
钙镁叶面肥混合喷施减轻“妃子笑”荔枝果实“退糖”现象原因分析
2024-01-31 04:17杜婧加彭俊杰陈甜甜周开兵
中国南方果树 2024年1期
杜婧加,彭俊杰,陈甜甜,税 贤,周开兵
(海南大学,海口,570100)
荔枝LitchichinensisSonn.属无患子科、荔枝属常绿乔木,分布于我国西南部、南部和东南部,其中广东省、福建省、海南省栽培最盛。“妃子笑”荔枝是我国荔枝主栽品种,其果实风味好,但存在果皮“滞绿”现象和果肉“退糖”现象[1-3],即果肉含糖量最高时果皮未全红,而果皮全红时果肉含糖量下降,导致其内在和外观品质不能同时最佳,影响经济效益。因此,有必要开展“妃子笑”荔枝果实内在和外观品质发育不同步问题的研究。在果实生长发育期,叶面喷镁肥和果穗套袋可以克服“妃子笑”荔枝果皮“滞绿”现象。王惠聪等研究荔枝果皮花青苷合成与相关酶关系发现,只有类黄酮糖基转移酶(UFGT)活性与荔枝果皮花青苷合成关系密切,随着“妃子笑”果皮中UFGT活性增加,花青苷含量上升;套袋处理抑制UFGT活性同时也抑制花青苷合成,除袋后UFGT活性和花青苷含量都迅速增加;在“妃子笑”荔枝生理落果期结束后,叶面喷施0.3%氯化镁水溶液同样可以解决果皮“滞绿”问题,叶面喷施0.3%氯化钙水溶液提高了果肉食用品质而缓解了果肉“退糖”现象,叶面喷施0.3%氯化钙和0.3%氯化镁混合水溶液克服了果肉“退糖”现象[4-7]。根据王展等[5]研究,镁处理克服“滞绿”问题是通过提高果皮脱落酸(ABA)含量而提高UFGT活性,进而促进花色素苷积累,实现果皮提前转红和着色更红。叶面喷施钙、镁混合液克服果肉“退糖”现象的生理生化机理尚不清楚。植物体内约有10%~20%的葡萄糖通过磷酸戊糖途径(PPP)代谢,而PPP产生多种糖类物质中间产物,对果实生长发育有重要影响,推测“妃子笑”荔枝果肉“退糖”现象可能与果肉呼吸作用有关,PPP为呼吸作用的重要途径之一。本文就此研究叶面喷施钙、镁肥对荔枝果肉PPP的影响,以期探讨叶面Ca、Mg混合营养克服果肉“退糖”现象的呼吸作用机理。
1 材料与方法
1.1 材料
试验在海南省临高县金牌农场五队荔枝园开展,属于热带季风气候区,年平均气温23~24 ℃,年平均日照时数2 175 h,年平均降雨量1 100~1 800 mm,且雨热同季,土壤肥沃,砖红壤;选择生长势一致,且无任何不良表现的16年生“妃子笑”荔枝20株,4月下旬开始叶面施肥处理。当地“妃子笑”荔枝主要物候期:花期2—3月,生理落果期4月初开始,4月下旬进入果实膨大期,5月中旬进入果实成熟期。
1.2 试验设计
