
植物酸奶品质优化的研究进展
2024-01-30 02:12:30尹新雅
食品工业科技 2024年3期
尹新雅,王 爽,朱 玲,张 晖
(江南大学食品学院,江苏无锡 214122)
根据团体标准T/WSJD 12-2020《植物蛋白饮料植物酸奶》的定义,植物酸奶是“以含有一定蛋白质的植物和(或)其制品为原料,经杀菌、发酵等pH 降低,发酵前或发酵后添加或不添加非动物来源配料加工制成的植物蛋白饮料产品”[1]。植物酸奶的消费量近年来持续上升,因其不含胆固醇,并且满足牛乳过敏人群、环保人士及关注动物福利人群的需求,同时有利于缓解畜牧业压力,符合当前绿色健康食品发展的潮流。但因起步较晚、相关生产工艺不成熟,导致其在营养、质构等方面存在诸多问题[2],严重影响其品质。
植物酸奶的品质特性与植物蛋白的功能特性(即乳化性与溶解性等)、乳酸菌产酸等息息相关,并且受到配方、发酵前加工及发酵方式的影响。目前,植物酸奶品质优化技术基本处于探索阶段,主要包括改良配方(如添加辅料、功能因子或稳定剂)[3-5]、优化加工技术(如采用热处理、酶解等技术)[6-7]及优化发酵方式(单菌、复合菌或直投式发酵剂进行发酵)[8-10],且不同调控方法对植物酸奶品质的作用效果存在差异。本文从不同植物酸奶品质的主要特性及其调控技术等方面进行阐述,并对未来的研究方向进行展望,以期为植物酸奶品质优化的研究开拓切实有效的调控技术提供科学指引。
1 不同植物酸奶品质的主要问题
植物酸奶可选用的原料包括豆类、坚果类、谷类、其他原料(椰浆、藜麦),以及它们的制品[1]。不同原料来源的植物酸奶具有各自的优势,但是也存在不同的品质问题。
1.1 豆类酸奶
豆类酸奶的主要原料是大豆、豌豆等[1]。大豆中胆固醇含量较低,蛋白含量高,是植物乳及其制品的良好原料来源。然而,大豆酸奶在风味、质构等方面还存在影响其品质的问题,比如豆腥味重、质地不够细腻。另外,豆乳中不含乳糖,葡萄糖含量少,只含有一定的蔗糖、棉子糖和水苏糖,导致传统的酸奶发酵剂不能很好地在其中生长。因为保加利亚乳杆菌缺乏相应的酶系,分解蔗糖、水苏糖等二糖的能力较弱,进而引发产酸减少,而嗜热链球菌可以分解蔗糖,但是产酸能力弱[11]。因而大豆酸奶的酸度与传统酸奶有差距。
豌豆的氨基酸组成平衡,豌豆蛋白的致敏性较低且消化率较高,胰蛋白酶制剂等抗营养因子低于大豆,制备酸奶后蛋白含量几乎可以与传统酸奶持平,逐渐进入人们的视野。然而,豌豆酸奶存在2-甲氧基-3-异丙基-(5 或6)-甲基吡嗪为主的异味物质,因此携带“青草味”等不愉快的风味[12]。同时,豌豆蛋白的溶解性较低,乳化性较差,因而引发豌豆乳出现严重的分层现象。这种现象会导致蛋白质在酸奶发酵过程中沉淀,乳清大量析出,无法成功发酵出均一稳定的酸奶体系。即使可以形成豌豆酸奶,其也会因凝胶强度弱、析水率高或颗粒感明显而影响消费者的喜爱程度[13]。
1.2 坚果酸奶
坚果酸奶的原料来源主要有榛子、核桃、腰果、杏仁等[1]。坚果中的不饱和脂肪酸较多,对肥胖、高胆固醇等人群有益。然而,坚果酸奶在营养、质构及风味上存在一定问题。比如,腰果中的三种储存蛋白(Ana o1、Ana o2 和Ana o3)会引起严重的过敏反应,研究表明,发酵可以破坏Ana o1 和Ana o2 蛋白,但是Ana o3 蛋白的稳定性较高,在腰果酸奶中依然存在[14],所以腰果酸奶不适宜对其过敏的人群食用。与传统酸奶相比,杏仁酸奶中的蛋白质含量普遍较低[15],这和杏仁蛋白的弱疏水性和低分子量有关,进而导致其在杏仁乳中的溶解性和乳化性差,形成了不稳定的酸奶体系。