
基于表型性状构建禾初级核心种质
2024-01-30 12:19毛名义杨文荟管艳伟潘宗东周丽洁余显权
中国稻米 2024年1期
毛名义 杨文荟 管艳伟 潘宗东 周丽洁* 余显权*
(1 贵州大学农学院,贵阳 550025;2 贵州大学水稻产业技术研究院,贵阳 550025;3 黔东南州农业科学研究院,贵州 凯里 556000;第一作者:244745381@qq.com;*通信作者:zhoulijie2006@163.com;yxqgy@126.com)
贵州独特的地形地貌和生态环境孕育了丰富的稻种资源。“禾”是贵州省黔东南山区侗族等少数民族生活的独特生态环境下经长期自然演变和人工选择所形成的一类地方性水稻品种,具有地域性强、耐冷、耐阴、耐贫瘠环境和耐烂锈田等特点[1]。在禾类资源中具有浓郁芳香气味、质佳味美的香禾糯是黔东南侗族等少数民族糯食文化的载体,素有“一家蒸饭满寨飘香”的美誉。禾在品质和抗性等方面的优良特性及蕴含的大量优异基因是当前水稻遗传改良中亟需的基因源,具有重要的利用价值。种质资源是品种遗传改良的重要物质基础,大量收集和保存种质资源是挖掘和利用特异新种质的基础[2]。然而,数量庞大的资源群体也给种质保存与研究利用带来一定困难,构建核心种质不仅可以降低资源保存和管理工作的投入和费用,也能提高特异种质筛选和利用的效率。
1984 年,FRANKEL[3]率先提出“核心种质”概念,之后相关的研究报道逐年增多。核心种质是指以最少的资源数量和遗传重复,最大程度代表整个遗传资源的多样性,核心种质的建立在保证遗传多样性的同时又减少资源数量,避免了因不必要的保存而对资源造成的浪费[4]。近年来,国内外学者基于农艺性状或分子标记成功构建了水稻[5-8]、玉米[9]、小麦[10]、高粱[11]、甘薯[12]、番茄[13]、马铃薯[14]等作物的核心种质。禾作为一类特殊的气候生态类型水稻,其独特的民族文化内涵、遗传多样性和优良品质引起了学者们的关注[15-17]。刘春晖等[18]在基因组层面揭示禾的遗传演化规律和基因组印记,并与社会科学交叉融合,研究表明,禾起源于福建、广东、广西和江西,随侗族祖先的迁徙传入黔东南。多样化的地理起源和黔东南山区复杂的自然环境蕴育了丰富的禾资源,而对禾核心种质构建的研究鲜有报道。本研究以来源于黔东南地区黎平、从江、榕江三地和广西三江县的402 份禾种质资源为材料,探究禾核心种质构建的最佳取样策略,并对构建的禾初级核心种质进行评价,以期为禾种质资源的保存、特异资源挖掘和高效利用提供理论依据。
