
耐热型西番莲品种“钦蜜9号”花器官发育特征与繁育系统研究
2024-01-31 04:17黄永才
中国南方果树 2024年1期
廖 芬,黄永才,桂 杰,陈 格,蒋 萍,杨 柳
(广西壮族自治区农业科学院/广西作物遗传改良生物技术重点实验室,南宁,530007)
西番莲又名百香果,是西番莲科(Passifloraceae)西番莲属(Passiflora)多年生常绿藤本植物,是热带特色优稀果树,原产南美洲中北部,主要生长在中南美洲、澳洲、非洲及亚洲[1]。近年来,随着我国消费者对西番莲的认可,鲜食、加工市场对西番莲的需求量急剧攀升,西番莲种植面积不断扩大,由于其生长期短(4~6个月),生产效益快,已成为我国热区(省、市)扶贫和乡村振兴的重要果树之一[2]。花是植物重要繁殖器官,其发育进程和生物学特征影响授粉和结果。特别是花粉活力与寿命、柱头可育性、花期相遇时间、杂交指数和异交倾向等都是植物有性繁殖过程的热点问题[3]。近年来天气异常,特别是高温已成为西番莲花芽败育或花器官败育的重要因素之一。研究西番莲花器官发育进程及育性,对解析西番莲耐热性适应机制和推进抗热杂交育种具有重要指导意义。近5年来,国内外西番莲研究主要集中于抗病选育[4-5]、栽培条件优化[6-7]和品质分析等方面[8]。在耐热性研究方面,主要针对花粉萌发进行相关研究。Hiroshi[9]等观察了温度对14个西番莲杂交栽培品种花粉萌发的影响,结果发现不同品种花粉萌发的最适温度不同,温度范围26~36 ℃,其中杂交种Minami-jujisei萌发最高温度为36 ℃,Passiflora.laurifolia的萌发温度最低,为26~28℃。紫果西番莲(Passifloraedulis)在昼夜温度33/28 ℃的高温条件下会生产小而低糖的果实,夏季夜间高温(30 ℃)还会对果皮的着色产生抑制作用[10]。田青兰等[11]以“台农1号”西番莲为研究对象,发现花期前20~35 d、开花当天及花后5~20 d出现高温(最高温度和平均温度分别高于36.19 ℃和26.24 ℃)会限制其花芽形成和着果。Atsushi等[12]比较不同温度(30~40 ℃)对两种日本栽培品种“Summer Queen”“Ruby Star”光合作用、蒸腾作用、呼吸率和叶绿体含量的影响发现,当温度超过35 ℃时,2种西番莲各生理指标参数随温度增加而下降,而高温环境下“Ruby Star”比“Summer Queen”更稳定。