另外,杏仁酸奶处理不当会呈现涩味,降低酸奶的品质。榛子乳也逐渐被用于发酵酸奶,但其蛋白质溶解度较低,放置易沉淀,同时,体系中油滴尺寸较大,易上浮于液体表面,形成“脂肪膜”,体系稳定性差,导致乳体系分层[16],而发酵会促使分层更为明显,呈现显著的蛋白沉淀、脂肪漂浮和乳清析出等现象。所发酵的固体酸奶外观上缺乏典型的凝胶特征,搅拌型酸奶因粘度低、析水多而缺乏醇厚的口感。此外,核桃不断被用于植物基产品开发中,但是据报道,核桃乳因较多的油脂和醛类物质易呈现出哈味或生草味,进而给核桃酸奶风味带来了负面影响[17]。并且,核桃蛋白中的谷蛋白含量较多,因而导致其溶解性较差,限制了其酸奶产品的生产。
1.3 谷类酸奶
燕麦、小米等是谷类酸奶的原料主要来源[1]。燕麦富含可溶性膳食纤维和β-葡聚糖,对人体健康有积极影响,然而,燕麦酸奶在质构、风味上仍有待改善。燕麦中的球蛋白在中性或微酸性条件中溶解性较差,这主要是因为燕麦球蛋白二级结构中,β-折叠会向无规卷曲转变,引发不溶性聚集体形成[18],导致燕麦乳体系不均匀,使燕麦酸奶品质受损。此外,燕麦中脂质含量较多,容易氧化产生己醛、戊醛和一些羰基化合物,给燕麦酸奶带来不愉快的气味,且长链脂肪酸会导致酸奶产生苦味[19]。小米酸奶方面,营养不足、稳定性差是其面临的主要问题。因为小米乳中蛋白含量较低,而蛋白含量不仅关系到酸奶的营养,还和凝固型酸奶的形成有关[20]。并且,在不添加稳定剂或增稠剂的情况下,小米乳难以发酵出质地良好的小米酸奶[21]。
1.4 其他
除了以上三种类型植物酸奶之外,藜麦和椰子也可作为原料发酵植物酸奶[1]。藜麦营养丰富,富含蛋白质、低升糖指数的碳水化合物、植物甾醇等物质。然而,尽管藜麦蛋白含量高于燕麦、小米等谷类原料,和豆类中的蛋白含量相似[22],但可溶性蛋白含量不足,有研究指出,纯藜麦酸奶的酸度只有30 °T,而纯大豆酸奶的酸度却能达到70 °T,即使它们最终的pH 都是4.0,这可能和大豆中可溶性蛋白含量更多有关,其羧基和氨基分别提供和接受质子,大大增强了大豆蛋白的缓冲能力,即需要更多的产酸量才能达到相同的pH[22-23]。对于椰子来说,其脂肪(31%~35%)和蛋白质(3.5%~4.0%)含量较高,且富含必需氨基酸、维生素等营养物质[24]。另外,大豆及坚果中存在植物性致敏源已有报道,但椰子引发的过敏现象十分罕见。然而,椰子蛋白的乳化性较弱,这导致椰奶表现为一种相对不稳定的水包油乳液[24],故椰子酸奶的质构有待改善。
综上,植物酸奶在品质上存在亟待改善之处。目前,在营养方面,不同植物酸奶的蛋白质含量不一,功能性有待提高,且豆类、坚果类酸奶中可能存在抗营养因子或致敏原,影响人体对营养物质的吸收;质构方面,一般未经处理的植物乳发酵出的酸奶,存在乳清析出、质地粗糙以及分层等诸多缺陷,这和植物蛋白、淀粉等大分子会随时间的延长而不断沉淀聚集、脂肪等比重轻的分子易上浮有关;另外,谷类、豆类及坚果类酸奶若处理不当,容易携带不良风味,因此有必要从改良风味的角度找出针对性的调控手段来掩盖、消减异味;发酵特性方面,植物乳作为发酵基质时,因为可溶性碳水化物浓度通常较低、多数植物蛋白的溶解度较弱,导致乳酸菌的生长速度较慢,酸化程度非常有限。不同植物酸奶的优点及品质缺陷见表1。

表1 不同植物酸奶的主要优势及品质问题Table 1 Main advantages and quality defects of different plant-based yogurt