1 材料与方法
1.1 试验材料种植
402 份禾品种资源材料由贵州省黔东南州农业科学院提供,来源地如下:贵州省黔东南州黎平县228份、从江县158 份、榕江县11 份,广西三江县5 份。供试材料于2022 年在黔东南州从江县高增乡高增村种植,4 月28 日播种,6 月7 日移栽,每份材料种植15 行,每行10 株,行距30 cm,株距10 cm,田间肥水管理及病虫害防治均按当地禾种植习惯进行。
1.2 表型统计及赋值
参照韩龙植[19]《水稻种质资源描述规范和数据标准》对9 个质量性状(穗枝集散度、穗型、倒伏性、谷粒形状、芒色、种皮颜色、熟期性、颖尖色、颖色)按表1 进行统计与赋值;17 个数量性状(株高、穗长、每穗总粒数、每穗实粒数、结实率、一次枝梗数、谷粒长、谷粒宽、谷粒长宽比、千粒重、剑叶长、剑叶宽、倒2 叶长、倒2叶宽、单株穗数、芒长、产量)按平均值(X)和标准差(δ)分为10 级,1 级<X-2δ,10 级≥X+2δ,中间每级相差0.5δ。

表1 禾9 个质量性状分级与赋值
1.3 核心种质构建策略
1.3.1 表型分组及聚类分析
基于禾颖尖颜色差异,将402 份禾种质材料分为5 组(表2),对26 个表型数据进行标准化处理后采用欧式距离结合离差平方和法以组为单位利用SPSS 20.0 软件进行聚类分析。根据聚类图结果,从遗传距离最小的一组中选择具有特殊表型性状的一份材料进入下一轮聚类,在剔除其余材料后并对入选材料再次聚类,直至剩余材料达到需要的取样量。
1.3.2 取样策略分析
在分组及逐步聚类基础上,利用变异系数变化率、极差符合率、表型保留比例和多样性指数4 个参数检验以4 种组内取样比例(简单比例法、对数比例法、平方根比例法和多样性比例法)结合6 种总体取样规模(5%、10%、15%、20%、25%和30%)形成的24 种取样策略(表3),筛选最佳取样策略。

表3 候选核心种质评价参数
1.3.3 核心种质评价
筛选出最佳取样策略后,基于原始种质与核心种质的26 个表型性状,利用最大值、最小值、均值、变异系数和多样性指数5 个特征值检验在最佳策略下构建的核心种质变异范围及均匀度。采用均值差异百分率、方差差异百分率、极差符合率、变异系数变化率和表型保留比例5 个参数评价核心种质的代表性。当核心种质的均值差异百分率小于20%且极差符合率大于80%说明构建的核心种质具有代表性,同时变异系数变化率、方差差异百分率和表型保留比例等参数值越大越能代表原始群体遗传多样性[20-21]。