设置叶面喷施0.3%氯化钙水溶液、0.3%氯化镁水溶液、0.3%氯化钙+0.3%氯化镁混合水溶液,喷清水为对照等4个处理,分别记为0.3%CaCl2、0.3%MgCl2、0.3%CaCl2+0.3%MgCl2处理、对照(清水),单株小区,重复5次。
1.3 田间处理与采样方法
生理落果期结束时,即假种皮包满种子且果蒂出现红色开始不同处理叶面喷施,具体处理时间为2021年4月18日(花后35 d)、4月25日(花后42 d)和5月2日(花后49 d)上午8:00—9:00。4月18日分别在每株样树的树冠中部外围4个方向随机选取大小一致且生长中庸的果实5个作为取样参照果,分别挂牌标记,此后取样时根据参照果平均纵横径和着色状态,在树冠中部外围4个方向选取相应大小和着色状态的果实作为样果。从稳果开始至果皮全红时为止,即在花后35 d(4月18日)、42 d(4月25日)、49 d(5月2日)、56 d(5月9日)、63 d(5月16日)、70 d(5月23日)取果样,每次取果实30个,放入液氮罐速冻并带回实验室,-80 ℃冰箱保存备用。
1.4 测定指标及方法
果肉可溶性糖含量测定参考Wang等[8]的方法及蒽酮比色法,称取果肉0.5 g放进研钵,用微波炉杀青30 s,加入90%乙醇5 mL充分研磨,10 000×g离心15 min,吸取上清液再加入90%乙醇5 mL提取1次,合并两次上清液放入90 ℃水浴锅中蒸干后,去离子水定容至10 mL,用注射器吸取少量,经0.45 μm滤膜过滤后待测。使用Waters 2695高效液相色谱仪测定糖分含量,配有蒸发光散射检测器,色谱柱为Boston Green Amino Column(4.6 mm×250 mm,5 μm,120A),流动相为乙腈∶水=8∶2,流速1 mL/min,柱温35 ℃,进样量10 μL;标准品用色谱级葡萄糖、果糖和蔗糖(北京坛墨质检科技有限公司产),流动相用色谱级乙腈(国药集团化学试剂有限公司产)。
果肉总呼吸速率、过磷酸戊糖(PPP)途径呼吸速率使用便携式O2/CO2顶空分析仪(Dansensor Check Point3)测定[9],单位为mL·kg-1·h-1CO2[10];使用10 mmol/L Na3PO4作为PPP途径专一性抑制剂。先测定果肉样品总呼吸速率后,加入抑制剂测定PPP途径的剩余呼吸速率,总呼吸速率减去各途径剩余呼吸速率得出单一途径呼吸速率观测值[11]。
