
小兴安岭3种槭树不同生长期叶性状变异及相关性分析
2024-01-23 05:35彭仲韬郭嘉兴王艺璇金光泽刘志理
南京林业大学学报(自然科学版) 2024年1期
彭仲韬,郭嘉兴,王艺璇,王 磊,金光泽,刘志理*
(1.东北林业大学生态研究中心,森林生态系统可持续经营教育部重点实验室,东北亚生物多样性研究中心,黑龙江 哈尔滨 150040;2.东北林业大学林学院,黑龙江 哈尔滨 150040)
植物功能性状是植物对各种环境条件和生物相互作用的长期适应后所表现出能通过测量并以数值表示的特征[1-3]。叶片通过光合作用供给植物大部分的干物质来源,此外叶性状作为植物功能性状的重要部分,它还对植被特性产生影响[4-6]。环境的不同会导致叶性状发生巨大变化[7],所以叶性状的多样性反映了植物对资源获取、对各种胁迫所做出应对的多种方式[8]。常见的叶性状包括比叶面积(specific leaf area, SLA)、叶干物质含量(leaf dry matter content, LDMC)、叶绿素含量(chlorophyll, Chl)和叶厚(leaf thickness, LT)等。比叶面积反映了植物对养分的利用率和截获光照的能力,因此它被认为是能体现植物资源利用策略和竞争能力的重要参数[9-11];叶干物质含量通常和相对生长速率呈负相关,另外它还与叶片对资源的存储能力有关[12];叶厚能反映植物在干燥、高光照等恶劣环境中的生存能力[13];叶绿素含量则直接影响植物光合作用潜力[14]。
季节(叶生长期)的变化对植物获取资源、植物种群的更新等至关重要[15-17],但也要求植物在各种不利条件下充分利用对它们生长有利的各种外界条件[18]。对不同季节的叶性状进行定量研究有助于更好地解析叶生活史策略[19]。另一方面,因为植物的气候敏感性使其成为反映气候变化的重要工具,所以植物生长发育过程(如叶片的生长衰落、开花结果等)的动态变化近年来备受关注[5, 20]。随着叶片生长,森林内空气温度、湿度以及叶片孔隙度等指标会发生明显改变[21-22]。Basnett等[11]的研究证明随叶片生长,叶厚显著增加以支撑逐渐增大的叶片面积;McKown等[23]认为比叶面积随树木物候的变化而呈显著减小趋势;Croft等[24]的研究表明叶绿素含量从叶片生长期开始时缓慢增长直至叶片生长期中期保持稳定,最后在叶片衰老时迅速下降。但叶性状通常并非独立变化,而是存在一定的协调关系[2,6,10]。“叶经济谱”第1次比较系统地描述了这种关系:它的一端是高比叶面积和生长速率,低叶干物质含量的“快速投资-收益”型策略,为获取性策略;另一端则相反,表现为保守性的“缓慢投资-收益型”策略[4, 25]。“叶经济谱”的概念被提出后,关于叶性状间权衡关系方面的研究越发受到重视[3-4, 6],植物通过调整性状之间的相关关系形成最适合自身的生存策略。Fajardo等[26]认为物候变化不仅会导致叶性状发生变异,还能影响到性状之间的协作关系。但关于同属不同种植物的叶性状随叶生长是否表现出相似的变化规律,以及性状间的关系是否都会受到叶生长期的显著影响尚不清楚。
花楷槭(Acerukurunduense)、青楷槭(A.tegmentosum)和五角枫(A.pictumsubsp.mono)为槭树科槭属落叶阔叶植物,广泛分布于我国东北地区的阔叶红松(Pinuskoraiensis)林中,且这3种槭树总体上呈现混合交错的分布状态,生态位并无明显分化。本研究以小兴安岭内的花楷槭、青楷槭、五角枫作为研究对象,在展叶期(6月初)、叶稳定期(7月中旬)和叶凋落期(9月初)测定比叶面积、叶干物质含量、叶绿素含量和叶厚这4个重要叶性状,主要研究叶性状的生长动态及种间差异,探究性状间的相关性,并检验叶生长期对性状间相关性的影响机制,以期为揭示落叶阔叶植物的叶生活史策略提供科学依据。