2 结果与分析
2.1 总体取样规模和组内取样比例分析
基于禾颖尖颜色差异,将402 份禾种质资源分为5 组(褐色Ⅰ、黄色Ⅱ、秆黄色Ⅲ、黑色Ⅳ、紫色Ⅴ)在6种(5%、10%、15%、20%、25%和30%)总体取样规模下,以4 种组内取样比例方法(简单比例、平方根比例、对数比例和多样性比例)分别计算24 种策略下每组取样数量(表2)。由表2 可知,禾颖尖颜色中褐色在原始种质中数量最多(190 份),然后依次为黄色(138 份)、秆黄色(30 份)、黑色(23 份)和紫色(21 份)。褐色在简单比例法中取样率高达47.26%,在对数比例、平方根比例和多样性比例中取样率分别为26.56%、34.13%和28.73%。紫色颖尖在简单比例法取样率最低为5.22%,但在另外3 种比例法下取样率均有所提高,其中对数比例法(15.41%)>多样性比例法(12.51%)>平方根比例法(11.35%)。综上可知,对数比例与多样性比例二者能增加稀有颖尖色的取样份数,让各组的取样数量更加均匀,对禾资源中稀有颖尖色(黑色、紫色)有一定的校正效果。
为获得最佳总体取样规模及组内取样比例方法,选取变异系数变化率、极差符合率、表型保留比例和多样性指数4 个参数进行综合分析比较,结果(表3)表明,4 种组内取样比例法中,简单比例法、平方根比例法和多样性比例法在25%和30%取样规模下变异系数变化率小于100%,而对数比例法在6 种取样规模变异系数变化率均大于100%。极差符合率随取样规模增加而变大,简单比例、平方根比例、多样性比例和对数比例在5%的取样规模下分别为73.26%、71.26%、78.12%和80.10%,仅对数比例的极差符合率大于80%。在10%的取样规模下对数比例极差符合率大于简单比例,其余两种比例法均小于80%。平方根比例与多样性比例在15%的取样规模下才达到80%。简单比例法、对数比例法和多样性比例法其多样性指数在10%的取样规模下达到最大,而平方根比例在15%取样规模时才达到最大,多样性指数随着取样规模的增大出现先升高后降低的趋势。表型保留比例随着取样规模的增加而增加,当取样规模>10%后,其增幅不大,但多样性指数却在逐步降低,说明过度的筛选反而会让核心样本各性状的变异量下降,在剔除原始种质中冗余的样品时应适度。综上所述,表明4 种组内取样比例法中对数比例法最好,6 种取样规模中10%的取样规模也最适宜。
