
Si-SETDB1通过SPG20甲基化对肺腺癌细胞迁移侵袭的影响
2024-01-30 05:27赵宝山孙光蕊郑竞雄梁宗英
河北医学 2024年1期
赵宝山, 杨 阳, 孙光蕊, 郑竞雄, 梁宗英
(1.承德医学院附属医院胸外科, 河北 承 德 067000 2.河北省胸科医院临床药理试验部, 河北 石家庄 050000)
肺腺癌是肺癌中最多见的病理分型,且发病率逐年增多[1]。传统肺腺癌治疗以手术和术后放化疗为主,随着医学的发展,现代精准医学的分子靶向治疗和免疫治疗作为治疗肺腺癌的新方法,已取得巨大进展[2]。痉挛性截瘫-20(SPG20)是近年来发现的最新的与多种癌症相关的基因,其甲基化修饰与多种实体肿瘤的增殖、侵袭和转移密切相关[3]。SET结构域分支型1(SET domain bifurcated1,SETDBl)是组蛋白甲基化转移酶家族中重要一员,其功能可以催化多种基因发生甲基化抑制基因的表达,与多种肿瘤密切相关[4]。国内外针对SETDB1和SPG20在肺腺癌发病中的相关性研究鲜有报道,鉴于此,本文旨在通过检测肺腺癌组织细胞中SETDB1表达和SPG20甲基化水平,探究下调SETDB1对SPG20甲基化及A549细胞迁移和侵袭的影响。
1 资料与方法
1.1一般资料:2020年1月至2021年1月在承德医学院附属医院胸外科手术切除原发肺腺癌患者60例,其中男性16例,女性44例;≥60岁22例,<60岁38例;Ⅰ期26例,Ⅱ期16例,Ⅲ期18例;低分化8例、中分化38例,高分化14例;淋巴结转移16例,无淋巴结转移44例。取患者癌组织及同一患者正常肺组织作为研究对象。
1.2主要试剂:甲基化试剂盒购于上海晶抗生物工程有限公司;焦磷酸测序试剂盒购于北京阅微基因技术股份有限公司;DNA提取试剂盒购于广州皓宝生物科技有限公司;免疫组化试剂盒购于广州露生科技有限公司;SETDB1、SPG20单克隆抗体购CST公司。A549细胞系购于广州皓宝生物科技有限公司。
1.3方 法
1.3.1qRT-qPCR方法:提取RNA,分光光度计定量;应用Prime ScriptTM RT Reagent kit反转录;应用SYBR Green qPCR Master荧光定量试剂盒检测mRNA的表达;相对表达量以2-ΔΔCt计算。
1.3.2免疫组化(SP法):组织切片二甲苯及梯度酒精脱蜡;3%H2O2浸泡去内源性的过氧化氢酶,然后倒掉H2O2,在清水中洗两次,加入柠檬酸缓冲液修复抗原;载玻片冷却至室温后PBS洗2次,加入血清非免疫山羊血清封闭,加入一抗孵育;PBS处理后,加入二抗孵育;DAB显色,苏木精染色;中性树胶封片固定。
1.3.3免疫印迹实验:提取蛋白,制备电泳蛋白上样液,进行凝胶电泳。电泳后90V 60~80min转膜。重复洗膜。孵育一抗(SPG20抗体1∶2000;SETDB1抗体1∶2000;GAPDH抗体1∶1000),4℃过夜。次日孵育二抗,重复洗膜。暗室发光显像。
1.3.4焦磷酸测序法:DNA提取试剂盒提取总DNA,采用ED DNA Methylation-Gold kit进行亚硫酸盐转化。SPG20甲基化扩增正向引物序列为5'-AGGAAGTATGAAAGAATGTATTGTAAAG-3',反向引物为5'-CCCCTCAAAATTAAACAACCTTTCTCTACA-3'。产物进行焦磷酸测序,PyroQ-CpG软件分析甲基化状态。
1.3.5细胞培养、转染及分组:实验组:SETDB1 siRNA组(si-S)、阴性对照组:SETDB1 阴性siRNA组(si-N)、空白对照组:未处理A549细胞组(NC)。应用10%胎牛血清的1640培养液在37℃ 5%CO2培养箱中培养。稀释SETDB1 siRNA或阴性对照siRNA于Lipofectamine 3000溶液中,按试剂盒说明书操作,采用siRNA/lipofectamine混合溶液各自培养于含10%胎牛血清的DMEM液中转染。
1.3.6MTT实验:取对数生长期的细胞悬液,计数后将各组细胞以1×103个/孔接种于96孔板。细胞转染后,2h、12h、24h、36h、48h观察细胞状态,每个时间点拿出一块板,每孔加入噻唑蓝溶液30μL,室温孵育4h后,加入150μL DMSO,80r/min摇匀。酶标仪测定570nm处OD值。
1.3.7划痕修复实验:取对数生长期的细胞悬液,计数后接种于24孔板。细胞培养至铺满孔底。无菌移液枪头划痕,PBS缓冲液洗去脱落细胞并拍照; 常规培养,48h后再次拍照并计算迁移距离。细胞迁移距离=0h初始划痕宽度-48h后划痕宽度。
1.3.8侵袭实验:应用基底膜基质胶将上室包被24h,于上室接种各组细胞混悬液,下室加DMEM培养基。孵育箱内常规培养48h后弃培养基,棉签擦去上室未穿膜细胞,4%多聚甲醛固定下层细胞,应用结晶紫染色,倒置显微镜下计数穿膜细胞。
1.4统计分析:SPSS21.0软件统计分析。计数资料用率表示,χ2检验。计量资料用均数±标准差表示,均数比较采用t检验及单因素方差分析,相关性分析采用Pearson法,以P<0.05为差异有统计学意义。
2 结 果
2.1组织中SETDB1和SPG20 mRNA表达量:qRT-PCR结果显示,SETDB1 mRNA相对表达量为(1.62±0.11),高于正常肺组织的(0.30± 0.04)(t=3.232,P<0.05);SPG20 mRNA相对表达量在癌组织中为(0.27±0.05),低于正常肺组织的(1.40±0.09)(t=2.768,P<0.05)。见图1。