2 植物酸奶品质的常见调控措施
目前,关于植物酸奶品质调控技术的研究相对较少,主要集中于三个方向:一是改良植物酸奶的配方;二是利用加工技术改变植物蛋白的结构(去折叠、聚合、交联等)进而提高蛋白质的溶解性和乳化性,以及减小油滴的尺寸,使其均匀分布在乳体系中以达到提高稳定性和细腻度的目的;三是改变发酵方式来增加植物蛋白的网络交联或是产生特殊的芳香物质,以达到改良质构和风味的作用(表2 和图1)。
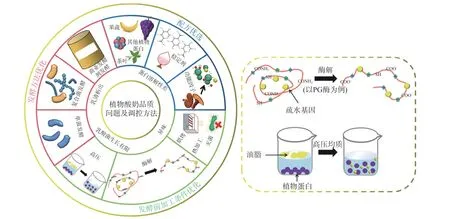
图1 植物酸奶品质的主要缺陷及调控措施Fig.1 Main defects and regulations of plant-based yogurt quality

表2 植物酸奶品质的调控方法、效果及优缺点Table 2 Methods,effects,advantages and disadvantages of regulations for plant-based yogurt quality
2.1 配方优化
2.1.1 添加辅料 单一原料的植物酸奶可能在营养功能、风味等方面存在不足,所以通过添加辅料的方式不仅可以强化产品营养,对于改善风味和质构也具有重要作用(表2)。目前主要有三种途径,一是添加酸甜可口、富含维生素与矿物质的果蔬汁,或营养价值高的药食同源物质,如陈皮、山楂、枸杞、沙棘等,一般将其以汁液或浸提液的形式与主料混合发酵。二是在原有植物原料基础上,采取不同来源的植物原料混合发酵,如此不仅能优化酸奶的质构,还能与酸奶营养互补。可选蛋白和淀粉含量较高的植物原料作为辅料,且该辅料不可携带异味。藜麦在此方面的作用效果较为明显[13,23],其中有较多B 族维生素、生育酚、叶酸,可以发挥对人体有益的功能性作用[3],将其制成浆后发酵而成的酸奶风味较为和谐。同时,藜麦中淀粉、蛋白占比较高,可增加乳体系中蛋白含量;受热膨胀后淀粉可发挥“填充效应”,从而加强蛋白质网络对水分子的束缚,提高硬度,减少析水,因而可以弥补其他原料植物酸奶在质构方面的不足。研究表明,纯大豆酸奶的持水力最低(43.3%),而将藜麦浆和大豆浆混合发酵(比例为4:6 时,持水力可以增加到57.6%),很大程度减少了纯大豆酸奶质构上的不良性质[23],且酸奶的持水力随着藜麦比例的增加而增加,同时产品在储存期间稳定性也保持良好。可见加入藜麦之后,体系中蛋白-蛋白交联增多,淀粉含量提高,蛋白质网络结构对水的束缚作用明显增强。三是加入具有特殊香味的辅料,例如茶叶,抹茶粉就是新鲜茶树叶经过杀青、干燥、碾磨后得到的具有芬芳茶香味的产品,可以起到良好的掩蔽作用,将其与纳豆豆浆共同发酵,掩盖了纳豆激酶的“氨臭味”[10],使纳豆酸奶的风味得以改善。
2.1.2 添加功能因子 随着人们对营养健康的需求不断增多,含有功能因子的食品愈加受到青睐。食品功能因子种类繁多,成分与功能各不相同。目前,多糖、黄酮、植物多酚等常被用作功能因子添加在牛乳酸奶中,主要目的在于改善营养功能特性及质构。然而,将其用于植物酸奶的报道较少。因此,可参考功能因子对牛乳酸奶品质优化的效果,选择适当的物质优化植物酸奶的品质(表2)。真菌多糖在酸奶抗氧化作用方面具有很好的应用,研究指出,牛乳酸奶添加茶树菇多糖后,在一定浓度范围内,抗氧化能力与多糖的量有正相关性。在高剂量给药组小鼠血清中,检测到超氧化物歧化酶(SOD)活力提高了55.87%,谷胱甘肽过氧化物酶和过氧化氢酶活力分别提高了60.86%和76.51%[25],这可能一方面是由于真菌多糖可提高酸奶或人体内的抗氧化酶活性,使衰老引起的氧化损伤得到修复或抑制,另一方面和真菌多糖的结构中存在具有抗氧化性的醇羟基有关,能够引发脂质过氧化的延缓,进而抑制活性氧的产生。