2.2 组内取样方法的确定
为进一步明确组内取样方法中聚类和随机取样的效果,按对数比例法下10%和15%的取样规模进行分析比较。结果(表4)显示,在10%和15%的取样规模下,以聚类法进行组内取样时,其变异系数变化率、极差符合率、表型保留比例和多样性指数4 个检验参数均较随机取样大,说明组内取样方法中聚类法能提升核心种质变异的均匀度与丰富度。
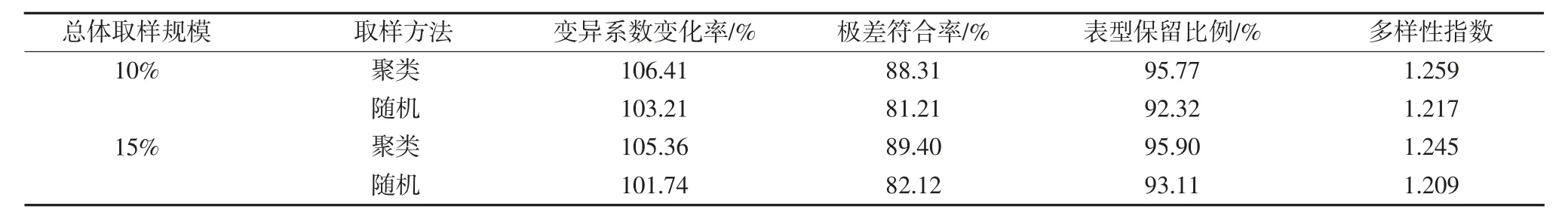
表4 组内不同取样方法的比较
2.3 核心种质评价
2.3.1 原始种质和核心种质各性状特征值比较
为明确所构建的核心种质与原始种质各性状特征值之间的差异,统计了核心种质与原始种质2 个样本26 个农艺性状的最大值、最小值、均值、变异系数和多样性指数。由表5 可知,调查的9 个质量性状(穗枝集散、穗型、倒伏性、芒色、种皮颜色、粒型、熟期性、颖尖色和颖色)原始种质与核心种质的最大值与最小值一致。穗长、倒2 叶长、单株穗数、结实率、一次枝梗数、谷粒长、谷粒长宽比、倒伏性、穗型、粒型和穗枝集散等性状原始种质与核心种质的均值差异不大,原始种质与核心种质的变异系数差异较小,二者的多样性指数也相对一致。综上,原始种质与核心种质大部分特征值相等或者差异不大,说明构建的核心种质能较好的代表原始种质。
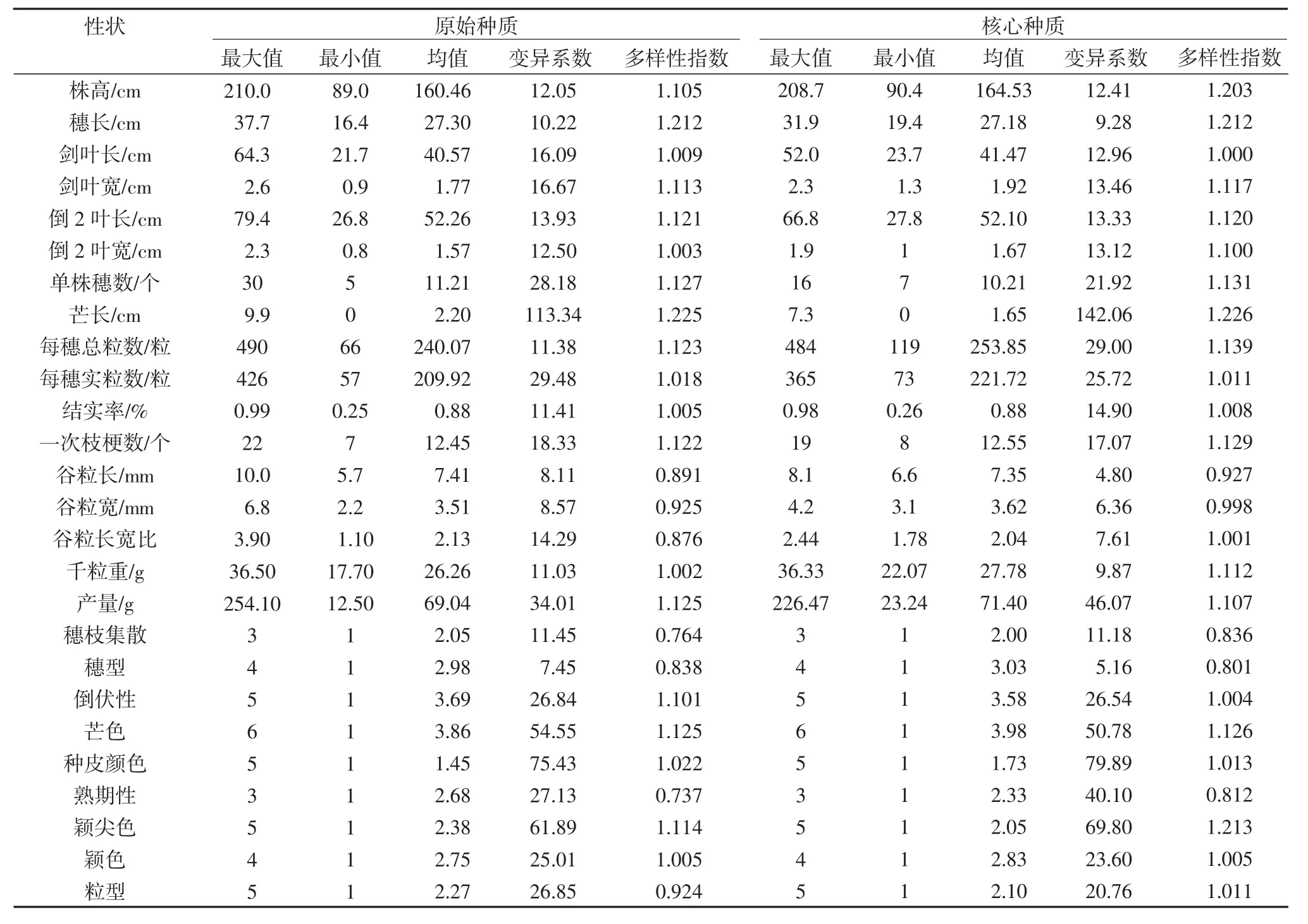
表5 原始种质与核心种质特征值的比较
2.3.2 核心种质评价参数分析
由表6 可知,本研究构建的核心种质的均值差异百分率为12.70%,极差符合率为88.31%,符合均值差异百分率小于20%且极差符合率大于80%这一核心种质构建标准,表明构建的核心种质具有代表性,其变异系数变化率为106.41%,表型保留比例为95.77%和方差差异百分率为23.50%。核心种质26 个性状中,20 个性状极差符合率大于80%,5 个性状接近80%,只有倒2 叶长较小(66.48%)未达到预期效果。核心种质变异系数变化率中有21 个性状大于100%。株高、穗长、剑叶长等19 个性状的表型保留比例均为100%,其余7个性状均大于90%。综上所述,构建的核心种质具有代表性,能较好代表原始种质。
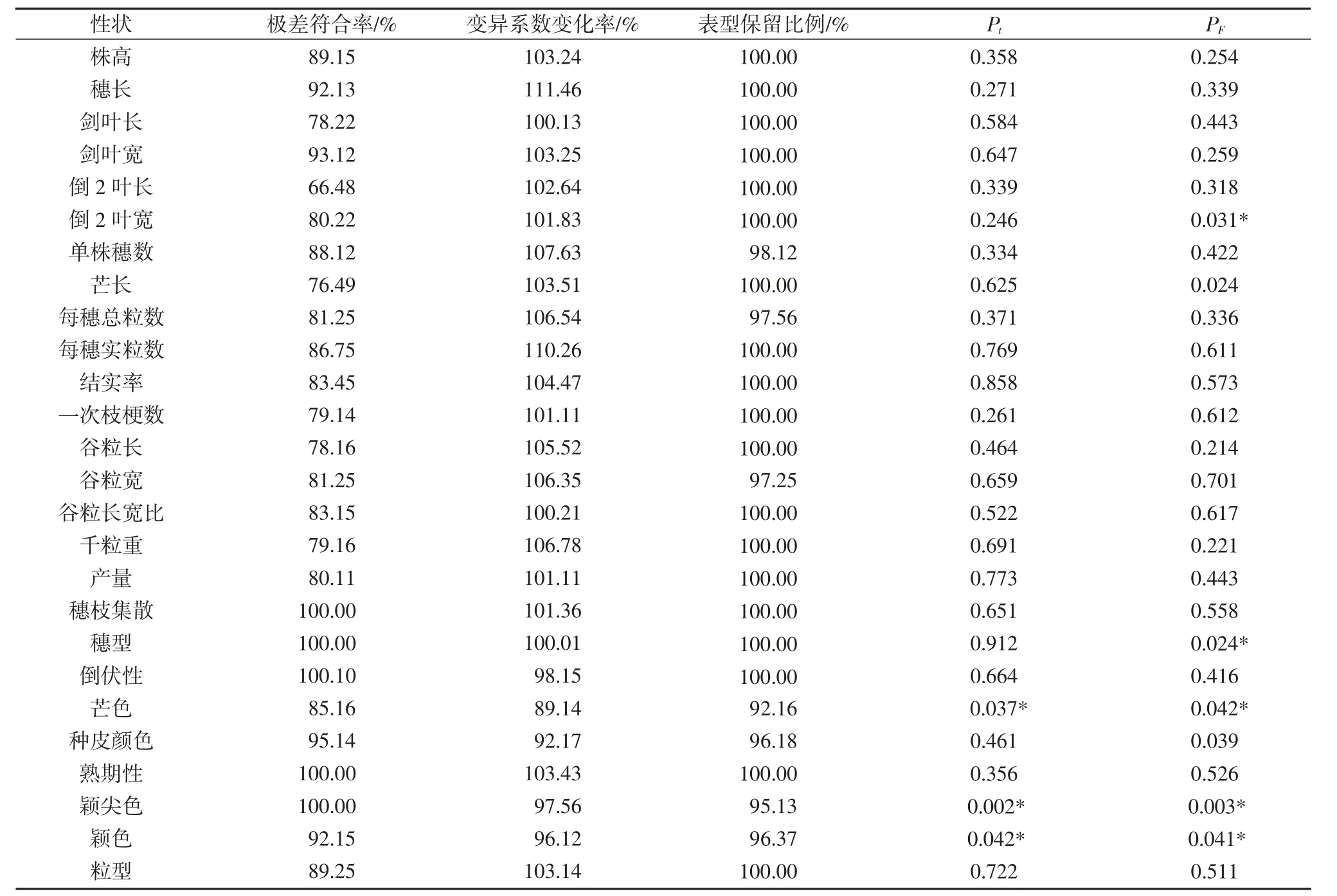