我们团队分别于2019—2021年夏季高温季节(8月)对西番莲主产区,广西玉林市、贵港市、钦州市、防城港市进行产业调查,发现主栽品种“台农1号”“小黄金”“维蜜黄金”西番莲在高温季节(>35 ℃)着果率低于5%。针对当前国内缺乏耐高温西番莲品种问题,我们在大量收集国内外优异种质资源的基础上,长期开展高温胁迫(>35 ℃)条件下优异西番莲种质资源筛选,于2020年选育出耐高温、高产优质的黄金西番莲(P.edulisf.flavicarpa)新品种“钦蜜9号”,该品种平均单果质量69.17 g,纯甜无酸,糖酸比为12.7,可溶性固形物含量19.4%,不耐冷但耐高热,在夏季>35 ℃高温连续10 d以上仍可正常开花结果,而其他黄果西番莲在此温度条件下极少开花结果,说明该品种花器官发育可以耐受高温。目前西番莲适应高温的花器官发育过程鲜见报道。本试验观察“钦蜜9号”花器官不同发育阶段性状,分析其花粉活力、柱头可育性等,解析耐高温品种繁育特点和耐热机制,为创制耐热型西番莲杂交群体和新品系选育提供参考。
1 材料与方法
1.1 材料
耐热型西番莲品种“钦蜜9号”来自广西壮族自治区农业科学院里建科学研究基地西番莲资源圃,北纬23.20°,东经18.16°,海拔200 m,年日均最高气温17~37 ℃,日均低温11~27 ℃,每年6—10月日均高温33~37 ℃,年降水量1 100~1 300 mm,亚热带季风气候。
试验于2021—2022年7—10月开展,此期是“钦蜜9号”持续开花期,试验地夏季35~37 ℃高温主要集中在7—8月,2021年7—8月35 ℃ 以上高温天气共35 d,2022年7—8月共35 d;其中2021年7月12—22日持续11 d,2022年7月20日至8月4日持续16 d。9—10月仍有8~13 d 35 ℃ 以上高温天气(见图1)。
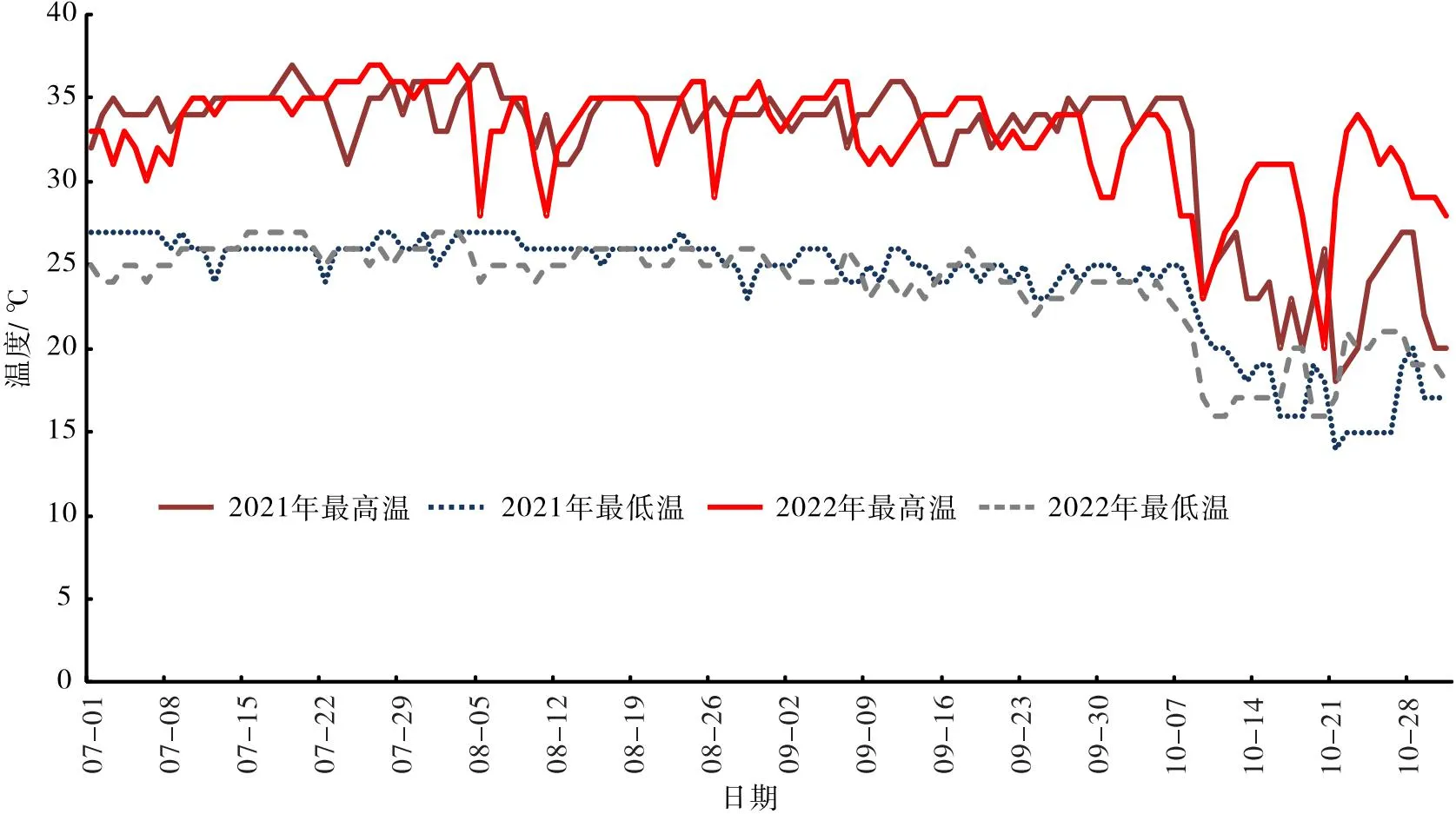
图1 2021—2022年7—10月供试果园温度情况
在“钦蜜9号”二级蔓花序形成后,选取花序完整、植株健壮,生长良好无病株的枝蔓,标记不同发育阶段花朵进行观测和记录;根据生长日期,采集不同发育阶段花朵装在密封袋中带回实验室,进行室内观察和数据测定。
1.2 花器官特征测定
不同发育阶段的花蕾表型测定:分别取不同发育阶段花蕾20个,采用游标卡尺测定花蕾横径、纵径,花蕾纵径为萼筒基部到花冠顶部距离,花蕾横径为最宽位处的距离;将花蕾各部分分解,在立体解剖镜下观察花器官内部结构发育变化情况。
完全开放的花器官表型测定:采用游标卡尺测定完全打开后的花器官直径,萼片直径和长度,花瓣长度和宽度,萼筒直径和高度,花药柄长度,花药长度和宽度,柱头长度和宽度。
1.3 花粉数量测定
