特征及繁育系统的研究鲜有报道,仅刘淑娟等[10]对唐古特瑞香的花部性状及坐果情况进行了初步研究。
繁育系统是植物内在遗传机制和外部环境影响相互作用下的生理表达,对种群的优化及动态具有重要意义[11],其亦是直接影响后代遗传组成的所有有性特征的总和,主要包括开花进程,花部形态特征,花粉活力与柱头可授性,花粉-胚珠比,杂交指数,访花者观察等方面[12-13]。本研究以唐古特瑞香野生种群植株的花器官为试验材料,对其开花物候、花部特征及繁育系统进行观察和研究,深入了解唐古特瑞香开花进程及生殖特性,探究其繁育类型及开花量大坐果率低的限制因素,以期为今后规模化、优质化人工繁育唐古特瑞香及祖师麻药材产量提升提供理论依据和实践基础。
1 材料与方法
1.1 试验地点
本试验所研究的唐古特瑞香野生种群位于甘肃省武威市天祝县安远镇东直沟村(102°52′18″ E,37°14′53″ N),该地区地处天祝藏族自治县北部,乌鞘岭北麓,属温带大陆性半干旱气候,海拔2 700 m左右,年平均气温-0.2℃,年日照时数3 300 h,年平均空气湿度53%,年降雨量600 mm,无霜期年平均120 d左右。1月平均气温-13℃,7月平均温度16℃。
1.2 开花进程及花部形态特征观测
参考Defni等[14-15]的方法,于2023年5—6月对唐古特瑞香进行群体、花序及单花等3个水平的开花进程进行观测。于现蕾期随机选定10株长势基本一致,生长情况良好的植株,记录单株的始花期(植株中5%的花朵开放期日)、盛花期(植株中50%的花朵开放期日)、末花期(植株中95%的花朵开放结束期日)以及花期持续时间;选定植株中随机标记10个花序记录花期,每日定时观察花序形态及变化情况。于盛花期随机采集30朵发育良好的单花进行解剖观察,用游标卡尺(申工牌,精度为0.02 mm)对花各部位进行测量。
1.3 花粉形态、活力及柱头可授性测定
将完全盛开单花的花药置于5 mL的离心管,用搅拌棒将花药研碎释放花粉,用磷酸缓冲液漂洗3次,再经不同浓度的乙醇(70%,80%,90%,99.5%)逐级脱水,脱水后再用乙酸异戊酯置换乙醇,放入电热恒温鼓风干燥箱(DHG-9123A)进行干燥后,利用扫描电子显微镜(Quanta 450 FEG)对花粉粒群体、个体及其表面纹饰进行观察、测量并拍照,重复3次。
参考陈宇娇等[16]的方法,采集各时期的单花,花粉活力测定采用I2-KI法和TTC染色法,重复3次。柱头可授性采用联苯胺-过氧化氢法,将各时期单花柱头连同子房浸入联苯胺-过氧化氢反应液(1%联苯胺∶3%过氧化氢∶水的体积比4∶11∶22)中,用体视显微镜(奥斯微T2-3M180)观察并拍照。根据柱头是否呈蓝色以及柱头周围是否有气泡产生判断其可授性,柱头染色越深且气泡越多表明可授性越强。重复3次。
1.4 繁育系统类型评判
1.4.1花粉胚珠比(P/O)测定 采集待开的单花(花药成熟但未散粉)20朵,将全部花药移至离心管,放入设置为28℃的电热恒温鼓风干燥箱(DHG-9123A)进行24 h烘干,干燥完成后用搅拌棒将离心管内的花药研碎释放花粉。用70%的乙醇稀释0.01 g果胶酶后,用蒸馏水定容至100 mL。取5 mL果胶酶溶液滴入装有花粉的离心管后进行24 h摇匀,使花粉全部散开。用血细胞计数板统计花粉粒总数(P),重复3次。依据Cruden等[17]的标准计算P/O值并判断繁育系统类型。闭花受精(P/O为2.70~5.40);专性自交(P/O为18.10~39.0);兼性自交(P/O为31.90~396.90);兼性异交(P/O为244.70~2 558.60);专性异交(P/O为2 108.00~19 525.00)。