表6 核心种质与原始种质26 个性状评价参数和t 检验及F 检验
2.3.3 26 个性状均值的t检验和F检验
对构建的初级核心种质与原始种质各性状均值进行t检验,由表6 可知,26 个表型性状有3 个性状具有显著性差异,分别为芒色、颖色和颖尖色,这是由于按颖尖色分组后,对数比例法取样的修正作用在一定程度上提高了稀有颖尖色的取样数量,使核心种质丰富度得以提高。F检验结果表明,倒2 叶宽、穗型、芒色、颖尖色和颖色5 个性状具有显著差异,说明核心种质中这些性状保留了更多的遗传变异且分布较均匀,遗传冗余度较小。
3 讨论与结论
随着高产杂交水稻大面积推广以及人们对禾种质资源保护意识淡薄等原因,导致黔东南地区禾种植面积减少,禾种质资源流失严重。地方水稻品种是我国水稻种质资源的重要组成部分,较选育品种具有更丰富的遗传多样性。禾作为黔东南山区一类特殊生态型地方水稻品种,在抗逆方面表现突出,并具有糯性强、香味浓等特性,是当前水稻遗传改良中选择特异性基因材料的重要来源。种质资源承载着作物的优良基因,是育种的前提与物质基础。大量研究表明,在水稻遗传改良过程中选育出的某些突破性水稻品种或优良双亲类型都与种质资源密不可分,其也是评判一个国家农业综合实力的标准之一[22-23]。如何快速准确的从种质资源中挖掘出可以为水稻遗传改良服务的优异基因是育种工作中急需解决的现实问题,而基于表型性状构建核心种质成为种质资源开发及种质创新中一种高效且实用的方法。
选择合适的分组方法对核心种质构建尤为重要,常见分组方法有单一性状、来源地、遗传标记与育种体系等。李国强等[24]在构建大白菜核心种质研究中,以白菜表型性状中的结球变种进行分组。胡建斌等[25]与徐宁等[26]在构建1 200 份甜瓜与4 877 份小豆核心种质时均按材料来源地进行分组。李自超等[27]在构建云南地方稻初级核心种质过程中提出了水稻分组体系,发现丁颖分类体系>省+籼粳体系>单一性状分组>稻作区分组>不分组。本研究在借鉴前人分组方法上基于禾颖尖颜色的不同分为5 组,用颖尖色分组较用其他性状分组的组间数量差异小,能提高禾资源中那些稀有颖尖色的取样数量,使取样更加均衡。
