
不同暖季型草坪草品种对盐碱地土壤理化性质和微生物群落的影响
2024-01-30 04:28李晓慧沈泽宇邹景文隋德宗陈静波郭海林
植物资源与环境学报 2024年1期
姚 祥, 李晓慧, 沈泽宇, 邹景文, 隋德宗, 陈静波,①, 郭海林,①
〔1. 江苏省中国科学院植物研究所(南京中山植物园) 江苏省植物资源研究与利用重点实验室, 江苏 南京 210014;2. 江苏省林业科学研究院, 江苏 南京 211153〕
土壤盐渍化问题引起了学者广泛关注。据中国第2次土壤普查资料,中国盐渍土面积3.47×107hm2,相当于耕地面积的1/3[1]。江苏沿海滩涂总面积6.53×105hm2左右,约占中国滩涂总面积的1/4[1]。盐碱地是重要的后备土地资源,但由于沿海滩涂盐碱含量高,重度盐碱地含盐量高达0.7%~1.0%,土壤养分匮乏,一般植物很难生长[2]。通过生物措施改良盐碱地是目前盐碱地改良和利用的发展战略。关于植被对盐碱地土壤理化性质和土壤微生物影响的研究较多,有研究结果表明种植耐盐植物可显著改善土壤理化性质和土壤酶活性[3],不同植物根际的微生物群落结构不同[4],无植被覆盖盐碱地土壤真菌和放线菌丰富度低于有植被覆盖盐碱地土壤,且不同植物覆盖土壤的微生物群落存在差异[5]。
草坪草因其强适应性,在盐碱地绿化中得到广泛应用。早期关于冷季型草坪草在盐碱地中适应性评价的研究较多。例如:李发明[6]在甘肃河西盐碱地区进行了毛稃羊茅〔Festucarubrasubsp.arctica(Hackel) Govoruchin〕的种植研究,发现该种表现出强适应性;王海生等[7]在山东东营盐碱地区进行了多年生黑麦草(LoliumperenneLinn.)、高羊茅(FestucaelataKeng ex E. B. Alexeev)和草地早熟禾(PoapratensisLinn.) 3种草的40个品种的适应性研究,发现这3种草均有少数品种表现良好;刘金荣等[8]在甘肃张掖高寒盐碱地区进行了21种冷季型草坪草的适应性研究,发现草地早熟禾表现最好;汤巧香[9]在天津盐碱地区进行了冷季型草坪草6属11种122个品种的适应性研究,发现草地早熟禾、高羊茅和多年生黑麦草具有良好适应性。此外,部分暖季型草坪草也具有强耐盐性,为天然的盐生植物,如大穗结缕草(ZoysiamacrostachyaFranch. et Sav.)[10]和海雀稗(PaspalumvaginatumSw.)[11]等。已有研究结果表明:海雀稗品种‘P006’、沟叶结缕草〔Zoysiamatrella(Linn.) Merr.〕品种‘Z014’和结缕草(ZoysiajaponicaSteud.)品种‘Z080’等在江苏如东沿海滩涂重度盐碱地上表现良好[12]。海雀稗、狗牙根〔Cynodondactylon(Linn.) Pers.〕和结缕草等在美国盐碱地区应用广泛[13]。Zong等[14]在中国南海礁沙上进行了海雀稗、巴哈雀稗(P.notatumFlüggé)、狗牙根、细叶结缕草〔Zoysiapacifica(Goudswaard) M. Hotta et S. Kuroki〕和钝叶草(StenotaphrumhelferiMunro ex Hook. f.) 的适应性研究,发现狗牙根表现最好,且狗牙根根际细菌物种丰富度显著高于其他4种草。但关于暖季型草坪草对盐碱地土壤理化性质和微生物群落影响的研究尚不全面,不利于盐碱地土壤的进一步改良。