1.4.2杂交指数(OCI)判定 采集盛花期的单花30朵,测量花朵直径、开花行为以及柱头与花药的空间距离。参照Dafni[14]杂交指数的判断标准:(1)花朵直径<1 mm,记为0;花朵直径在1~2 mm,记为1;花朵直径在2~6 mm,记为2;花朵直径>6 mm,记为3。(2)对花药开裂和柱头有可授性的时间先后顺序进行比较,如同时成熟或者雌蕊先熟记为0;雄蕊先熟则记为1。(3)当柱头与花药高度空间距离相同时被记为0;如若两者间空间分离记为1。以上三者相加之和为OCI值,当OCI=0时为闭花受精;当OCI=1时为专性自交;当OCI=2时为兼性自交,有异交可能;当OCI=3时为兼性异交,有时需要传粉者;当OCI=4时为异交,部分自交亲和,多数需要传粉者。
1.4.3访花者观察 参考徐杰[18]的方法,随机选择标记已经开放的30个花序连续观察3 d,并对昆虫访花行为、频率、昆虫种类等进行观察记录。每天访花观察时间为上午10:00至下午16:00,每次观察持续时间为1 h,每隔1 h观察一次并记录观察数据,并将访花昆虫进行拍照鉴定。
1.4.4开花及坐果率统计 参考刘淑娟等[10]的方法,随机选择标记未开放的30个花序,盛花期统计单花开花数,末花期后统计结果数并计算坐果率。
1.5 数据分析
采用Microsoft Excel 2021和IBM SPSS Statistics 26进行数据处理和分析比较,用Duncan′s法进行显著性差异分析,用Power point 2021,Origin 2021进行作图。
2 结果与分析
2.1 唐古特瑞香开花物候及花部形态特征
2.1.1开花进程 通过对花期物候观察,唐古特瑞香花芽在4月下旬开始萌动,5月中上旬进入始花期,5月中下旬为盛花期,6月上旬进入末花期(表1),群体开花持续时间为17~22 d,单个花序开花持续时间为8~11 d,单花开放持续时间7~9 d。

表1 唐古特瑞香开花进程Table 1 Flowering process of D.tangutica
根据对唐古特瑞香开花物候的观察,将其头状花序的开花进程分为8个时期(图1):

图1 唐古特瑞香花序开花进程Fig.1 Flowering process of inflorescence of D.tangutica注:A-B,花芽形成期;C-D,苞片开裂期;E-F,花序着色期;G-H,花序松散期;I-J,初花期;K-L,盛花期;M-N,末花期;O,凋谢期Note:A-B,Flower bud formation period;C-D,Bract dehiscence period;E-F,Inflorescence coloring period;G-H,Inflorescence loosening period;I-J,Flowering period;K-L,Blooming stage;M-N,Final flowering period;O,Withering period
(1)花芽形成期:花芽自4月上旬形成并开始膨大,花序被坚硬厚实且带有绒毛的苞片包裹,该时期持续20~25 d;
(2)苞片开裂期:花芽内单花的生长膨大使得苞片开裂,直至整个花序内的数朵单花完全露出,该时期持续3~5 d;