图1 癌组织和正常肺组织中mRNA相对表达量比较
2.2组织中SETDB1和SPG20蛋白表达(SP法):免疫组化结果显示,SETDB1在癌组织中呈高表达,阳性表达率为81.67%,高于正常肺组织的26.67%(P<0.05);SPG20在癌组织中阳性表达率为28.33%,低于正常肺组织中的61.67%(P<0.05)。见表1、图2。
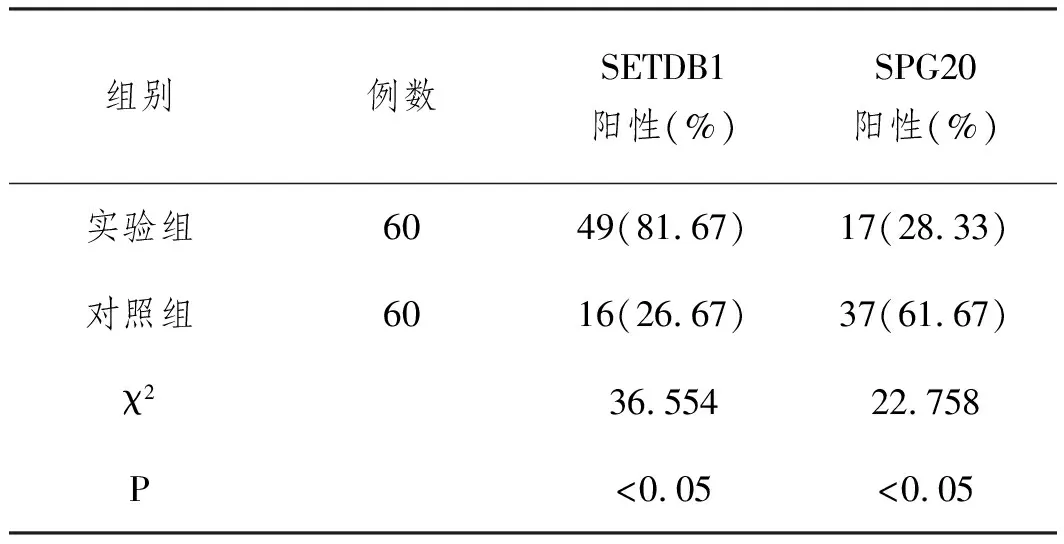
表1 SETDB1和SPG20在组织中的表达差异