取裂蕾期前1~2 d花药未裂开时、开花时、开花第1天和第3天的花药,放入培养瓶中,置于有硅胶的干燥皿中待花药干燥后散落于管中。待花粉充分散出后,每管滴入1%偏6磷酸溶液1 mL,加盖充分摇匀,使花粉悬浮充分,XB-K-25型血球计算板统计花粉数量,重复6次[13],每个花药花粉数量(个)=(80个小方格花粉粒总数×400×10 000×稀释倍数)/80 。
1.4 花粉活力和萌发能力测定
采用I2-IK染色法测定花粉活力,取裂蕾期前花药未裂开时、开花时、开花第1天和第3天花药花粉,放在载玻片上滴入0.5% I2-IK溶液1~2滴,显微镜下观察统计,每个发育期取花药6个,每个花药观察5个视野数据,以着色花粉数量作为其活力指标。花粉活力(%)=有活力的花粉数/花粉总数×100[13]。
花粉萌发能力测定采用培养基萌发法,参考刘洁云等[14]的方法,取不同发育阶段花药花粉,放在有花粉培养基的凹面载玻片上,26 ℃下黑暗培养2 h,显微镜下检测花粉管长度大于花粉直径的花粉数量,每个发育期取花药6个,每个花药观察5个视野数据,花粉萌发率(%)=萌发花粉数/花粉总数×100。
1.5 柱头可授性的测定
取不同发育阶段花朵柱头置于培养瓶中,加入1%联苯胺∶3%过氧化氢∶水=4∶11∶22的反应液,柱头需全泡在反应液中,黑暗处理25 min,观察柱状颜色和气泡产量数量判断柱头可育性。柱头全变粉色,产生大量大气泡的可育性最强,标记为+++;柱头全变粉色,气泡量较多的可育性较强,标记为++;柱头一半以上染成粉色,气泡量少为具有可育性,标记为+;柱头粉色区域小于1/3,气泡极少或没有,部分有可育性,标记为-/+;柱头无粉色或无气泡,无可育性,记为-[13]。
1.6 异交指数(OCI)和花粉/胚珠(P/O)测定
随机挑选完全打开花朵20朵,统计异交指数(OCI);同时解剖子房,观察记录胚珠数,计算花粉胚珠比(P/O),P/O =花粉总数/胚珠数。根据Dafni的异交指数标准[15]和Cruden的花粉/胚珠标准[16]判断“钦蜜9号”的繁育系统类型,具体为花朵直径<1 mm为0分,1~2 mm为1分,2~6 mm为2分,>6 mm,3分;雌雄蕊情况为花药开裂,柱头有可授性,或雌蕊先熟为0分,雄蕊先熟为1分,柱头与花药在同一高度为0分,空间分离为1分。OCI=花朵直径得分+雌雄蕊得分;OCI为0时,闭花受精;OCI为1时,专性自交;OCI为2时,兼性自交;OCI为3时,自交亲和,有时需要传粉;OCI为4时,异交,部分自交亲和,异交需要传粉者。P/O=2.7~5.4为专性自交,P/O=18.1~38.0为兼性自交,P/O=2 588~396 244.7为兼性异交,P/O=2 108~195 525为专性异交。