(3)花序着色期:花序内单花由外向内,自上而下的开始由嫩绿色变为紫色,位于花序中心位置的单花先抽出,靠近苞片位置的单花后抽出,该时期持续3~5 d;
(4)花序松散期:花序内单花抽出后不再受苞片的约束,单花之间的空间进一步释放,同时单花花冠持续膨大,由紫色变为浅紫色,该时期持续2~3 d;
(5)初花期:花序内单花由边缘向中心依次开放,花序内可见2枝新叶同步抽出,该时期持续2~4 d;
(6)盛花期:花冠完全展开,内侧为白色,花冠管口可见4粒上轮花药呈对角排列,新叶从花序内完全抽出,该时期持续5~7 d;
(7)末花期:花序内单花萎蔫,花冠开始闭合,花冠管口可见上轮花药已干枯,授粉成功的花冠管基部开始膨大,该时期持续2~3 d;
(8)凋谢期:花序整体干枯,呈黄褐色,成功坐果的单花子房持续膨大,花冠管随之脱落,该时期持续3~5 d。
2.1.2花部形态特征 唐古特瑞香为顶生或腋生头状花序,花序轴短且伴有成对新叶、新枝同生,花序直径为30~34 mm,花序高18~29 mm,每个花序的单花数量为7~15朵不等(图2)。单花均由花序边缘向中心依次开放,花冠呈十字形,辐射状中心对称,花冠直径为13.98~17.32 mm;花瓣数4,花瓣外侧呈紫红或浅紫色,内侧为白色,披针形,似丝绒质地,长5.85~7.92 mm,宽3.52~5.64 mm;花序轴基部苞片数8~15,有尖鳞片状,坚硬有韧性,呈黄绿色,外侧带有细长绒毛。

图2 唐古特瑞香花序Fig.2 Inflorescence of D.tangutica
唐古特瑞香单花为两性花,雄蕊数8,花丝极短,花药分上下2轮,上轮花药位于花冠管口,下轮花药位于花冠管中部,上、下2轮花药相距1.02~2.18 mm;雌蕊位于花冠管基部,长3.11~3.96 mm,花柱极短,介于子房和柱头之间;柱头呈绒毛状,带有可吸附花粉粒的粘液,距上轮花药3.21~5.51 mm,距下轮花药1.16~2.09 mm;子房位于花冠管基部,为卵型,呈绿色,直径为1.34~1.63 mm;子房壁厚实,珠被内胚珠数为1,居中位置(图3,表2)。

图3 唐古特瑞香单花形态及精细解剖图Fig.3 Single flower morphology and detailed anatomy of D.tangutica注:A1和A2,花侧视和俯视图;B,花离析;C,花纵剖;D,子房纵剖;E1和E2,花药Note:A1 and A2,Flower side view and top view;B,Anatomy of the flower;C,Longitudinal profile of the flower;D,Longitudinal profile of the ovary;E1 and E2,Anthers
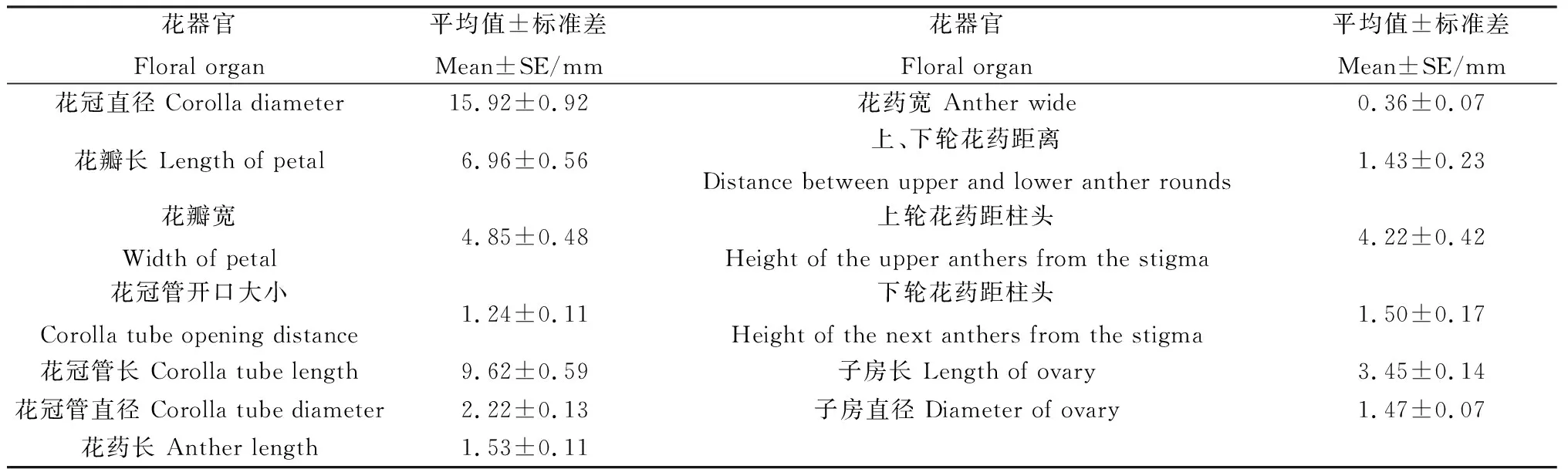
表2 唐古特瑞香花器官特征统计Table 2 Statistics of the floral organ features of D.tangutica
2.2 花粉形态、活力及柱头可授性
2.2.1花粉形态 唐古特瑞香花粉粒藏于药室内(图4A),未发育完全的花粉粒为半球形,半球横切面略微凹陷(图4B);发育完全的花粉粒呈圆球形,赤道轴长约25.27 μm(图4C)。花粉粒表面呈刺状-不规则颗粒网状复合纹饰,颗粒的不规则排列形成大小不一的孔隙,刺基部相对较大,刺较短,顶部渐尖(图4D)。