图2 癌组织和正常肺组织中蛋白的表达比较
(SP-HE ×40)A:SETDB1在癌组中高表达; B:SETDB1在正常肺组织中呈低表达;C:SPG20在癌组中呈低表达;D:SPG20在正常肺组织中呈高表达
2.3组织中SETDB1和SPG20蛋白表达:Western blot结果显示,SETDB1在癌组织中表达量为(84.16±3.41)%,高于正常肺组织的(34.76±2.37)%(t=4.425,P<0.05);SPG20在癌组织中表达量为(16.37±2.04)%,低于正常肺组织的(62.64±1.41)%(t=3.742,P<0.05)。见图3,4。

图3 SETDB1在癌组织和正常肺组织中的表达量
2.4组织中SPG20甲基化水平:焦磷酸测序法检测显示,癌组织中SPG20甲基化率为(64.75±4.21)%,高于正常肺组织的(15.36±2.57)%(t=2.354,P<0.05)。见图5。

图5 组织中SPG20甲基化水平差异
2.5癌组织中SETDB1蛋白和SPG20甲基化的相关性:癌组织中SETDB1蛋白表达和SPG20甲基化水平具正相关,其相关系数r=0.642 (P<0.05);见表2。

表2 癌组织中SETDB1蛋白与SPG20甲基化水平的相关性(n)
2.6癌组织中SETDB1蛋白和SPG20甲基化与临床病例特征的相关性:统计分析结果显示,SETDB1蛋白阳性表达和SPG20甲基化与TNM分期、组织分化程度及淋巴结转移密切相关(P<0.05);与性别和年龄无关(P>0.05)。见表3。
2.7细胞系中SETDB1和SPG20的蛋白表达:Western blot结果显示,SETDB1在A549细胞中呈高表达,而SPG20呈低表达(t=2.562,P<0.05);正常支气管上皮细胞中SETDB1呈低表达,SPG20则呈高表达(t=5.143,P<0.05)。见图6。
2.8siRNA干扰SETDB1效果验证:转染成功后,细胞内可见较强的绿色荧光(图7)。qRT-PCR结果显示,si-S组SETDB1 mRNA相对表达量为(0.35±0.04),显著低于si-N和NC组[(1.27±0.13)和(1.31±0.14)](F=26.4,P<0.05,表4,图8)。si-S组SETDB1蛋白表达量显著低于si-N和NC组(P<0.05),见图9。
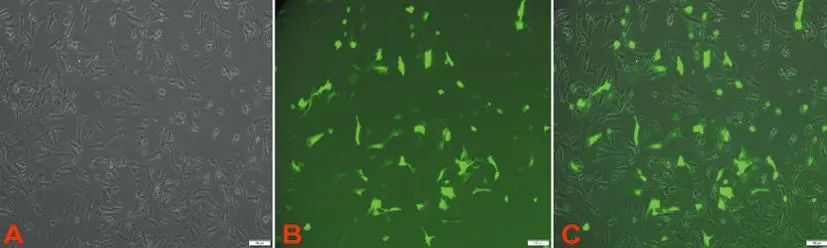
图7 RNA干扰SETDB1转染荧光效果
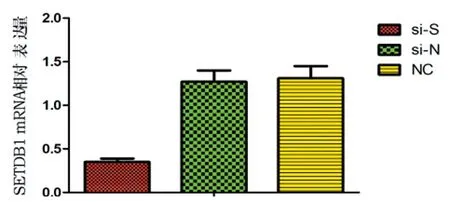
图8 RNA干扰后各组SETDB1 mRNA相对表