此外,其他植物提取物如多酚、黄酮类物质的抗氧化功效优良。郭俊花等[4]研制的添加槐米提取物的牛乳酸奶中,发酵第1 dDPPH 自由基的清除率达到了60.3%(50 g/mL),且在储存过程中清除率有上升趋势,原因是槐米中含有黄酮类物质,不仅可以直接对羟自由基、超氧阴离子等清除和阻断,还可以提高体内抗氧化系统的作用,抑制自由基产生,从而起到延缓衰老的作用。尽管多糖、黄酮等功能因子在酸奶中的应用前景广阔,然而,多酚、黄酮等功能因子自身易于被氧气等外部条件破坏,稳定性较差,故其作用于植物酸奶的抗氧化功效在储存期内是否能够维持,有待深入研究。
此外,真菌多糖还可以改善酸奶的质构,这主要体现在酸奶持水力得到提高,但高浓度多糖会降低酸奶持水力。张岩等[32]评价了松茸多糖对传统酸奶品质的影响,结果表明松茸多糖对乳酸菌的促生长、酸奶持水力、硬度的影响呈现倒“U”型,可见这些积极作用并不表现出对多糖浓度的依赖性。原因是一定浓度松茸多糖可以引发蛋白质聚集进而增强分子间相互作用,还可以抑制乳酸菌在储存过程中产酸,从而消减酸奶后酸化现象,减少其析水率,达到优化酸奶组织状态的目的。但是过多的松茸多糖反而降低酸奶持水力,因为其会阻碍蛋白形成致密、牢固的凝胶网状结构。
2.1.3 添加稳定剂 稳定剂是饮料、酸奶中的常见成分,主要有胶类、藻酸类衍生物、盐类等,稳定剂可通过加强蛋白质分子之间的交联或者填充在蛋白质网络的孔隙中,形成了更致密的三维网络,从而提高酸奶体系的持水力、硬度、粘度[5](表2)。稳定剂可以分为单一型和复配型,Kong 等[33]用低酰基结冷胶(LAG)促进了大豆酸奶凝胶形成,同时提高了其持水力。当LAG 的量从0.025%增加到0.1%时,大豆酸奶的持水力从90.75%上升到了96.13%,可见胶类物质的存在有助于大豆酸奶凝胶的稳定性,这可能与多糖和蛋白质分子之间交联形成了孔隙更小、结构更密的网络,使凝胶对水的拦截和保留能力增强有关;此外,多糖中含有较多的亲水基团,可以增加氢键的数量来结合更多水分子。类似的结论在复配稳定剂改善酸奶品质的研究中也被提出,据胡颖等[5]报道,黄原胶与CMC 按1:2 复配后,改善了苏籽酸奶质地,原因也是蛋白分子之间的共价交联被多糖增加,交联程度的增加效应使得更多水分子得以保留,因而提高了酸奶的机械性能。然而,传统稳定剂无法满足清洁标签的要求以及人们对健康无添加食品的需要。因此,对于能够替代传统稳定剂的其他物质的探寻和研究仍需深入开展。
2.2 加工技术优化
2.2.1 热加工 热加工技术因成本低廉、容易操作,是目前植物酸奶品质优化最常用的方法之一(表2)。热加工主要的对象是植物乳和/或原料,对植物乳进行热处理不仅能够起到灭菌作用,产生独特风味物质,还能引起蛋白质去折叠、聚合及二硫键形成等改善植物酸奶的风味和质构,且改善程度取决于温度及持续时间。但是不当的热处理会使植物蛋白在加热过程中变性,进而引发植物乳体系变得松散、出现絮状沉淀,无法进一步发酵。植物乳在发酵前需经过灭菌,目前关于植物酸奶的灭菌条件尚无定论。在不加稳定剂的条件下,笔者团队前期考察了热处理(121 ℃,15 min;95、90、85 ℃分别持续5、10、15、30 min;65 ℃,30 min)对纯核桃酸奶的灭菌效果,发现85~121 ℃处理的核桃酸奶均表现出不同程度的变性,体系不仅出现了絮状物,同时95 ℃的高温使得核桃蛋白形成了热凝胶,且121 ℃处理的样品还发生了褐变;而通过巴氏杀菌(65 ℃,30 min)处理的样品不仅能够顺利发酵,而且在微生物检测中也未出现杂菌污染,同时乳酸菌的活菌数也达到团体标准的要求。也有报道指出[34],用90~100 ℃对巴旦木乳杀菌 5~10 min,在充分杀菌的同时也不会使巴旦木蛋白变性,这可能与不同植物蛋白的结构与性质有关,因而热稳定程度不同;又或者是与有无稳定剂的存在有关。