1.7 传粉方式试验
择标记好的花朵50朵,分别进行自然授粉、人工异花授粉、人工自花授粉和不授粉等4种处理,统计各处理着果数,计算着果率。
1.8 数据分析
采用Excel 2007软件整理数据,SPSS 19.0软件进行方差分析。
2 结果与分析
2.1 花器官各发育阶段特征
“钦蜜9号”是较耐热黄果西番莲品种,经过多年调查发现,在夏季温度超过35 ℃以上时仍正常花芽分化并开花结果。“钦蜜9号”一般移植后40~50 d进入生殖期,可持续开花到当年10月,花朵开放时间一般为13:00—15:00,单花开放时间约24 h。“钦蜜9号”进入生殖生长期后,在枝条叶片同节位卷须基部出现点状突起的花芽,花芽分化完成后慢慢长大,苞片生长较快,1周后拔开苞片可以看到花蕾。苞片中可见点状突起花蕾时记为现蕾0 d,整个发育进程大致可分为:现蕾期、花冠发育期、雌雄蕊成熟期、裂蕾开花期和谢花期(见图2),持续时间约25~27 d。

注:1—4为现蕾期(现蕾0~6 d),5—9为花冠发育期(现蕾后7~12 d),10—12为雌雄蕊成熟期(现蕾后13~20 d),13-15为裂蕾开花期(现蕾后21~22 d),16—17为谢花期(现蕾后23 d和25 d)。
2.2 开花习性和花器官形态特征
“钦蜜9号”花开放可分为裂蕾、萼片打开、花瓣打开、雌雄蕊散开、花药向上翻转、柱头向下接触花药、授粉等阶段,花开放时的形态见图2。
花朵完全开放时,即完全成熟花直径平均值6.86 cm,最大值8.12 cm,最小值5.8 cm。萼片长平均值3.93 cm,最大值4.9 cm,最小值3.38 cm;宽平均值1.42 cm,最大值1.51 cm,最小值1.37 cm。薄膜状花瓣长平均值4.23 cm,最大值5.01 cm,最小值3.33 cm;宽平均值1.44 cm,最大值1.54 cm,最小值1.38 cm。丝状花瓣长平均值2.43 cm,最大值2.68 cm,最小值2.03 cm。萼筒直径平均值1.05 cm,最大值1.32 cm,最小值0.82 cm;高度平均值1.45 cm,最大值1.74 cm,最小值1.21 cm。雄蕊,花药柄长平均值0.94 cm,最大值1.02 cm,最小值0.9 cm。花药长平均值1.1 cm,最大值1.22 cm,最小值0.97 cm;宽平均值0.54 cm,最大值0.59 cm,最小值0.51 cm。雌蕊中,子房高度平均值0.6 cm,最大值0.67 cm,最小值0.55 cm;直径平均值0.44 cm,最大值0.46 cm,最小值0.41 cm。柱头宽平均值0.61 cm,最大值0.66 cm,最小值0.56 cm;长平均值1.43 cm,最大值1.58 cm,最小值1.36 cm。