图9 RNA干扰后各组SETDB1蛋白表达量
2.9下调SETDB1对SPG20甲基化和蛋白表达的影响 RNA干扰下调SETDB1后,si-S组SPG20甲基化水平显著下降(F=29.5,P<0.05),而si-N和NC组甲基化水平无差异(P>0.05);见表5。si-S组SPG20蛋白表达量升高,显著高于si-N组及NC组(P<0.05)。见图10。
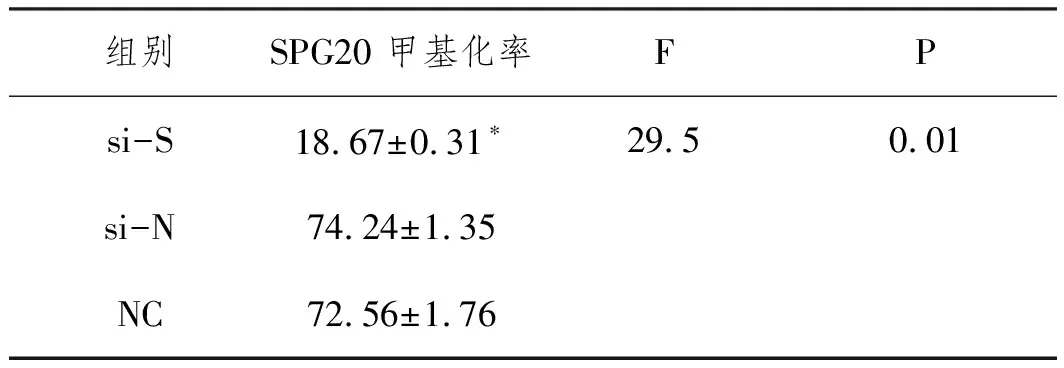
表5 下调SETDB1后各组SPG20甲基化率差异(%)
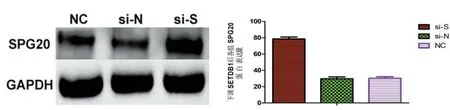
图10 下调SETDB1后各组SPG20蛋白表达量
2.10下调SETDB1对肺腺癌细胞功能的影响 MTT结果显示,在12h、24h、36h和72h时间点,si-S组细胞570nm吸光度值均小于si-N组和NC组,细胞存活能力下降(P<0.05);而si-N组和NC组相比无统计学差异(P>0.05)。见表6,图11.A;划痕愈合实验结果显示,与si-N组和NC组迁移距离相比,si-S组细胞迁移距离短(P<0.05);见表7,图11.B;侵袭实验结果显示,与si-N组和NC组相比,si-S组侵袭细胞数显著减少(P<0.05);见表8、图11。
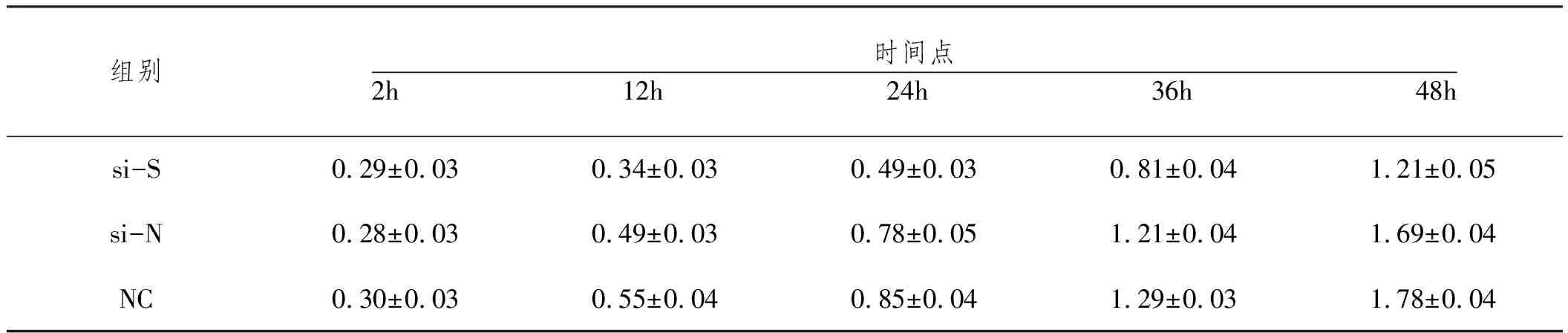
表6 下调SETDB1后各组细胞不同时间点吸光度值

表7 下调SETDB1后48h后各组细胞迁移距离(μm)