目前,在江苏及周边地区广泛应用的暖季型草坪草主要为从美国引进的Tif系列杂交狗牙根[15-16]。这些草坪草虽品质良好,但是耐盐性较差,在重度盐碱地上需换土才能成功建植[17]。针对以上情况,本项目组从2000年开始进行耐盐草坪草新品种选育,建立了暖季型草坪草耐盐评价体系[18-19],选育出狗牙根品种‘阳江’(‘Yangjiang’)、杂交结缕草(Z.japonica×Z.tenuifolia)品种‘苏植1号’(‘Suzhi No. 1’)和杂交结缕草(Z.sinica×Z.matrella)品种‘苏植3号’(‘Suzhi No. 3’)等一系列坪用价值高、耐盐性强、耐粗放管理的草坪草新品种[17]。选育的耐盐草坪草新品种和盐碱地草坪相关技术体系已在江苏的盐城大丰、盐城东台、南通如东以及山东潍坊等盐碱地区的草坪生态修复建设中得到大面积应用。基于以上研究,本研究分析了在江苏盐城盐碱地中种植优质耐盐新品种‘阳江’和‘苏植3号’对盐碱地土壤理化性质和土壤微生物群落的影响,以期为暖季型草坪草在盐碱地生态修复中的应用提供科学依据。
1 研究地概况和研究方法
1.1 研究地概况
研究地位于江苏省盐城市大丰麋鹿国家级自然保护区江苏省林业科学院沿海造林试验站内,地理坐标为东经120°49′~120°52′、北纬33°00′~33°20′,海拔2~3 m。该地区属亚热带与暖温带过渡地带,具有海洋性气候和季风性气候,年平均气温13.7 ℃~14.5 ℃,年平均降水量980~1 100 mm,降水集中在6月至9月。研究区土壤粉沙质,盐渍化,0~15 cm土层土壤的平均盐度为1.4%,主要植物有棒头草(PolypogonfugaxNees ex Steud.)、马齿苋(PortulacaoleraceaLinn.)、碱蓬〔Suaedaglauca(Bunge) Bunge〕和盐角草(SalicorniaeuropaeaLinn.)等。
1.2 材料
供试材料为江苏省中国科学院植物研究所自主选育的暖季型多年生草坪草新品种‘阳江’和‘苏植3号’,二者均通过匍匐茎繁殖,选当年生草种在试验区种植。
1.3 方法
1.3.1 试验设计 于2021年6月,在试验区内对‘阳江’和‘苏植3号’进行盐碱地生态修复示范种植,以原生境草地(以棒头草为主)为对照,示范地和对照地的水肥管理措施相同。每个草种示范地面积2 000 m2,包含4个区域,每个区域约500 m2。对照地4块,每块面积120 m2。草种种植1年后,在各示范地和对照地内随机选取1个面积2 m×2 m的样地,即每个草种和对照均4次重复。在各样地内随机选取3个点,3个点内植株地下部生物量的均值即为该样地植株地下部生物量,另随机取3个点土壤样本混为1个样本用于后续土壤理化性质和微生物群落分析。
1.3.2 植株地下部生物量和土壤理化指标测定 去除地表植物体和枯枝落叶,使用口径5 cm的不锈钢土钻采集0~15 cm土层土壤,土钻内径面积(19.625 cm2)即为土壤取样面积,洗去土壤后获得植株地下部,于80 ℃烘干至恒质量,使用千分之一电子天平称量植株地下部干质量,并根据公式“植株地下部生物量=土钻中植株地下部干质量/土钻内径面积”计算植株地下部生物量。同样使用土钻采集0~10 cm土层土壤,过筛(孔径2 mm)去除枯枝落叶等杂质后进行土壤理化性质和微生物群落分析。采用重铬酸钾氧化法[20]测定土壤有机质含量;使用雷磁PHS-3C pH计(上海仪电科学仪器股份有限公司)测量土壤pH值;使用FieldScout EC110便携式电导率仪(美国Spectrum Technologies公司)测量土壤电导率。