图4 唐古特瑞香花粉形态Fig.4 Pollen morphology of D.tangutica注:A,群体视野;B和C,赤道面观;D,外壁纹饰Note:A,Group vision;B and C,Polar view;D,Outer wall decoration
2.2.2柱头可授性与花粉活力动态 依据唐古特瑞香单花发育的形态特征变化,可将现蕾期至末花期划分为6个时期(图5A)。P1:单花通体为嫩绿色,花瓣与花冠管结合部有楞状凸起;P2:花瓣和花冠管开始伸长、膨大,花瓣顶尖部位开始着色;P3:花瓣和花冠管进一步伸长、膨大,花瓣顶部完全着紫色,花冠管由嫩绿色变为紫色;P4:4裂花瓣微开,花瓣外侧变为浅紫色;P5:4裂花瓣完全展开,花瓣内侧呈白色,花冠管口可见上轮4颗黄色花药,并伴有浓烈香气;P6:花瓣萎蔫卷曲,颜色由浅紫色变为黄绿色至浅褐色,香气逐渐消失。

图5 唐古特瑞香单花各发育时期(P1-P6)的花形态(A)和柱头可授性(B)变化Fig.5 Variation in flower morphology (A) and stigma fertility (B) at various developmental stages (P1-P6) of D.tangutica
柱头可授性测定结果显示(图5B),唐古特瑞香单花发育不同时期的柱头可授性存在较明显差异。P1时期柱头颜色有变化,但仅有个别气泡产生,具有微弱的可授性;P2时期柱头着淡蓝色,伴有少量的气泡产生,具有较弱的可授性;P3时期柱头着蓝色,并产生大量绵密气泡,反应强烈,可授性最强;P4时期柱头着蓝色,伴有一定量的气泡产生,具有较强的可授性;P5时期柱头着淡蓝色,伴有个别气泡产生,可授性较弱;P6时期柱头颜色有变化,仅有极个别气泡产生,表明柱头仍具有可受性,但活力极低。
花粉活力测定结果显示(图6),唐古特瑞香花粉活力随单花发育进程呈先升后降的趋势。P1-P2时期花药为绿色,花粉粒尚未成熟,不具备活力;P3时期花瓣仍处于闭合状态,此时已能透过花药壁可见花粉团粒变黄,且花粉具有一定活力,为32.49%;P4时期花药未散粉,通体呈黄色,花粉活力最强,为67.60%;P5时期少部分花药破裂,散粉量小,花粉活力有一定幅度下降,为41.59%;P6时期花粉活力显著降低,为11.66%,之后伴随花器官凋落,花粉散尽。在对唐古特瑞香花粉活力测定时,使用I2-KI染色法和TTC染色法进行对比后发现,TTC染色后的各时期花粉均未着色,无法判断花粉活力;而经I2-KI染色的各时期花粉虽活力偏高,但能够较为容易的区分花粉着色的强弱差别。
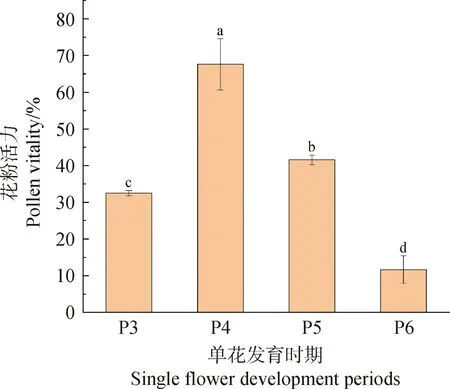
图6 唐古特瑞香花粉活力变化Fig.6 Changes of pollen vitality of D.tangutica注:不同字母表示不同时期显著性差异(P<0.05)Note:Different letters indicate significant differences between periods (P<0.05)
2.3 繁育系统类型
2.3.1花粉胚珠比(P/O) 唐古特瑞香单花雄蕊数为8,单花药平均花粉数为(97.5±18.37)粒,单花平均花粉粒数为(780±146.97)粒,单花胚珠数为1个,花粉-胚珠比(P/O)为(780±146.97),依据Cruden[17]标准判定,唐古特瑞香的繁育系统属于兼性异交(表3)。