从图2可以看出,“钦蜜9号”花器官由花梗、苞片、萼片、薄膜状花瓣、丝状花瓣、雄蕊和雌蕊组成。苞片3片,长1.7~2.0 cm,宽1.5~1.8 cm,叶脉因有花青素而呈褐红色斑点;萼片5枚,平均长度3.93 cm,平均宽度1.42 cm,外面为绿色,从基部向上颜色由浅绿色逐渐加深,至顶部为深绿色,每片萼片外部中间有条突起的脊;每片苞片和萼片上着生有数个小颗粒凸起的蜜腺。薄膜状花瓣白色,5枚,平均长度4.23 cm,平均宽度1.44 cm;丝状花瓣无数枚围成一圈,尾部略卷曲,靠近花萼筒的约1/2长度为深紫色,外部约1/2长度为白色;花瓣基部与萼片底部联合形成圆柱形萼筒,萼筒平均直径1.05 cm,高度1.45 cm,萼筒内部有蜜腺。雄蕊5枚,长度一致,花药黄色,长方形囊状。子房上位,顶部柱头3枚,绿色,柱头分裂为二,呈蹄状结构。花药和柱头基部均着生于子房顶部,柱头高度略高于花药3~5 mm。
裂蕾后,花在当天13:00—15:00开放。开放时,花部外面包裹的5片绿色萼片逐片打开,接着白色薄膜花瓣打开,连同里面一圈丝状花瓣也开始展开;5枚雄蕊缓慢向四周散开,此时花药已裂开,散开后开裂的花药囊转头向下;雄蕊散开后,原本聚拢在一起的雌蕊的3个柱头也散开,此时整朵花完全开放。花完全开放后,原来转头向下的花药会慢慢地反转向上,同时柱头也向下弯,向花药靠拢,最后碰在一起,花粉授到柱状上,授粉完成,在开放过程中可以看到花粉也会黏到柱头。授粉结束后1~2 d,可以观察到子房明显膨大,整朵花的花被部分又开始收拢,花药、柱头、花瓣、萼片等开始慢慢枯萎,进入谢花期,此后是果实生长期。
现蕾期:现蕾后3~4 d,花蕾直径约1 mm,除丝状花瓣外其他器官均已形成,雄蕊和柱状为嫩绿色,花药内部腔室已形成,花药与子房紧密靠近。现蕾后6 d时,花蕾直径约2 mm,花蕾高度约至苞片的1/2,4个花药室已清晰可见,并充满内容物;萼片开始增厚,子房横切面呈三角形,内膜可以看到一些小突起,子房与花药有明显的空隙。
花冠发育期:分3个时期,一是现蕾后7~8 d,花蕾横径0.4~0.8 cm,纵径0.5~1.0 cm,纵横径比1.25,花蕾高度与苞片长度齐平,丝状花瓣开始出现,最初为白色,1~2 d后基部开始变紫,此时萼片明显变厚,胚珠已明显可见,花药颜色转为黄色,柱头深绿色,已有可授性。花药室未见明显内容物,子房横切面从三角形慢慢变为圆形,子房与花药的空隙变得更大。