除了热加工的条件之外,热加工方式也是植物酸奶品质的重要影响因素。例如灭菌的方式不同,最终产品的品质会有区别,Hickisch 等[26]分别用巴氏杀菌(80 ℃,60 s)和超高温加热(UHT,140 ℃,10 s)对羽扇豆蛋白进行热处理,发现UHT 对羽扇豆酸奶质地的改善更佳,这可能是由于该过程不会导致羽扇豆蛋白完全变性,而且还能引发蛋白发生更多的交联,形成了更为致密的网络结构。此外,烘烤、漂烫等也是针对原料常用的热加工技术。烘烤最大的优势是可以促进植物原料产生独特的香味,红豆在140 ℃下烘烤30 min 制成的酸奶豆香浓郁,这和红豆当中蛋白与糖类发生了美拉德反应有关,但是需控制烘烤的时间和温度,因为过度的烘烤会使原料通过美拉德反应产生丙烯酰胺、呋喃等污染物[30]。可见,不同来源的植物酸奶中所含的蛋白在组成和结构上不尽相同,其所能承受的热加工方式、程度等也不尽相同,因此热处理对植物酸奶风味、质构等特性的影响还需深入探究。
2.2.2 酶处理 酶解技术可以显著增加植物蛋白的溶解性、乳化性等功能特性。植物酸奶的酶改性已有研究,如转谷氨酰胺酶(TGase)可以促进赖氨酸的ε-氨基与谷氨酸的γ-羟酰胺基发生分子内交联,形成分子量更大的聚合物,加强蛋白质网络持水力,提高凝固型酸奶硬度(表2)。研究报道,TG 酶与乳酸菌联合发酵酸奶,与不添加TG 酶的样品相比,大豆-牛乳双蛋白体系形成的凝胶结构更加致密,网络孔隙更小[27]。这是因为TG 酶诱导蛋白交联成更大的聚合物,分子量增加,导致蛋白质胶粒网状结构增强,更有利于对水分的束缚,使其不易析出。而Nie 等[13]研究显示,仅用TG 酶处理后,绿豆蛋白酸奶的持水力并没有提高,这是因为TG 酶一般不使蛋白质去折叠,并且引起的蛋白网络交联是不均匀的,因而对水分的束缚作用较小。可见TG 酶对植物酸奶质构的作用并未形成统一定论。另外,TG 酶在动物酸奶中的研究更为丰富,将其用于纯植物基酸奶质构改善的报道较少,这可能是因为动物蛋白和植物蛋白结构的不同,所以该酶对纯植物基酸奶质构的影响还需更多阐明。
蛋白质谷氨酰胺酶(PG)处理在提高植物蛋白的溶解性和乳化性方面也具有广阔的应用前景,其一方面可以使蛋白质去折叠,引发疏水基团的暴露,从而更好地在界面展开;另一方面可将谷氨酰胺的酰胺基转变成羧基,减少分子内或分子间的氢键,增强静电排斥[28],增加植物蛋白可溶性(表2)。目前关于PG 酶对植物酸奶品质优化的研究较少,笔者团队研究发现TG 酶对核桃蛋白的改性作用并不大,核桃蛋白依旧会在热处理和高压均质中变性,而PG 酶处理后的核桃蛋白溶解性得到提高,乳化性明显增强,在不添加增稠剂的条件下依然能得到均匀稳定的发酵乳体系,且酸奶凝胶的硬度也得到增强。原因是PG 酶诱导的脱酰胺引发蛋白质构象变化,暴露出隐藏在内部的疏水位点,同时分子间静电排斥增强,从而引起溶解性和稳定性的增加。尽管PG 酶能够改善植物蛋白的功能性,但是其成本较高。所以是否还有其他酶,不仅价格低廉,而且可提高植物蛋白溶解性进而得到稳定酸奶体系,这一点仍需深入研究。