葡萄糖-6-磷酸脱氢酶(G-6-PDH)提取参考史青纯[12]与何亚辉[13]的方法,略作修改,称取果肉2 g,冰浴条件下加入0.05 mol/L磷酸钾缓冲液(pH值6.8)5 mL研磨,4 ℃,8 000 r/min离心20 min,取上清液待用;冷却后用酶标仪在340 nm 波长下测定吸光度(OD值),通过标准曲线计算样品酶活性。6-磷酸葡萄糖脱氢酶(6-PGDH)参考Zhang等[14]和宋丛丛[11]的方法制备酶液,用酶标仪在340 nm 波长下测定吸光度(OD值),通过标准曲线计算样品酶活性。
果肉钙、镁含量测定,称取果肉1 g左右,用去离子水研磨震荡过夜后,利用NOVAA400P原子吸收分光光度计,采用火焰原子吸收法测定水溶性钙和水溶性镁含量[15]。
1.5 数据分析
采用SAS 9.1.3软件统计分析数据,采用ANOVA作方差分析和DUNCAN法作多重比较分析。
2 结果与分析
2.1 果肉主要可溶性糖含量变化
从图1可以看出,各处理的果肉果糖均呈现上升趋势;花后56 d,对照(清水)趋于平缓;花后63~70 d,3个施肥处理均显著高于对照(清水),其余测定时期,各处理果糖均无显著性差异。花后49~56 d,0.3%MgCl2和0.3%CaCl2+0.3%MgCl2处理的葡萄糖含量显著高于0.3%CaCl2处理和对照(清水);花后70 d,3个施肥处理达到最高且显著高于对照(清水)。

图1 叶面喷施钙镁肥后“妃子笑”荔枝果肉果糖和葡萄糖含量变化
从图2可以看出,花后42 d,0.3%CaCl2+0.3%MgCl2处理的果肉蔗糖显著高于其余处理;花后56 d,0.3%CaCl2处理显著低于0.3%CaCl2+0.3%MgCl2处理;花后63 d,0.3%CaCl2、0.3%MgCl2处理显著低于其余两个处理。花后49 d前,所有处理的果肉可溶性糖均呈上升趋势;花后56 d及之后,0.3%CaCl2+0.3%MgCl2处理趋于稳定,未出现“退糖”,其余处理呈现不同程度的“退糖”现象。

图2 叶面喷施钙镁肥后“妃子笑”荔枝果肉蔗糖和可溶性糖含量变化
2.2 果肉呼吸速率变化
2.2.1 总呼吸速率
从图3可以看出,各处理的总呼吸速率动态变化呈单峰曲线,不同处理高峰时间不同,0.3%CaCl2处理达到高峰时间最早。花后35 d,各处理间差异不显著;花后42 d,0.3%MgCl2和0.3%CaCl2处理显著高于对照(清水)和0.3%CaCl2+0.3%MgCl2处理,前二者间和后二者间分别无显著性差异;花后49 d,0.3%MgCl2处理显著高于其他3个处理,后3者无显著性差异;花后56 d,0.3%CaCl2处理显著高于0.3%MgCl2和0.3%CaCl2+0.3%MgCl2处理,对照(清水)与其他3个处理差异不显著;花后63 