不同的组内取样比例方法也对核心种质代表性产生影响。李嘉伟等[28]在基于表型构建传统菊花核心种质中,比较了4 种组内取样比例法的优劣,发现对数比例法>多样性比例法>平方根比例法>简单比例法。李慧峰等[29]基于4 种比例法构建甘薯核心种质,结果表明,在对数比例法下构建的核心种质其效果最佳,平方根比例法最差。李自超等[30]在研究云南地方稻种资源取样策略中发现,组内取样比例中对数比例法和平方根比例法效果最好,多样性比例法次之,简单比例法最差,因为对数比例法和平方根比例法对那些数量基数较小的组别可相对提高其取样数量,一定程度上提高了某些小组别的遗传重复。
前人在构建不同作物核心种质时发现,核心种质的数量大约占原始种质的5%~30%,且大多集中于10%左右。BROWN[31]认为,以原始种质数量的5%~10%比例下构建的核心种质就能保留住原始种质70%以上的遗传变异。李自超等[27]认为,原始种质样本数量的多少可用于判定核心种质占原始种质的比例,一般来说,原始种质样本数量大的其核心种质所占的比例偏小,原始种质样本数量小的其核心种质所占的比例偏大。李秀诗等[32]以248 份薏仁原始种质构建核心种质,发现最佳取样比例为25%。胡建斌等[25]基于1 200 份甜瓜种质材料构建核心种质中发现,在15%的取样比例,其各个检验指标均达到最佳水平。WANG 等[33]以2 794 份大豆为材料构建核心种质,其取样比例仅为2%。同时,李国强等[24]认为,考察农艺性状的多少也能决定取样比例的大小,若考察的农艺性状偏少,各种遗传变异不能得到充分体现,为避免丢失重要变异类型,在构建核心种质过程中应该适当增加表型性状的调查数目,不同作物考察表型性状数量也不同,一般来说在10~30个。本研究中所考察的26 个表型性状是基于前人对粳稻、野生稻或地方稻等核心种质构建结果,该数量表型性状能较好反映禾表型变异的同时对表型数据有效性进行保证。本研究较前人基于表型构建核心种质的原始材料数量上相对偏少,为保证取样科学性与合理性,设置5%、10%、15%、20%、25%和30%6 种总体取样规模,经分析对比后,10%取样规模下所构建的核心种质遗传多样性指数最大,4 种组内取样比例法其表型保留比例都大于90%,其中对数比例法表型保留比例为95.77%,说明能充分代表禾原始种质的遗传多样性。对比分析在10%和15%的总体取样规模下以随机和系统聚类两种取样方式的效果,均以聚类取样为宜,与前人的研究结果相似[27-28]。基于不同作物各性状变异结构分布不均衡情况,采用随机取样可能会丢失某些重要种质资源,故在构建核心种质过程中大多采用系统聚类取样来降低偏差。