2.2.3 高压处理 高压处理技术是使植物乳体系均一化的重要调控手段,可减小植物乳中脂肪滴的尺寸,使其均匀地分散在乳体系当中。同时,植物蛋白会在高压、空化、剪切、湍流中去折叠,进而提高乳化性,消减脂肪漂浮和蛋白质沉降导致的失稳现象(表2)。目前,高压处理条件尚无定论,常见的条件一般分为两类:一是20~40 MPa,循环2 次(有的研究会采用第一次5 min,第二次10 min 的模式[35]);二是超高压均质,压强在100~400 MPa 范围内,循环1~2 次。研究表明[36],马铃薯蛋白乳发酵前经过200 MPa 的高压处理后,体系中脂肪及蛋白质颗粒的尺寸会明显减小,200 MPa 处理的马铃薯酸奶的持水力比压力为120 MPa 处理时提高了7%,白度和细腻度都增加,且发酵产品的乳清析出有所改善,这主要是因为大的脂肪滴会破坏蛋白质网络的均匀性,而高压处理使其变成更小的液滴填充在蛋白质基质中,形成对水具有较高截留能力的精细网络,且高压处理一般影响蛋白质的非共价键,不引起共价键的破坏,进而提高其功能性。可见,高压处理是提高植物乳的稳定性的有效途径,且在一定范围内,酸奶的质构特性随着压力的增加而提升。但是最终发酵产品的质构特性与所用的压力与均质时间之间的关系还有待深入阐明。
2.3 发酵方式优化
目前采用的发酵方式包括:单菌发酵、复配菌发酵及直投式发酵剂发酵。然而,不同发酵剂对酸奶质构、风味等存在不同影响,这就需要在植物酸奶的加工过程中选择适宜的菌种进行发酵(表2)。
2.3.1 单菌发酵 单菌发酵有一定优势,如植物乳杆菌是酸奶发酵的常用菌种,其单独发酵时可以赋予酸奶独特的香味,对产品风味特性的改善具有相应的促进作用。另外,一些乳酸菌具备胞外多糖分泌能力,可改善酸奶质构,优化酸奶组织状态。据报道,魏斯氏菌MG1 制备的藜麦酸奶具有良好的持水力(>90%),这和该菌产生的胞外多糖加强蛋白质网络的交联度,提高其缚水能力有关[22]。然而,酸奶质构和胞外多糖的产生量之间呈何种相关性,如何促进菌株产生胞外多糖等问题还需进一步探讨。
2.3.2 复配菌发酵 因为单菌发酵具有局限性,比如,利用罗伊氏乳杆菌单一发酵凝固型酸奶时,发现其凝乳效果较差[8],所以复配菌发酵成为热门的研究方向。在复配菌组合中,传统酸奶一般采用保加利亚乳杆菌和嗜热链球菌发酵,但它们在植物乳中的作用有限。如,植物乳中缺乏乳糖,通常需添加外源糖(以蔗糖为主),而保加利亚乳杆菌对蔗糖等二糖的水解能力不强,故产酸减少;另外,虽然嗜热链球菌可以分解蔗糖,但其产酸能力弱[11];且这两种菌耐胆盐、粘附定殖效果较差[37],因此不同菌种复配发酵越来越广泛。朱晓丽等[8]用罗伊氏乳杆菌同嗜热链球菌和保加利亚乳杆菌进行复合发酵薏米酸奶,发现3 菌株协同发酵比其中任2 株菌组合发酵的样品质构特性更好、风味物质的种类更多,苦味、涩味大大降低,且具有降血脂功效。然而,并不是菌种越多,发酵的酸奶品质越好。据报道[31],嗜酸乳杆菌、鼠李糖乳杆菌等8 种乳酸菌经不同组合复配发酵的大豆酸奶中,8 菌发酵样品的质构特性并不优于较少数量菌种复配发酵的酸奶,这可能是在数量较多的情况下,菌株之间存在对营养物质的竞争作用以及弱势菌的生长受到抑制。