d,总呼吸速率从高到低依次为0.3%MgCl2处理>对照(清水)>0.3%CaCl2处理>0.3%CaCl2+0.3%MgCl2处理,对照(清水)与0.3%MgCl2和0.3%CaCl2处理差异不显著;花后70 d,0.3%MgCl2处理显著高于对照(清水),0.3%CaCl2处理和对照(清水)差异不显著,0.3%CaCl2+0.3%MgCl2处理显著低于其他处理。说明0.3%MgCl2处理对总呼吸速率表现一定的促进作用,而0.3%CaCl2+0.3%MgCl2处理呈现一定的抑制作用。
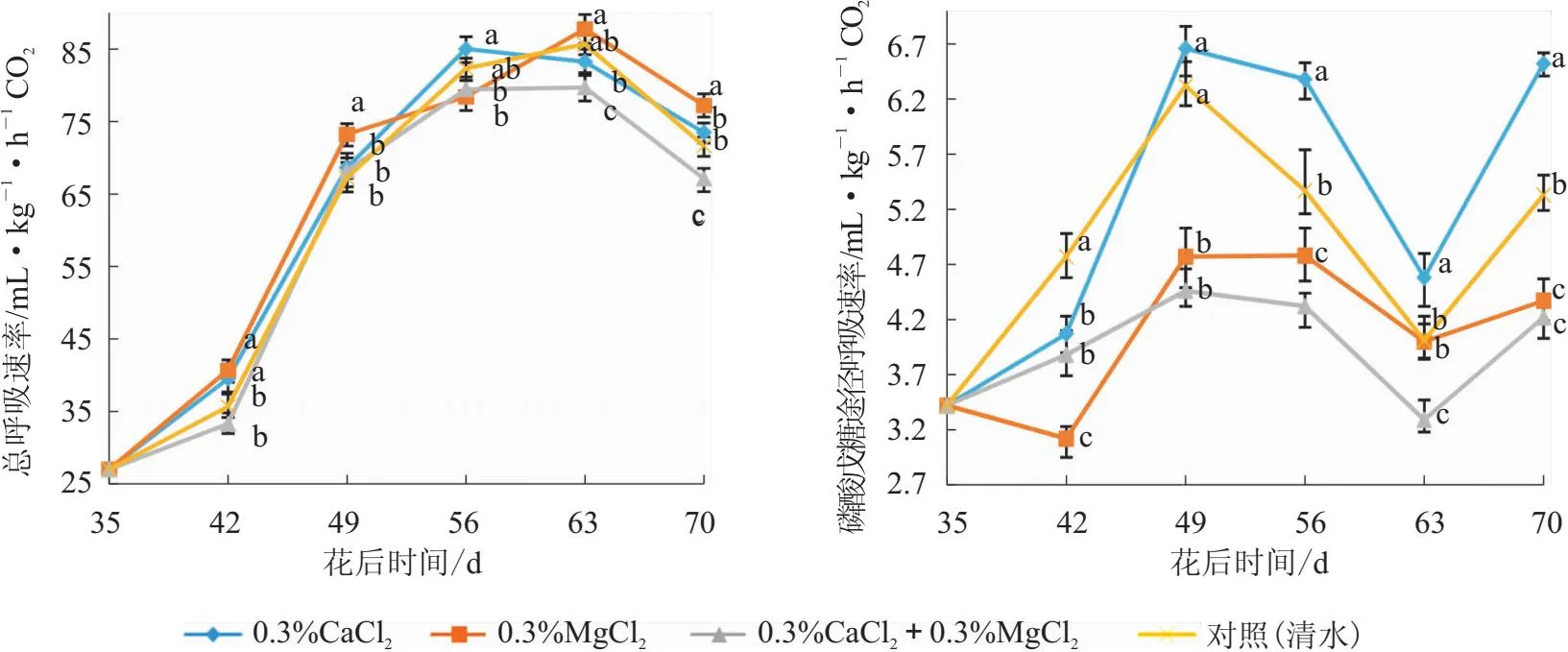
图3 叶面喷施钙镁肥后“妃子笑”荔枝果肉总呼吸速率和磷酸戊糖途径呼吸速率变化
2.2.2 磷酸戊糖途径(PPP)呼吸速率
从图3可以看出,0.3%CaCl2、0.3%CaCl2+0.3%MgCl2处理和对照(清水)PPP呼吸速率均呈现升高后降低再升高的趋势,0.3%CaCl2和0.3%CaCl2+0.3%MgCl2处理在花后56 d下降,对照(清水)则在花后49 d开始下降;0.3%MgCl2处理呈现上升至平缓的趋势。花后49 d,0.3%CaCl2处理和对照(清水)显著高于0.3%MgCl2处理和0.3%CaCl2+0.3%MgCl2处理,其两两间差异不显著;花后63 d,0.3%CaCl2处理显著高于其他处理,0.3%CaCl2+0.3%MgCl2处理显著低于其他处理,对照(清水)和0.3%MgCl2处理间差异不显著;花后70 d,0.3%CaCl2处理显著高于其他处理,0.3%MgCl2和0.3%CaCl2+0.3%MgCl2处理显著低于对照(清水),且前两者差异不显著。综上所述,0.3%CaCl2处理前期抑制PPP而后期呈现促进作用,0.3%CaCl2+0.3%MgCl2处理和0.3%MgCl2处理则抑制PPP。
2.3 PPP关键酶活性变化
2.3.1 6-磷酸葡萄糖脱氢酶(6-PGDH)活性
从图4可以看出,对照(清水)的6-PGDH活性呈现两次下降—上升—下降的变化趋势;0.3%CaCl2处理整体呈现下降—上升的趋势;0.3%MgCl2处理在前期呈下降趋势至花后42 d后上升,花后49 d又下降,至花后56 d有上升;0.3%CaCl2+0.3%MgCl2处理下降至花后49 d后趋于平缓,花后70 d略有回升。花后42 d,3个施肥处理的6-PGDH活性均显著低于对照(清水);花后56 d,各处理6-PGDH活性从高到低依次为对照(清水)>0.3%CaCl2>0.3%CaCl2+0.3%MgCl2>0.3%MgCl2,且各处理间差异显著;花后63 d和70 d,0.3%CaCl2和0.3%MgCl2处理显著高于其余处理。说明0.3%CaCl2处理前期呈抑制 6-PGDH活性的趋势,后期呈促进作用;0.3%MgCl2处理前期呈抑制 6-PGDH活性,末期呈促进作用。