1.3.3 土壤微生物DNA提取及测序 每个土壤样本取0.5 g,使用天根土壤DNA提取试剂盒〔天根生化科技(北京)有限公司〕提取DNA,使用NanoDrop 2000 分光光度计(美国Bio-Rad Laboratories公司)检测DNA质量,然后使用稀释后的基因组DNA (1 ng·μL-1)进行PCR扩增。使用引物F515(5′-GT GCCAGCMGCCGCGGTAA-3′)和R806(5′-GGACTA CHVGGGTWTCTAAT-3′)扩增细菌16SrRNA V4区基因[21],使用引物ITS5-1737F(5′-GGAAGTAAAAG TCGTAACAAGG-3′)和ITS2-2043R(5′-GCTGCGT TCTTCATCGATGC-3′)扩增真菌ITS1区基因[22],然后使用质量体积分数2%琼脂糖凝胶电泳检测PCR产物,最后使用MinElute凝胶回收试剂盒(德国Qiagen公司)回收目的条带。回收产物委托诺禾致源生物科技有限公司进行扩增子测序。
1.3.4 土壤微生物生物信息学分析 使用fastp软件对得到的raw tag进行质控,得到高质量clean tag,之后使用Vsearch软件将clean tag与数据库比对去除嵌合体[23],最终得到有效数据(effective tag)。使用QIIME2软件对得到的effective tag降噪并过滤掉丰富度小于5的序列,从而获得最终的扩增子序列变异(ASV)以及特征表。使用Silva138.1数据库注释16SrRNA V4区域[24],使用UNITEv8.2数据库注释ITS1区域[25],从而得到每个ASV的物种信息[26];每个ASV比对到的该物种序列数即为该物种的丰富度,每个物种的序列数与该样本中所有物种总序列数的比值为该物种的相对丰富度。使用FAPROTAX软件预测细菌功能[27],使用FunGuild软件预测真菌功能[28-29];使用R 4.0.3软件计算各样本在某个分类上的Z值,计算公式为Z值=(某一样本在该分类上的相对丰富度-所有样本在该分类上的平均相对丰富度)/所有样本在该分类上的标准差,并进行聚类分析,细菌和真菌均展示前30个注释功能。原始数据上传至NCBI中的SRA数据库(https:∥www.ncbi.nlm.nih.gov/),BioProject号为PRJNA962702。
1.4 数据处理和分析
使用SPSS 16.0软件对植株地下部生物量以及土壤的有机质含量、pH值、电导率以及细菌和真菌的Shannon-Wiener多样性指数进行方差齐质性检验(Levene检验,P>0.05)后进行单因素方差分析,检验组间差异显著性。使用QIIME2软件中的Anosim函数分析草坪草种植样地土壤中细菌和真菌群落结构与对照样地间的差异显著性[30];使用QIIME2软件计算细菌和真菌的Shannon-Wiener多样性指数以及组间差异显著的功能细菌和真菌(固氮细菌、丛枝菌根真菌和粪腐生菌)的相对丰富度。对组间差异显著的功能细菌和功能真菌相对丰富度的组间差异显著性进行非参数wilcox秩和检验。使用R 4.0.3软件对细菌和真菌多样性以及功能菌丰富度与植株地下部生物量和土壤理化指标间进行Pearson相关性分析。
2 结果和分析
2.1 植株地下部生物量和土壤理化指标的比较