2.3.3 直投式发酵剂发酵 无论是采用单一还是复配菌株,一般需要经过培养、驯化等较为繁琐的步骤。为了提高植物酸奶的生产效率,直投式发酵剂带来了更多便利,实际应用效果也较为可观。研究表明,在9 种商品发酵剂中,7 菌型酸奶发酵剂对核桃粕酸奶的质构具有最优的改善效果,其中持水力和硬度分别达65%和2.082 N[9],且风味较为协调,这和该7 菌型发酵剂能够适应核桃粕酸奶的环境有关。可见,直投式发酵剂可用于植物酸奶的发酵,但其发酵效果会根据原料的种类而有区别,需有针对性地进行筛选。
综上,不同的优化措施均可一定程度改善植物酸奶的品质,但也有各自的缺陷,多种调控方法联用在实际应用中更为常见。如,Nie 等[13]利用椰子油、pH 调节与TG 酶处理联合制备了凝固性较好的绿豆蛋白酸奶,其乳清析出较少、网络结构较致密。另外,汤木果等[38]将均质和热处理配合使用,提高了澳洲坚果酸奶的硬度并改善了其风味。可见,多种方式联合处理比使用单一方法更能有效地改善植物酸奶的品质。本文对多种调控措施联合改善植物酸奶品质的例子进行了总结,详见表3。

表3 多种调控方式联合使用对植物酸奶品质的改善效果Table 3 Effects of combination of different regulatory methods on the improvement the qualities of plant-based yogurt
3 小结与展望
植物酸奶正成为食品行业中市场份额快速增长的创新型产品,它在可持续性、环境保护等方面比传统酸奶更具优势。植物酸奶的品质是影响其商业价值、消费者喜爱度的关键因素。由于原料不同,植物酸奶存在不同的品质问题。大豆酸奶易携带不良风味;坚果酸奶往往因蛋白溶解性差,存在分层现象;谷类酸奶,如燕麦酸奶易发生脂肪氧化,小米酸奶存在蛋白含量低的问题。植物酸奶的品质特性与蛋白溶解性、乳酸菌活性等关系密切,并且受到配方、加工条件及发酵方式的影响。植物酸奶品质调控技术如添加辅料、热处理、高压、酶解和发酵技术,都不同程度改善了植物酸奶的不良品质特性且效果不同。添加辅料可提高植物酸奶的营养;酶解、高压等加工技术可提高蛋白溶解度、使油滴均匀分布或加强蛋白网络交联,进而稳定植物乳体系、促进胶凝、减少析水;不同的发酵方式会产生不同的芳香物质或营养物质。同时,这些方法也具有各自的局限性,因此可通过多种调控技术联合处理来弥补单一方法对植物酸奶品质优化作用的不足。目前,植物酸奶品质的影响因素及其作用机理的阐述仍然较浅显,植物酸奶品质优化的方式相较于传统酸奶仍然较少。未来应开展更多关于天然稳定剂、原料制浆方式、益生菌及生物活性物质的递送体系作用于植物酸奶品质的研究,并结合发酵动力学、代谢组学、微观结构观察等实验,进一步明确植物酸奶质构、风味改善的主要机理。同时通过体外消化或动物、细胞模型开展营养评价研究,以期更加有效地提升植物酸奶的品质。
猜你喜欢
中老年保健(2022年3期)2022-08-24 02:59:30
食品安全导刊(2021年21期)2021-08-30 08:22:00
中国生殖健康(2020年8期)2021-01-18 03:05:44
新世纪智能(高一语文)(2020年10期)2021-01-04 00:43:56
新世纪智能(高一语文)(2020年10期)2020-12-31 09:26:20
少儿美术(快乐历史地理)(2020年7期)2020-11-26 06:25:50
家庭影院技术(2019年1期)2019-01-21 02:25:02
中国生殖健康(2018年3期)2018-11-06 07:20:20
启蒙(3-7岁)(2018年7期)2018-07-10 09:38:16
食品工业科技(2014年11期)2014-03-11 18:16:08