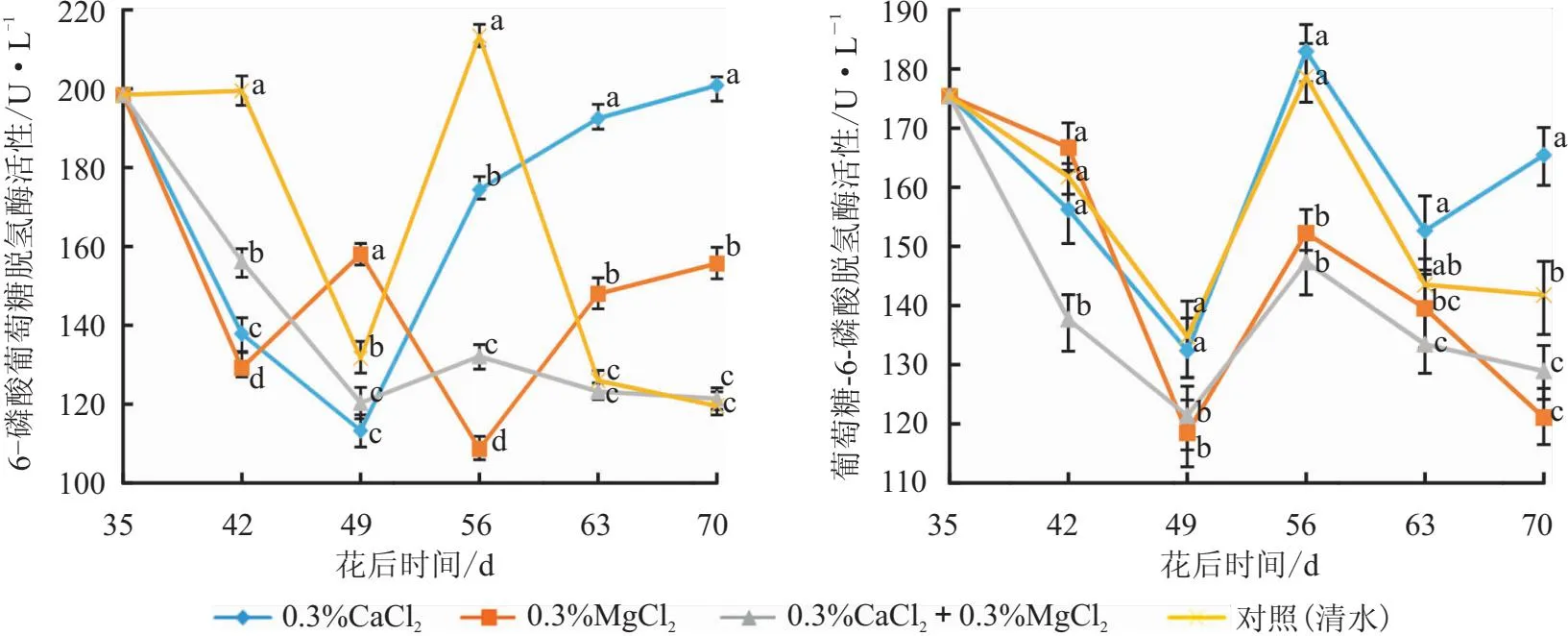
图4 叶面喷施钙镁肥后“妃子笑”荔枝果肉6-磷酸葡萄糖脱氢酶和葡萄糖-6-磷酸脱氢酶活性变化
2.3.2 葡萄糖-6-磷酸脱氢酶(G-6-PDH)活性
从图4可以看出,对照(清水)和0.3%CaCl2处理的G-6-PDH活性呈现两次下降—上升趋势,前期下降至花后49 d后上升;0.3%MgCl2处理呈现下降—上升—下降的趋势;0.3%CaCl2+0.3%MgCl2处理呈现下降—上升—平缓的趋势,于花后70 d达到峰值;花后42 d,0.3%CaCl2+0.3%MgCl2处理显著低于其他处理;花后49 d和56 d,对照(清水)与0.3%CaCl2处理间无显著性差异,且均显著高于其余两个处理,其余两个处理间均无显著性差异;花后63 d,0.3%CaCl2处理与对照(清水)无显著性差异,且显著高于0.3%CaCl2+0.3%MgCl2处理;花后70 d,0.3%CaCl2处理显著最高,而0.3%CaCl2+0.3%MgCl2和0.3%MgCl2处理之间无显著性差异且显著低于对照(清水)。说明0.3%MgCl2和0.3%CaCl2+0.3%MgCl2处理呈抑制G-6-PDH活性的趋势;0.3%CaCl2处理在采收期对G-6-PDH活性具有一定的促进作用。
2.4 果肉钙和镁含量变化
2.4.1 果肉全镁和水溶性镁含量
从图5可以看出,对照(清水)和0.3%CaCl2处理的果肉全镁含量呈现两次下降—上升趋势,均于花后56 d达到峰值;0.3%MgCl2和0.3%CaCl2+0.3%MgCl2处理在前期下降,至末期上升。花后42 d,3个施肥处理的果肉全镁含量均显著低于对照(清水),0.3%CaCl2+0.3%MgCl2处理显著最低,0.3%CaCl2和0.3%MgCl2处理之间无显著性差异;花后56 d,对照(清水)显著最高,而0.3%CaCl2+0.3%MgCl2处理显著最低;花后70 d,0.3%MgCl2处理显著最高,0.3%CaCl2+0.3%MgCl2处理次之,且显著高于对照(清水)和0.3%CaCl2处理。可见,3个施肥处理在前期降低了果肉全镁含量,0.3%MgCl2和0.3%CaCl2+0.3%MgCl2处理在后期提高果肉全镁含量。