不同暖季型草坪草品种种植样地的植株地下部生物量和土壤理化指标见表1。结果显示:‘阳江’和‘苏植3号’种植样地植株地下部生物量分别为206.847和265.669 g·m-2,均显著高于对照样地(39.204 g·m-2);‘阳江’和‘苏植3号’种植样地土壤有机质含量分别为12.122和10.378 g·kg-1,均显著高于对照样地(8.418 g·kg-1);‘阳江’和‘苏植3号’种植样地土壤pH值分别为pH 8.15和pH 8.18,均显著低于对照样地(pH 8.56);‘阳江’和‘苏植3号’种植样地土壤电导率分别为13.04和14.33 μS·m-1,均显著低于对照样地(93.53 μS·m-1)。
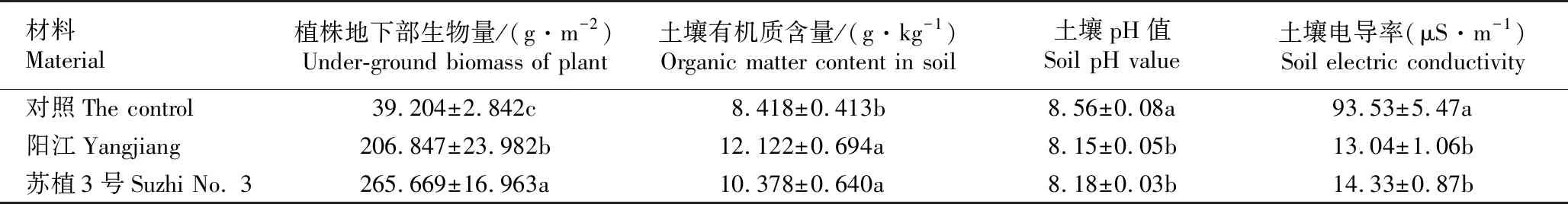
表1 不同暖季型草坪草品种种植样地的植株地下部生物量和土壤理化指标1)
2.2 土壤细菌和真菌群落结构的比较
Anosim分析结果显示:‘阳江’和‘苏植3号’种植样地土壤中细菌群落结构与对照样地差异显著,P值分别为0.030和0.035;‘阳江’和‘苏植3号’种植样地土壤中真菌群落结构与对照样地也差异显著,P值分别为0.030和0.026。
结果(表2)显示:‘阳江’和‘苏植3号’种植样地土壤中细菌的Shannon-Wiener多样性指数分别为10.473和10.433,均显著高于对照样地(9.848);‘阳江’和‘苏植3号’种植样地土壤中真菌的Shannon-Wiener指数分别为5.250和5.133,均高于对照样地(4.744),但差异不显著。‘阳江’和‘苏植3号’种植样地土壤中固氮细菌的相对丰富度分别为0.003 10和0.002 50,均显著高于对照样地(0.000 90);‘阳江’和‘苏植3号’种植样地土壤中丛枝菌根真菌的相对丰富度分别为0.007 34和0.011 91,均显著高于对照样地(0.000 52);‘阳江’和‘苏植3号’种植样地土壤中粪腐生真菌的相对丰富度分别为0.013 91和0.007 18,均显著高于对照样地(0.000 30)。
结果(图1和图2)还显示:总体上看,‘阳江’和‘苏植3号’种植样地土壤中相同功能细菌类群的丰富度较为接近,二者与对照样地土壤中相同功能细菌类群的丰富度差异均较大,说明‘阳江’和‘苏植3号’种植样地土壤中功能细菌类群关系较近,二者与对照样地的关系均较远;对照样地以及‘阳江’和‘苏植3号’种植样地间土壤中相同功能真菌类群的丰富度差异均较大,说明3种样地土壤中功能真菌类群关系均较远。

CK: 对照The control; YJ: ‘阳江’‘Yangjiang’; SZ: ‘苏植3号’‘Suzhi No. 3’. US: 未定义腐生Undefined saprotroph; Li: 地衣型Lichenized; WS: 木腐生Wood saprotroph; Un: 未指定功能Unassigned; FP: 寄生真菌Fungal parasite; AP: 动物病原Animal pathogen; En: 内生Endophyte; PP: 植物病原Plant pathogen; DS: 粪腐生Dung saprotroph; LeS: 叶腐生Leaf saprotroph; SS: 土壤腐生Soil saprotroph; Ec: 外生菌根Ectomycorrhizal; AM: 丛枝菌根Arbuscular mycorrhizal; LiS: 凋落物腐生Litter saprotroph; PS: 植物腐生Plant saprotroph.