表3 唐古特瑞香花粉胚珠比Table 3 Pollen-ovule ratios of D.tangutica
2.3.2杂交指数(OCI) 唐古特瑞香为两性花,单花花冠直径(15.92±0.92)mm,雌蕊先于雄蕊成熟,且柱头距上、下2轮花药均处于空间分离。依据Dafni[14]标准判定,唐古特瑞香杂交指数(OCI)值为4,该物种的繁育系统为异交,部分自交亲和且多数需要传粉者(表4)。

表4 唐古特瑞香杂交指数测定Table 4 Determination for out-crossing index of D.tangutica
2.3.3访花昆虫及访花行为 唐古特瑞香的访花昆虫偏好处于晴朗、无风或微风的盛花期,通常集中在一天中的10:00—16:00进行访花,访花的高峰时段为11:00—14:00,此时的访花者数量、访花者种类、访花的频次最多。下午16:00后由于气温的下降,访花者的访花行为明显减少。在阴雨或大风天气条件下,访花者的数量、种类及访花频次均存在大幅减少的情况。
有效的传粉介质是花朵进行有效传粉的必要前提之一。通过观察和鉴别,盛花期的唐古特瑞香访花者共涉及7目7科9种,包括:脉翅目草蛉科的草蛉(Chrysopaperla),鳞翅目螟蛾科的螟蛾(Pyralidae),膜翅目蜜蜂科的中华蜜蜂(Apiscerana)、三条熊蜂(B.trifasciatus)、灰熊蜂(Bombusgrahami),双翅目丽蝇科的丽蝇(Calliphoridae),半翅目盲蝽科的苜蓿盲蝽(Adelphocorislineolatus),蜘蛛目巨蟹蛛科的高脚蜘蛛(Heteropodavenatoria),缨翅目蓟马科的蓟马(Thripsvulgatissimus)(图7)。其中:中华蜜蜂、熊蜂等膜翅目的蜂类昆虫为唐古特瑞香最为有效的传粉者,其飞行速度快,持续访花的时间长,较长的吻能够有效的触及到花冠管中的2轮花药和柱头,同时翅膀的高频振动能将携带的花粉粒抖落至花冠管基部的柱头;缨翅目蓟马科的蓟马也是唐古特瑞香有效传粉者之一,其体长约为0.5~1.5 mm,能够轻易进入花冠管内基部,且其前胸后缘有缘鬃,周缘长有细长毛,能够将携带的花粉粒带至花冠管基部的柱头。其余各目、科访花者由于体型较大、无长吻等自身因素所限,且能够携带的花粉量有限,因此有效传粉效果不明显。

图7 唐古特瑞香访花昆虫Fig.7 Flower-visiting insects of D.tangutica注:A,草蛉;B,螟蛾;C,三条熊蜂;D,丽蝇;E,中华蜜蜂;F,灰熊蜂;G,苜蓿盲蝽;H,高脚蜘蛛;I,蓟马Note:A,Chrysopa perla;B,Pyralidae;C,B.trifasciatus;D,Calliphoridae;E,Apis cerana;F,Bombus grahami;G,Adelphocoris lineolatus;H,Heteropoda venatoria;I,Thrips vulgatissimus
2.3.4坐果率统计 对唐古特瑞香的开花数及坐果率统计结果显示,随机选取的30个花序的单花开花总数为346朵,坐果数为47个,坐果率仅为13.58%。其中:1个花序坐果率超过30%,7个花序坐果率在20%~30%之间,10个花序坐果率在10%~20%之间,7个花序坐果率在10%以下,5个花序坐果率为0。由此可见唐古特瑞香的开花量大,但自然坐果率极低,主要集中在20%以下。
3 讨论