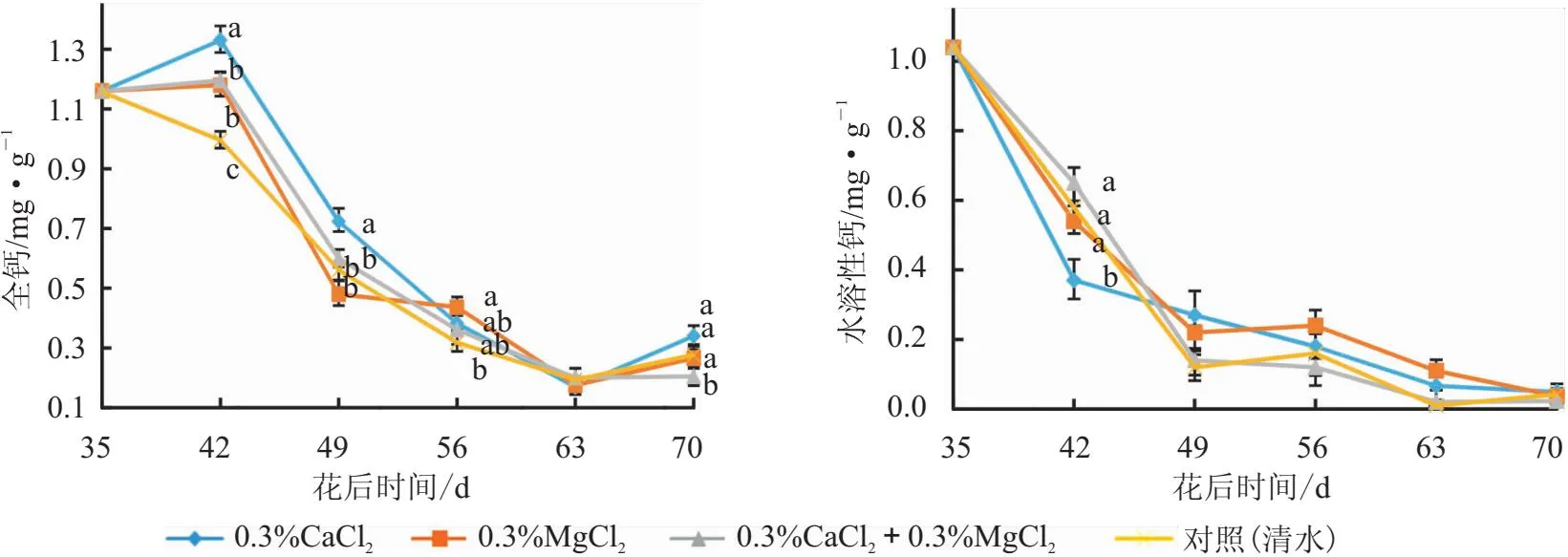
图6 叶面喷施钙镁肥后“妃子笑”荔枝果肉全钙和水溶性钙含量变化
从图5可以看出,各处理的果肉水溶性镁含量变化呈“L”形趋势。花后49 d,0.3%CaCl2处理显著高于其余处理,其余处理间无显著性差异;花后63 d,除了0.3%MgCl2处理显著高于对照(清水)外,其他处理与对照(清水)均无显著性差异;花后70 d,3个施肥处理均显著高于对照(清水)。说明不同施肥处理花后49 d后呈提高果肉水溶性镁含量的趋势。
2.4.2 果肉全钙和水溶性钙含量
从图5可以看出,花后42 d后,所有处理果肉全钙含量均呈现下降趋势;花后42 d和49 d,0.3%CaCl2处理显著最高;花后56 d,0.3%MgCl2处理显著高于对照(清水),其余处理和对照(清水)之间差异不显著。说明0.3%CaCl2处理具有提高果肉全钙含量的趋势。
所有处理果肉水溶性钙含量均呈“L”形下降趋势。花后42 d,0.3%CaCl2处理显著低于其他处理,其他3个施肥处理和对照(清水)之间无显著性差异;42d后,各处理间均无显著性差异。说明不同施肥处理基本上对果肉水溶性钙含量无明显影响。
3 讨论
3.1 对荔枝果肉呼吸途径的影响及其生理生化机制
观察荔枝果肉呼吸速率动态变化发现,采前果肉呼吸速率上升,采收期下降,这与“妃子笑”荔枝果肉采前“退糖”现象相符,说明因果肉呼吸速率升高而导致果肉糖分消耗增加,进而产生“退糖”现象。花后63 d后,0.3%CaCl2+0.3%MgCl2处理对呼吸速率具有显著抑制作用,进而降低糖分损耗,从而在一定程度上延缓果肉“退糖”。PPP分为氧化反应过程和非氧化反应过程。PPP的氧化步骤是还原型烟酰胺腺嘌呤二核苷酸磷酸(NADPH)的主要来源,它参与抗氧化剂再生[16]。该途径生成大量的NADPH+,作为供氢体参与多种代谢反应,是体内利用葡萄糖生成5-磷酸核糖的唯一途径,而核糖-5-磷酸和NADPH是核酸和脂肪酸等生物分子合成的原材料[17],也是合成DNA、RNA 的前体物质,这些物质对植物细胞分裂生长必不可缺[18]。本试验结果表明,0.3%CaCl2+0.3%MgCl2处理抑制了PPP呼吸速率,使核糖核酸合成速率降低,进而导致葡萄糖消耗减慢;由于葡萄糖和果糖在异构酶催化下可以相互转化[19],也会导致果糖消耗减少,从而使“妃子笑”荔枝果肉在一定程度上克服“退糖”现象。