表8 下调SETDB1后48h后各组细胞穿膜细胞数

图11 下调SETDB1对肺腺癌细胞存活能力、迁移和侵袭能力的影响
3 讨 论
早期肺腺癌多无症状,部分患者确诊时已是进展期,从而导致其总体预后不佳[5]。因此,如何提高肺癌患者总体生存率,改善其预后具有十分重要的临床意义。
SPG20基因也被称为SPART,最初的研究表明其参与了Troyer综合征。然而最新研究表明,SPG20参与了肿瘤组织内表皮生长因子受体的运输和线粒体的能量代谢过程。研究证实,SPG20在胃癌中呈现低表达状态,这可能与胃癌组织中SPG20高甲基化相关[6]。SPG20在结直肠癌和肝癌中低表达,且其高甲基化修饰状态作为早期事件已经被证实[7]。课题组前期通过免疫组化实验证实,SPG20在肺腺癌组织中内呈低表达,这与SPG20在胃癌、结肠癌等消化道肿瘤研究中的低表达状态一致[8]。本研究通过免疫组化和蛋白质免疫印迹实验检测肺腺癌组织标本,结果显示SPG20在癌组织中呈低表达,而正常肺组织则呈高表达,差异显著。通过基因甲基化分析,结果表明肺腺癌组织中SPG20基因呈高甲基化。统计结果分析显示,SPG20高甲基化与肺腺癌TNM分期、分化程度以及淋巴结的转移具有相关性;SPG20的阳性表达随着癌组织TNM分期、组织程度分化的增加而逐渐降低,且有淋巴结转移的阳性表达明显降低。这说明SPG20在肺腺癌的发生发展过程可能起到“抑癌基因”作用,其低表达状态减弱抑癌作用,进而促进了肺腺癌的发生和发展。
甲基化修饰需要甲基转移酶的参与,甲基化转移酶在甲基化修饰过程中的作用至关重要。多项研究表明,甲基转移酶参与了多种肿瘤的增殖、侵袭和转移[9]。SETDB1是甲基转移酶家族中重要一员,参与了肿瘤中多个基因的转录和沉默。SETDB1具有调控胚胎干细胞生长和增殖、调节小鼠软骨发育等多种生物学功能。近年来的研究发现,SETDB1在人体的多种癌症如结肠癌、肝癌、乳腺癌等中均表达上调,与肿瘤的恶性程度和转移程度密切相关[10-11]。本研究通过免疫组化及蛋白免疫印迹检测肺腺癌中SETDB1水平,结果显示,SETDB1在癌组织中呈阳性表达,与正常肺组织之间存在显著差异。统计分析结果显示,SETDB1的表达水平与肺腺癌TNM分期、组织分化程度及淋巴结转移密切相关;SETDB1的阳性表达随着肺腺癌TNM分期、癌组织分化程度的增加而增加,且有淋巴结转移的阳性表达明显增加,明显起到了“促癌”的作用。这与国外研究在非小细胞肺癌癌组织中SETDB1高表达状态,与肺癌之间具有强相关性的研究结果一致。
课题组前期研究发现,SPG20在肺腺癌组织中的低表达与其发生甲基化密切相关。研究证实,SETDB1作为甲基转移酶可以催化多种基因发生甲基化。在肺腺癌中SETDB1作为甲基转移酶与SPG20的甲基化修饰是否相关,及二者在肺腺癌中的相关性及相互作用机制鲜有研究。本研究对SPG20甲基化和SETDB1在癌组织中的相关性分析结果显示,肺腺癌组织中SETDB1高表达而SPG20甲基化呈高水平,二者呈正相关。进一步检测肺腺癌细胞系中SETDB1和SPG20蛋白的表达,结果证实,肺腺癌细胞系中SETDB1在肺腺癌细胞中呈高表达,而SPG20呈低表达;正常支气管上皮细胞中SETDB1呈低表达,SPG20则呈高表达,这与肺腺癌组织中检测结果一致。进一步通过体外细胞实验验证甲基化转移酶SETDB1与SPG20甲基化的相关性,结果显示,RNA干扰下调SETDB1后SPG20基因甲基化率明显降低,SPG20蛋白表达均明显上升。结合SPG20甲基化和SETDB1二者相关性结果,笔者推测在肺腺癌中SETDB1作为甲基转移酶促进了SPG20的甲基化,进而抑制了SPG20的表达,使得肺腺癌的恶性程度增加。今后研究中可以将SETDB1及SPG20甲基化作为切入点,深入研究二者具体致癌机制,为肺腺癌的基因靶向治疗提供新的理论依据。
综上所述,SPG20甲基化和SETDB1与肺腺癌的分期、分化程度及转移密切相关,且二者呈正相关性,二者可能协同作用促进了肺腺癌的发生发展,在肺腺癌诊治中具有潜在的应用价值。
猜你喜欢
保健与生活(2022年11期)2022-06-09
生物学通报(2020年10期)2020-08-13
中国报道(2018年2期)2018-04-20
金融博览(2016年7期)2016-08-16
国际妇产科学杂志(2016年2期)2016-06-16
绿色中国(2016年1期)2016-06-05
现代检验医学杂志(2015年2期)2015-02-06
沈阳医学院学报(2014年4期)2014-12-27
遗传(2014年3期)2014-02-28
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