3.1 唐古特瑞香的开花物候及花部形态
植物的开花物候不仅是在长期演化过程中形成的遗传特性[19],更是贯穿植物繁育系统、授粉方式、果实生产的核心基础[20]。本研究中的试验对象位于海拔2 700 m左右的野生唐古特瑞香种群分布区,该地区冷凉的气候、充足的光照、较低的平均气温等环境因素促使该物种选择相对集中的开花模式,自5月中上旬至6月初,群体花期仅持续20 d左右,而为保证在有限的花期内最大限度的完成授粉,单花序花期长达8~11 d,单花花期可持续7~9 d,这种群体花期短而集中、个体花期持续时间较长的开花行为可能是该物种为适应气候环境采取的生殖策略。
从唐古特瑞香花器官的特征来看,其10 mm左右长的花冠管和1 mm左右宽的花冠管口为典型的管状花结构,是保护雌、雄蕊在高海拔林缘、沟谷地区多变气候的有效机制;从花器官的结构来看,其花药为上、下2轮排列,每轮4颗,极短的柱头使得与子房为一体生于花冠管基部,这种柱头缩入式雌雄异位[21]的花器官结构既是该物种规避自花授粉的自主行为,亦通常被认为是应对高海拔气候条件而避免雌雄器官功能干扰的花部机制[22],但狭小的花冠管以及雌、雄蕊空间结构的分离限制了访花行为,使得体型大于花冠管或短吻的访花者不能进行有效传粉;从花器官的发育机理来看,一方面唐古特瑞香柱头成熟与花药开裂的时间存在差异,柱头成熟在先,花药开裂在后,这既是避免自花授粉的有效行为,也是延长异花授粉时间的体现。另一方面通过观察可知唐古特瑞香的柱头属于能够分泌溢出物的湿型柱头[23],可有效的黏着花粉,且柱头从现蕾期至末花期均具有可授性,这也是该物种在高海拔地区为争取最大异花传粉时间的一种自主繁殖保障措施。有花植物的花粉活力和柱头可授性的高低决定着授粉的成功率,通过柱头可授性和花粉活力测定可知,唐古特瑞香在待开期的柱头可授性最强,而在初开期的花粉活力最强,具有雌雄异熟的特性,亦符合异花传粉的机制。值得注意的是,该物种在待开期的单花4裂花瓣处于闭合状态,即使传粉者携带大量异花花粉,但也无法接触到该时期可受性最强的柱头,这可能是导致该物种花多果少、坐果率不高的主要原因之一。上述唐古特瑞香花朵性器官的不寻常特征可能也是表明了该物种生殖生物学的持续进化趋势,生殖策略的改变往往会导致果实产量的减少[24]。在花粉活力测定中,TTC染色法无法判断花粉活力,这与杨秋雄等[25]对川白芷(Angelicadahurica)花粉活力测定时采用的染色方法和结果相似。TTC染色后花粉未着色可能与花粉壁结构、花粉蛋白质含量等因素有关。
3.2 唐古特瑞香的繁育系统
有花植物在内源遗传机制和外源环境因素的共同影响下,为提高自身的适应性,持续对雌雄资源配置进行一些列调整,最终形成为多种类型的繁育系统[26],这也是大多数有花植物为保障生殖成功而进化的自交或异交的混合授粉模式[27]。根据花粉胚珠比(P/O)判定标准可知,唐古特瑞香繁育系统为兼性异交;在杂交指数(OCI)的判定标准下,该物种繁育系统为异交,部分自交亲和且多数需要传粉者。上述2项试验对该物种的繁育系统类型判定基本一致,均是以异花授粉为主,存在自交亲和的情况。在花部形态特征的观察中,唐古特瑞香艳丽的花瓣颜色,浓烈的花香,以及与花冠管口齐平的4颗上轮花药,均是吸引访花者进行异花传粉的基础[28]。而在访花者的观察过程中,也进一步证实了唐古特瑞香狭小的花冠管结构对传粉过程的影响,这种花冠管内中上部2轮花药和基部柱头的空间分布特征,更需要长吻的(如蜜蜂科-中华蜜蜂、熊蜂)或是体型微小的(蓟马科-蓟马)等有效访花者才能进行传粉。