0.3%MgCl2处理在采收期不同程度抑制了PPP呼吸速率,在采收期0.3%MgCl2处理的果肉可溶性糖含量呈现下降趋势,因此,0.3%MgCl2处理未能缓解“妃子笑”荔枝果肉“退糖”现象。在采收期,0.3%MgCl2处理也对PPP途径呼吸速率表现为抑制作用,但并未表现出克服“退糖”现象。植物有多条呼吸代谢途径,其中EMP-TCA循环完全有氧呼吸也是重要途径。根据吉前华等[20]的研究,镁能促进果肉EMP-TCA循环、脱羧化作用,有利于作物体内的糖类转化和磷素的吸收转化,从而0.3%MgCl2处理能促进EMP-TCA循环,导致果肉总呼吸速率未被抑制,这可能是0.3%MgCl2处理未能缓解“妃子笑”荔枝果肉“退糖”现象的重要原因。因此,只有0.3%CaCl2+0.3%MgCl2处理表现出克服“妃子笑”荔枝果肉“退糖”现象。
3.2 对果肉钙、镁含量的影响及其生理生化效应
试验结果表明,3种施肥处理对果肉水溶性钙含量影响不明显,0.3%CaCl2和0.3%MgCl2处理提高了果肉全钙含量;3种施肥处理均提高了果肉水溶性镁含量,0.3%MgCl2处理提高全镁含量。这与周媛[21]的研究报道相同,适量0.3%MgCl2处理不仅能够促进果肉三羧酸循环中有机酸含量,促进氨基酸积累,提高呼吸速率,进而明显促进果实成熟;还能够明显提高番茄根、叶片和果肉中0.3%MgCl2含量,及叶片中钾钙含量,并使果肉0.3%CaCl2含量减少。
镁元素通过促进光合同化产物运输和利用来促进果实可溶性糖积累。根据孙青慧[22]、林丽琳[23]的研究,叶片喷施0.3%MgCl2处理后,叶片内光合色素含量提高,光合作用增强,可促进糖类等碳水化合物的积累及运输,增强碳代谢。碳代谢与光合作用密切相关,叶绿素是植物光合作用的主要参与者,镁是叶绿素的主要组成成分。林丽琳[23]研究发现,镁通过影响西瓜的光合系统来影响碳代谢过程,降低碳水化合物含量;而适当增加镁素用量可提高叶片叶绿素含量,增强光合作用强度,进而促进碳水化合物和可溶性糖的积累。本研究结果表明,0.3%MgCl2和0.3%CaCl2+0.3%MgCl2处理对果肉葡萄糖、果糖含量均呈现促进作用,0.3%MgCl2处理对果肉蔗糖含量表现抑制作用。这与柑桔上的研究结果一致,即施镁能够诱导温州蜜柑果肉蔗糖转化酶活性增强,促进蔗糖转化为果糖和葡萄糖,导致果肉蔗糖含量明显降低[21]。而0.3%CaCl2处理对荔枝果肉果糖、蔗糖含量并未表现促进作用,故在采收期也出现“退糖”现象。这与甜瓜研究结果不一致,可能与果树品种或喷肥方式不同有关[24]。