二是现蕾后9~11 d,花蕾横径0.8~1.4 cm,花蕾纵径1.0~2.2 cm,纵横径比1.25~1.57,花蕾略超出苞片2~3 mm,丝状花瓣基部变成紫色。花药室充满内容物,子房长大,胚珠已明显可见。三是现蕾后12~13 d,花蕾横径1.4~1.8 cm,花蕾纵径2.2~3.0 cm,纵横径比1.57~1.67,苞片基本定形,苞片高至花蕾1/2位置,各器官不断长大,子房与花药之间已无明显空隙。
雌雄蕊成熟期,分3个时期,一是现蕾后14~17 d,花蕾横径1.8~2.3 cm,花蕾纵径3.0~3.8 cm,纵横径比1.65~1.67,苞片高至花蕾的1/3位置,各器官不断长大,花药室充满内溶物,未看到成熟花粉粒。二是现蕾后18~19 d,花蕾横径2.3~2.5 cm,花蕾纵径3.9~4.7 cm,纵横径比1.69~1.88,苞片与萼筒同高,花蕾饱满,花药未裂开,但已可以看到成熟花粉,此时萼筒基本定形,胚珠不断长大并填满子房腔室。三是现蕾后20~21 d,花蕾横径2.5~2.6 cm,花蕾纵径4.8~4.9 cm,纵横径比1.88~1.92,花药已开裂,花蕾纵横径基本定形。
裂蕾开花期,现蕾后21~22 d,花蕾横径2.6 cm,花蕾纵径4.9 cm,纵横径比1.88,接近中午时开始裂蕾,13:00—15:00单花完全开放,开放后自花授粉约在1 h内完成,异花授粉主要通过昆虫如蜜蜂传粉。
谢花期,现蕾后22~27 d,授粉后2 d可以看到子房明显膨大,花药和子房开始有一点萎蔫,授粉后5 d时,子房直径已经可以达到4~5 cm,此时大部分花的花药和子房开始萎蔫或干枯(见图3)。
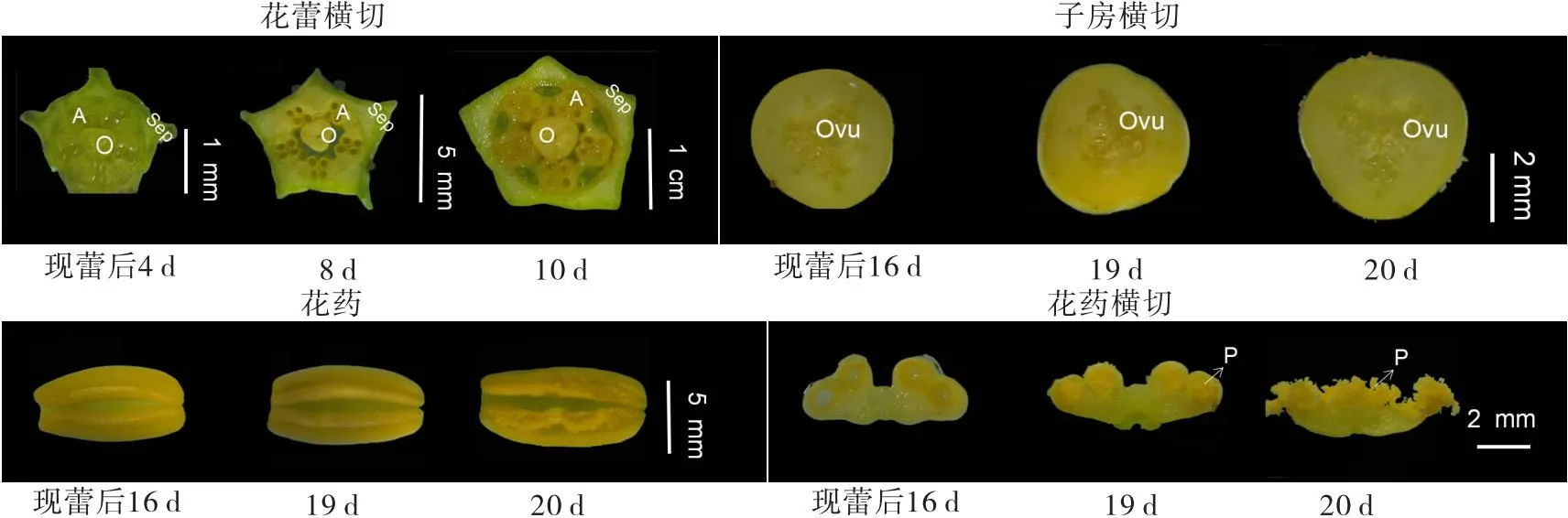
注:O为子房,A为花药,Sep为萼片,P为花粉,Ovu为胚珠。
2.3 不同发育阶段花粉活性变化
“钦蜜9号”在开花前1~2 d(现蕾后19~20 d)时花粉粒开始成熟,培养基培养30 min后开始陆续萌发。从表1可以看出,‘钦蜜9号’花粉粒数量在开花当天最多,有7.94×105个/花药 ,开花后花粉数量大量减少,授粉后第3天时花粉数量仅是开花当天的1/10左右。花粉活性均在80%以上,开花当天和授粉后1 d花粉活性分别达到90.11% 和92.17%。花粉萌发率在开花当天达到最高,为67.69%,授粉后1 d萌发率下降至24.9%,授粉后3 d萌发率仅有19.55%。花粉形态完整饱满,畸形率非常低,在1.04%以下。
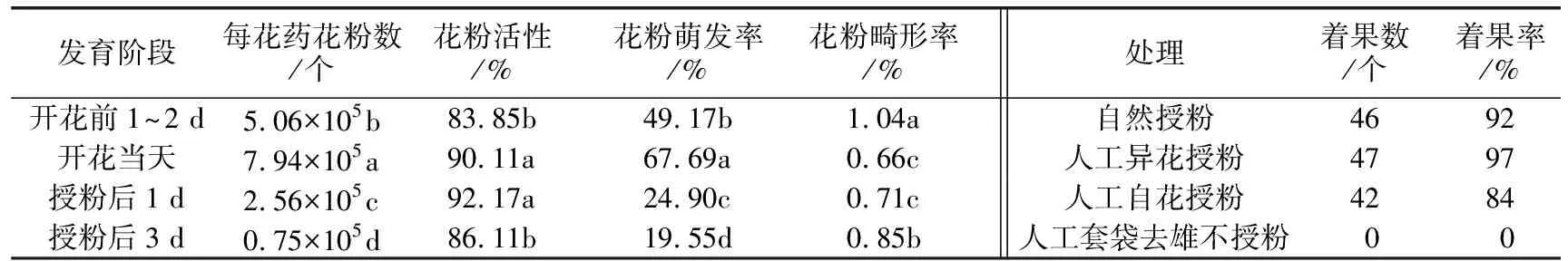
表1 “钦蜜9号”黄果西番莲不同发育期花粉数量、活性及不同授粉处理着果情况