同时,中华蜜蜂、熊蜂等膜翅目昆虫作为最有效传粉者可能还与唐古特瑞香开放时花香的化学成分有着密切的关系,正如Borg-Karlson等[29]在花香对欧亚瑞香(Daphnemezereum)授粉特化作用的研究中揭示了芳樟醇(Linalool)是花香挥发物中的主要成分,其是在欧亚瑞香开花时吸引相邻栖息地雄地的春秀矿蜂(Colletescunicularius)、地花蜂(Andrenacinerea)等膜翅目昆虫进行传粉的主要原因。同科同属的唐古特瑞香可能亦是因浓烈花香气中含有某种芳樟醇吸引了周围区域的中华蜜蜂、熊蜂等进行访花。另外,从植株上发现的蜘蛛网推测,前述访花者中的蜘蛛目巨蟹蛛科的高脚蜘蛛可能不是以传粉为目的进行访花,而是作为捕食者对其他访花者进行猎食,这与范少茹等[30]对黄菖蒲访花观察的结果相似。这一情况同样间接的影响了唐古特瑞香的坐果率。
然而,植物为保障繁殖的成功,在不良传粉环境下会以自交亲和或高水平的自花授粉作为生殖策略[31-32]。在武威市安远镇气候多变的高海拔地区,唐古特瑞香进入花期后,因连续阴雨、大风、降温等天气变化,使得访花者种类、数量、访花频次的减少,阻碍了异源有效传粉,而部分自交亲和机制则能够保障其基本的繁育需求[33]。但在对访花行为的进一步观察中发现,如中华蜜蜂、熊蜂等有效访花昆虫经常会在同一花序或同一单花进行连续访花,这就不可避免将访花者携带的异花花粉粒和自花花粉粒同时落至同一柱头,使得之后的访花者能够携带的异花花粉量减少,形成花粉贴现[34],这种情况极可能导致异花花粉在花粉竞争中处于劣势,加之柱头处有着2轮花药的天然生殖屏障,进而降低了异花授粉的发生[24],丧失了萌发机会;同时在以异交为主,部分自交亲和的机制下降低了雌性适合度,亦影响自花花粉的萌发率,最终导致无法坐果。本研究在花粉活力和花粉胚珠比试验过程中发现,处于盛花期的花药开裂情况不佳,用软毛刷收集花粉粒的难度较大,需将花药放置离心管后用搅拌棒将花药研碎释放花粉,与同属的金边瑞香(Daphneodoravar.marginata)花粉在盛花期较难自然散出的情况一致[35]。此研究结果表明盛花期的唐古特瑞香散粉量不高可能也是导致该物种开花量大坐果率低的原因之一。
4 结论
综上,造成唐古特瑞香坐果率低的限制因素并非为单一性或偶然性,与其柱头可授性和花粉活力的最强时期不遇、细长的花冠管需要有效传粉者、盛花期散粉量不高、异交竞争力弱、自交亲和度低,以及有效访花昆虫的数量和种类少等因素均有密切相关性。相比于在野外大规模开展繁复且低效的人工授粉工作,寻求驯化并养殖如膜翅目的中华蜜蜂、熊蜂等有效传粉者可能是提高唐古特瑞香坐果率行之有效的措施。同时,对于该物种在盛花期散粉量不高的原因,以及可否适当降低海拔高度的人工种植来提高坐果率还需进一步研究。
猜你喜欢
中国蜂业(2023年2期)2023-04-04
今日农业(2022年14期)2022-09-15
经济林研究(2022年2期)2022-07-20
广东第二课堂·初中(2021年5期)2021-06-06
爱你(2019年17期)2019-11-14
爱你·健康读本(2019年5期)2019-06-06
数码设计(2018年12期)2018-10-20
中成药(2018年7期)2018-08-04
西夏学(2018年1期)2018-04-29
西夏研究(2017年1期)2017-07-10