本文研究结果表明,0.3%CaCl2处理表现出促进PPP呼吸速率的作用。根据田秀英等的研究,钙离子通过维持细胞膜结构的完整性,进而抑制果肉呼吸强度及呼吸进程[25],这点与本文研究结果不一致。可能由于对呼吸跃变型果实,钙与果实呼吸作用大多呈负相关,钙处理能够推迟果实呼吸高峰出现并降低呼吸峰值;而对非呼吸跃变型果实,钙处理对果实呼吸的抑制作用主要体现在贮藏后期。
对于PPP途径关键酶活性,0.3%CaCl2+0.3%MgCl2和0.3%MgCl2处理对果肉G-6-PDH活性均呈现抑制作用,而对6-PGDH活性并未呈现规律性影响;0.3%CaCl2处理对6-PGDH活性在末期有一定的促进作用,对G-6-PDH无显著性影响。这与苏宏[26]的研究结果一致,不同浓度和种类的钙处理在常温条件下均显著提高了G-6-PDH活性,从而显著提高了PPP呼吸速率。本文结果表明,3个施肥处理对PPP呼吸速率的影响与对G-6-PDH关键酶活性的影响趋势基本一致。这说明呼吸速率主要受G-6-PDH调控,钙镁营养通过促进关键酶活性,进而影响该途径呼吸速率。
4 结论
“妃子笑”荔枝叶面喷施0.3%CaCl2+0.3%MgCl2可以克服果肉“退糖”现象,可能由于该处理提高了果肉水溶性镁含量,抑制了全钙含量,且在果实采前通过抑制G-6-PDH活性而抑制果肉PPP呼吸速率,表现出PPP和总呼吸速率降低,致使果肉生物合成速率受到抑制,减少果肉糖分呼吸消耗,使果肉糖含量未发生显著下降。0.3%CaCl2处理在果实采前促进了果肉全钙含量,进而促进了G-6-PDH关键酶活性,促进了PPP呼吸速率,从而未能抑制退糖。0.3%MgCl2处理虽然提高了果肉全镁含量,对G-6-PDH活性产生了抑制作用,但考虑镁元素可能通过EMP-TCA循环途径促进呼吸速率,从而对总呼吸速率无显著性影响。因此,0.3%CaCl2处理和0.3%MgCl2处理不能减缓果肉“退糖”现象。本文研究结果可为荔枝果实生长发育期叶面喷肥技术改良提供参考,也为荔枝果肉糖代谢理论问题的深入研究奠定了基础。
猜你喜欢
当代作家(2023年12期)2023-03-21
农业科技通讯(2023年1期)2023-02-12
作物学报(2022年1期)2022-11-05
河北科技师范学院学报(2022年2期)2022-08-26
农业工程学报(2022年7期)2022-07-09
——妃子笑
宝藏(2021年10期)2021-11-22
今日农业(2019年11期)2019-08-15
山东林业科技(2019年2期)2019-06-03
今古传奇·故事版(2017年16期)2017-09-22
中国果业信息(2017年4期)2017-05-09