2.4 柱头可授性
花蕾长至直径约4~5 mm时,即现蕾后7~8 d,柱头明显变绿色,此时柱头已有可授性,随着花器官不断发育和雌雄蕊成熟,柱头产生的气泡不断增加,可授性不断增强。现蕾后12 d时柱头可授性已较强,开花前3 d至开花前1 d(现蕾后18~21 d),柱头可授性达到最强,开花当天可授性略微有所下降,但仍较强。授粉后柱头可授性开始下降,至授粉后3 d柱头可受性已大大减弱,但仍有可授性,此时柱头已开始变干瘪,颜色变成褐黄色。具体情况为现蕾后8、10、12、14、16、18、19、20、21、22、25 d柱头可授性分别为+、+、++、++、++、+++、+++、+++、+++、+、-/+(+越多可授性越强)。
2.5 繁育系统评判
经观察,“钦蜜9号”柱头位置平均高于花药3~5 mm,雌雄蕊在空间位置上有分离,柱头在花瓣打开前就有可授性,花药开裂时间在裂蕾前1 d。根据 Dafni的标准评分,“钦蜜9号”的OCI为4,推断其繁育系统为异交型,部分自交亲和,异交需要传粉者。花粉/胚珠为3 163,属于兼性异交类型。授粉试验结果表明,“钦蜜9号”自花和异花均可育,且自交与杂交亲和性均较高(见表1),验证了前面的繁育类型。
3 讨论
3.1 “钦蜜9号”花器官发育进程特征
通过花器官发育过程中各阶段表型性状与内部器官发育进程相关联分析,可以从花蕾外观快速判断花蕾内部各器官的生长发育阶段,这对杂交育种工作中选择适合的杂交时间非常重要。本研究结果表明,“钦蜜9号”在开花前1~2 d,花药未裂时花粉粒已成熟,如果需要进行异花授粉,应该在此时进行。“钦蜜9号”在亚热带低海拔地区高温夏季仍可连续开花,现蕾后19~20 d花粉粒开始成熟,一般选择温度较高的午后开花,可能是其开花需要较多热量。在广西南宁高温夏季,该品种花器官发育未见有败育阶段,花粉数量、花粉活性及柱头可授性均正常,说明其花器官发育过程较耐高温,这与许多耐高温作物花器官性状相似[17-19]。植物开花后可自花授粉亦可异花授粉,且两种传粉方式的亲和性均较高,这可能是其对环境适应的一种调节特性,即可以保证其近交优势,又可以增加遗传多样性[20-21]。花粉数量及其活性、柱头可育性、花粉活性与柱头可育性是否同期等均会影响植物授粉效果和结果率[22-23]。本研究结果表明,“钦蜜9号”花粉数量和活性非常高,颗粒饱满,畸形率较低,开花时花粉萌发率也较高,同时柱头可授性在花粉活性和萌发最高时也达到最强,并且在时期上有重叠,创造了其自花授粉的良好条件。本研究也发现,“钦蜜9号”在开花前1~2 d时,花粉粒已成熟同时具有较高活性和萌发率,且花药和柱头存在空间一定距离的分离,这种特性使杂交处理操作方便[24-25]。因此,“钦蜜9号”杂交既可以作为母本,也可作为父本。