2.3 微生物多样性和功能微生物丰富度与植株地下部生物量和土壤理化指标的相关性分析
结果(表3)显示:细菌Shannon-Wiener多样性指数与植株地下部生物量和土壤有机质含量呈显著正相关,与土壤pH值和土壤电导率呈极显著负相关,相关系数分别为0.671、0.662、-0.830和-0.789;固氮细菌相对丰富度与植株地下部生物量呈极显著正相关,与土壤pH值和土壤电导率呈显著负相关,相关系数分别为0.734、-0.621和-0.705;丛枝菌根真菌相对丰富度与植株地下部生物量呈显著正相关,相关系数为0.653;粪腐生真菌相对丰富度与土壤pH值呈显著负相关,相关系数为-0.616。说明植株地下部生物量和土壤理化性质明显影响细菌多样性,植株地下部生物量以及土壤的pH值和电导率明显影响部分功能细菌的丰富度,植株地下部生物量和土壤pH值明显影响部分功能真菌的丰富度。

表3 暖季型草坪草种植样地土壤中微生物多样性和功能微生物丰富度与植株地下部生物量和土壤理化指标的相关性分析
3 讨论和结论
3.1 暖季型草坪草地下部生物量及其对土壤有机含量质的影响
‘阳江’和‘苏植3号’为优良的暖季型草坪草品种,具有发达的地下茎和根系系统,在0~30 cm土层内盘根错节形成根结皮,具有非常强的水土保持能力[31-32]。本研究中,‘阳江’和‘苏植3号’种植样地的植株地下部生物量均显著高于对照样地,且土壤有机质含量也显著高于对照样地。沿海滩涂地区盐碱含量高,养分匮乏,有机质含量偏低,土壤肥力低,一般绿化植物很难生长[33]。本项目组前期研究结果表明‘阳江’和‘苏植3号’可在重度盐碱地区种植并生长良好[17],植株地下部生物量和土壤有机质含量的增加有利于土壤养分的增加,为植物生长提供需要的养分,增加土壤肥力,促进团粒结构的形成以及土壤微生物的生长[33-34]。本研究进一步证明了‘阳江’和‘苏植3号’具有较强的耐盐性以及高的生长密度,在江苏滨海盐碱地种植这2个品种有利于增加盐碱地土壤肥力,在盐碱地生态修复中具有重要作用。
3.2 暖季型草坪草对土壤pH值和电导率的影响
比较2种暖季型草坪草种植样地和对照样地土壤的pH值和电导率发现, ‘阳江’和‘苏植3号’种植样地土壤的pH值和电导率均显著低于对照样地,对照样地土壤电导率为草坪草种植样地的6倍以上。土壤pH值降低可能是由‘阳江’和‘苏植3号’根系分泌物所致[35],也可能是因为草坪草对土壤的覆盖抑制了碱性阳离子上移,具体原因还有待进一步深入研究。‘阳江’和‘苏植3号’均具有双细胞的盐腺,为泌盐植物,在盐碱地种植后,这2种草均会通过根系从土壤中吸收大量盐离子[36],同时‘阳江’和‘苏植3号’具有超强的地面覆盖能力,成坪后会显著抑制盐碱地土壤返盐[37],而土壤电导率与土壤盐度存在正相关关系[38-39],土壤电导率的降低也伴随着土壤盐度的降低。因此,在盐碱地种植‘阳江’和‘苏植3号’后既降低了土壤pH值,又降低了土壤盐度,盐碱地土壤得到改良。
3.3 暖季型草坪草对土壤微生物群落的影响