核心种质的代表性是检验核心种质构建成败的重要环节,评价其代表性应根据不同作物属性选择合适的评价参数[34]。检验核心种质代表性仅以单一参数进行评价,其反馈结果具有局限性,应综合多个参数对核心种质代表性进行考量。本研究借鉴均值差异百分率、方差差异百分率、极差符合率、变异系数变化率和表型保留比例5 个前人高频使用的评价参数对构建的核心种质进行评价,结果表明,构建的47 份禾核心种质均值差异百分率和极差符合率分别为12.70%和88.31%,说明构建的核心种质具有代表性。变异系数变化率、方差差异百分率和表型保留比例作为评价核心种质的重要参数,值越大越能代表原始种质的遗传信息,47 份禾核心种质的3 个评价参数分别为106.41%、23.50%、95.77%,与前人研究结果中3 个参数变化范围一致[35-37],能较好代表原始种质遗传信息。本研究构建的核心种质的表型保留比例为95.77%,出现丢失部分原始种质表型的情况,因此,保存原始种质中扣除核心种质外的那部分种质资源可作为核心种质补充来源,同时也要不断搜集新的优异种质为核心种质更新换代作铺垫。综上,本研究构建的核心种质在表型水平上具有较好代表性,遗传冗余度较小,种质规模适宜,其作为原始种质的“精华”可优先加以利用。
本研究以收集402 份禾种质资源为材料,依据禾颖尖色差异将原始种质划分为5 组,通过比较分析不同的取样策略,结果显示,10%为最适总体取样规模,组内取样比例法以对数比例法最优,聚类取样法是组内最佳取样方法。在最优方案下构建了47 份禾核心种质,占原始材料11.69%。5 个评价参数均值差异百分率、极差符合率、变异系数变化率、方差差异百分率和表型保留比例分别为12.70%、88.31%、106.41%、23.50%和95.77%。结果表明,构建的核心种质具有较好的丰度和均度,能较好代表原始种质。
猜你喜欢
今日农业(2022年13期)2022-09-15
Journal of Geriatric Cardiology(2021年1期)2021-03-03
影像研究与医学应用(2019年9期)2019-04-24
中国麻业科学(2018年6期)2018-04-09
现代园艺(2017年21期)2018-01-03
中国实验诊断学(2017年11期)2017-11-28
西南农业学报(2016年5期)2016-05-17
广西林业科学(2016年3期)2016-03-16
中国康复理论与实践(2015年10期)2015-12-24
医学研究杂志(2015年5期)2015-06-10