3.2 “钦蜜9号”繁育系统特征及利用
根据Cruden和Dafni的标准,“钦蜜9号”繁育系统属于兼性异交,繁育方式为异交型,部分自交亲和,异交需要传粉者,为自交和异交混合交配系统;混合交配系统一般被认为是植物适应环境的最适交配体系[24-26],人工处理的结果也验证这一结论。本研究发现,“钦蜜9号”自交和异交亲和性均较高,这与其他异交植物不同[27-28];一般来说异交植物提高异交率,自交亲和力会较低,或通过花粉活性与柱头可授性不同期来避免自交发生,“钦蜜9号”自交和异交高亲和性比较特殊。此外,本研究结果发现,“钦蜜9号”花粉和柱头在开花5 d内虽然活性和可育性大多下降 ,但还具有一定育性,因此杂交后一定做好防护工作,以防其他花粉再次授到柱头上。
“钦蜜9号”兼具有性繁殖与无性繁殖,这可能是植物应对多变环境的生殖补偿机制,可以保证其后代延续[16,29]。本研究结果也发现,“钦蜜9号”柱头可授性和花粉萌发率在开花后2~3 d大幅下降,开花时间一般为24 h左右,这会影响其有性繁殖的概率,生产实践中也发现开花后如遇大风大雨天气,授粉率和结果率会大大降低,因此,可以利用其杂交亲和性高的特点,先通过有性杂交聚合不同品种优良性状,创制优良种质,筛选优异杂交后代后,再通过无性繁殖快速扩繁获得大量优异种群,再进一步筛选后代优良株系,以获得性状稳定的优株。
4 结论
“钦蜜9号”耐热性体现在其花器官繁育系统发育进程不受高温影响,花粉数量多、活性强,柱头可育性强,花粉活性与柱头可育性同步性高度一致,保证了其高结果率。其自交与异交亲和性均较高,杂交时既可作父本,也可作母本。本研究主要针对其花器官发育进程特征和繁育系统特性进行研究,仍需对该品种耐热性的生理生化过程和分子机制开展深入研究,以期能全面挖掘“钦蜜9号”的耐热机制,为西番莲耐热抗性育种奠定基础。
猜你喜欢
当代作家(2023年12期)2023-03-21
云南化工(2021年10期)2021-12-21
中国瓜菜(2021年5期)2021-07-19
西北农林科技大学学报(自然科学版)(2021年5期)2021-05-31
中国果树(2020年2期)2020-07-25
农民致富之友(2020年6期)2020-04-08
中国瓜菜(2019年2期)2019-07-02
草堂(2018年11期)2018-11-13
世界热带农业信息(2018年11期)2018-01-17
长江蔬菜·学术版(2017年6期)2017-07-27