植物根系与土壤细菌互作会影响土壤细菌多样性[40];植被类型不同还会影响土壤固氮细菌群落组成及丰富度[41]。相关研究结果[42-43]显示:植物的生物量和养分含量等与根际细菌群落多样性和组成呈显著相关。本研究中,‘阳江’和‘苏植3号’种植样地土壤中细菌群落结构与对照样地差异显著,其中,细菌Shannon-Wiener多样性指数和固氮细菌相对丰富度均显著高于对照样地;且细菌Shannon-Wiener多样性指数和固氮细菌相对丰富度与植株地下部生物量呈显著或极显著正相关。因此,推测造成以上差异的原因与植物物种和生物量不同有关。此外,有研究发现土壤的pH值和电导率也会影响土壤细菌群落[44-45];土壤电导率与土壤盐度紧密相关,而土壤盐度与细菌多样性呈负相关[46-47]。相关性分析结果显示:细菌Shannon-Wiener多样性指数和固氮细菌相对丰富度与土壤pH值分别呈极显著和显著负相关,与土壤电导率也分别呈极显著和显著负相关。说明细菌多样性和固氮细菌丰富度的增加还可能是因为土壤的pH值和电导率降低。土壤细菌多样性的增加有利于调节植物生长发育、抑制病原菌和加速土壤中营养元素矿化,在加速土壤中能量转化和物质循环方面有重要的作用[48]。而土壤中固氮细菌不仅可以固定空气中的氮气,还可以产生植物激素促进植物生长[49]。因此,种植‘阳江’和‘苏植3号’可以改变样地土壤理化性质,进而增加土壤细菌多样性和固氮细菌丰富度,有利于增加土壤肥力和调节植物生长发育。
‘阳江’和‘苏植3号’种植样地土壤中真菌Shannon-Wiener多样性指数与对照样地差异不显著,但丛枝菌根真菌和粪腐生真菌的相对丰富度显著高于对照样地,且丛枝菌根真菌相对丰富度与植株地下部生物量呈显著正相关,粪腐生真菌相对丰富度与土壤pH值呈显著负相关。这可能是因为丛枝菌根真菌与植物互利共生,丛枝菌根真菌从植物体中获得糖类等养分供自身生长,也会帮助植物吸收土壤中的氮、磷等矿质营养[50-51]。同时也反映出丛枝菌根真菌丰富度的增加可能与植株地下部生物量增加有关,粪腐生真菌丰富度的增加可能与土壤pH值的降低有关。
种植其他耐盐植物也可改善盐碱地土壤理化性质或增加微生物多样性。例如:种植苜蓿(MedicagosativaLinn.)、向日葵(HelianthusannuusLinn.)、玉米(ZeamaysLinn.)和多年生黑麦草可以降低盐碱地土壤的pH值和盐度,增加土壤有机质含量,其中种植玉米还可以增加土壤真菌多样性[52];种植费菜(PhedimusaizoonLinn.)和蒲公英(TaraxacummongolicumHand.-Mazz.)可以提高盐碱地土壤中细菌多样性[53]。与这些耐盐植物相比,暖季型草坪草的种植还可美化环境。江苏海岸线长,滨海盐碱地面积大,有着巨大的绿化需求,种植暖季型草坪草对该地区的滨海盐碱地具有重要意义。
综上所述,在江苏滨海盐碱地中种植‘阳江’和‘苏植3号’后显著增加了土壤有机质含量,降低了土壤的pH、电导率和盐度,增加了细菌多样性和固氮细菌丰富度以及丛枝菌根真菌和粪腐生真菌的丰富度。因此,在江苏滨海盐碱地中种植这2个品种有利于增加土壤肥力和土壤细菌多样性,提高重要功能细菌和真菌的丰富度,且有利于美化环境,在盐碱地生态修复中具有重要作用。
猜你喜欢
军事文摘(2024年6期)2024-02-29
五金科技(2022年1期)2022-03-02
广东开放大学学报(2021年5期)2021-11-19
中国核电(2021年3期)2021-08-13
河北画报(2020年21期)2020-12-14
五金科技(2020年4期)2020-09-23
河北果树(2020年1期)2020-02-09
小学生必读(低年级版)(2019年5期)2019-08-30
家教世界(2017年8期)2017-06-21
孩子(2016年4期)2016-04-13