1 材料与方法
1.1 研究区概况
野外调查在黑龙江凉水国家级自然保护区(128°53′20″E,47°10′50″N)进行。该区域以起伏的山地地形为主,最高海拔为707 m;该区属温带大陆性季风气候,年均气温-0.3 ℃,夏季(6—8月)平均气温17.5 ℃;年均降水量676 mm且主要集中在7月,此外该区域12月至翌年4月均被积雪覆盖;地带性顶极植被为阔叶红松混交林[28]。
1.2 样本采集
于2019年6月,在阔叶红松林分布区分别随机选取花楷槭、青楷槭和五角枫长势良好、胸径为3~5 cm的样树各3株,各样树立地的海拔、坡度、坡向相似,以排除地形因子对实验的影响;不同样树之间距离至少10 m,以防止小环境内各样树由于生长条件过于相似导致叶性状差异不明显。本研究以6月初、7月中旬及9月初分别代表展叶期、叶稳定期和叶凋落期。在每个生长期内,随机采集每株样树不同方位冠层未受到病虫害影响的完整样叶60片,即每种槭树在单个生长期共计采集180片样叶。将采集的样叶及时装入保温箱,带回实验室后即刻进行叶性状测定。
1.3 叶性状测定
将每片样叶表面擦拭干净,利用精度为0.01 mm的游标卡尺测定叶片的前端、中端和末端(注意避开主叶脉)3个点的叶厚度,取其均值作为该叶片的叶厚(leaf thickness,LT)。利用叶绿素仪(SPAD-502Plus)测定值表征样叶的叶绿素含量(Chl),获得样叶不同位置的测定值2次,精确到0.1%,并取其均值作为该样叶的叶绿素含量(该过程同样避开主叶脉)。利用扫描仪(Epson PerfectionV700 Photo,日本)扫描所有样叶,再结合ImageJ软件[29]计算每片样叶的比叶面积(specific leaf area, SLA),精确到0.001 cm2。在黑暗条件下将样叶置于清水中浸泡24 h后取出。用滤纸把叶表面的水擦净,利用电子天平(精度为0.000 1 g)称量叶饱和鲜质量。然后,在75 ℃条件下把样叶烘干至恒定后获得叶干质量(精度为0.000 1 g)。通过叶面积和叶干质量的比值获得比叶面积,通过叶干质量和叶饱和鲜质量的比值获得叶干物质含量(LDMC)。
1.4 统计分析
采用最小显著性差异(LSD)法检验3种槭树的叶性状,以及每种槭树在不同叶生长期叶性状之间的差异性。为使数据符合正态分布,分析时对所有数据进行对数转化。利用嵌套方差分析和方差分解来评估不同因子对叶性状变异的解释率。嵌套水平为生长期>树种>个体。进行各性状Pearson相关性分析,显著性水平均设定为P=0.05。对于相关性显著的性状组合,利用标准主轴分析法(standardized major axis,SMA)检验性状之间的相关关系是否因树种和生长期的不同而显著变化。如果性状间相关关系的斜率随生长期不同无明显变化,则计算其共同斜率,这说明生长期变化对性状间的相关关系无显著影响。最后利用主成分分析法(principal component analysis,PCA)探究不同种槭树间及不同叶生长期多性状间的协作关系。统计分析采用R软件[30],并利用SigmaPlot 14.0和R3.2.5进行绘图。
2 结果与分析
2.1 叶性状随生长期的变异
树种水平上,五角枫的叶干物质含量、叶绿素含量显著高于青楷槭和花楷槭,花楷槭的叶厚显著高于其他两种槭树,且花楷槭和五角枫比叶面积的差异不显著,但均显著高于青楷槭(图1),叶性状变异的方差分解见图2。
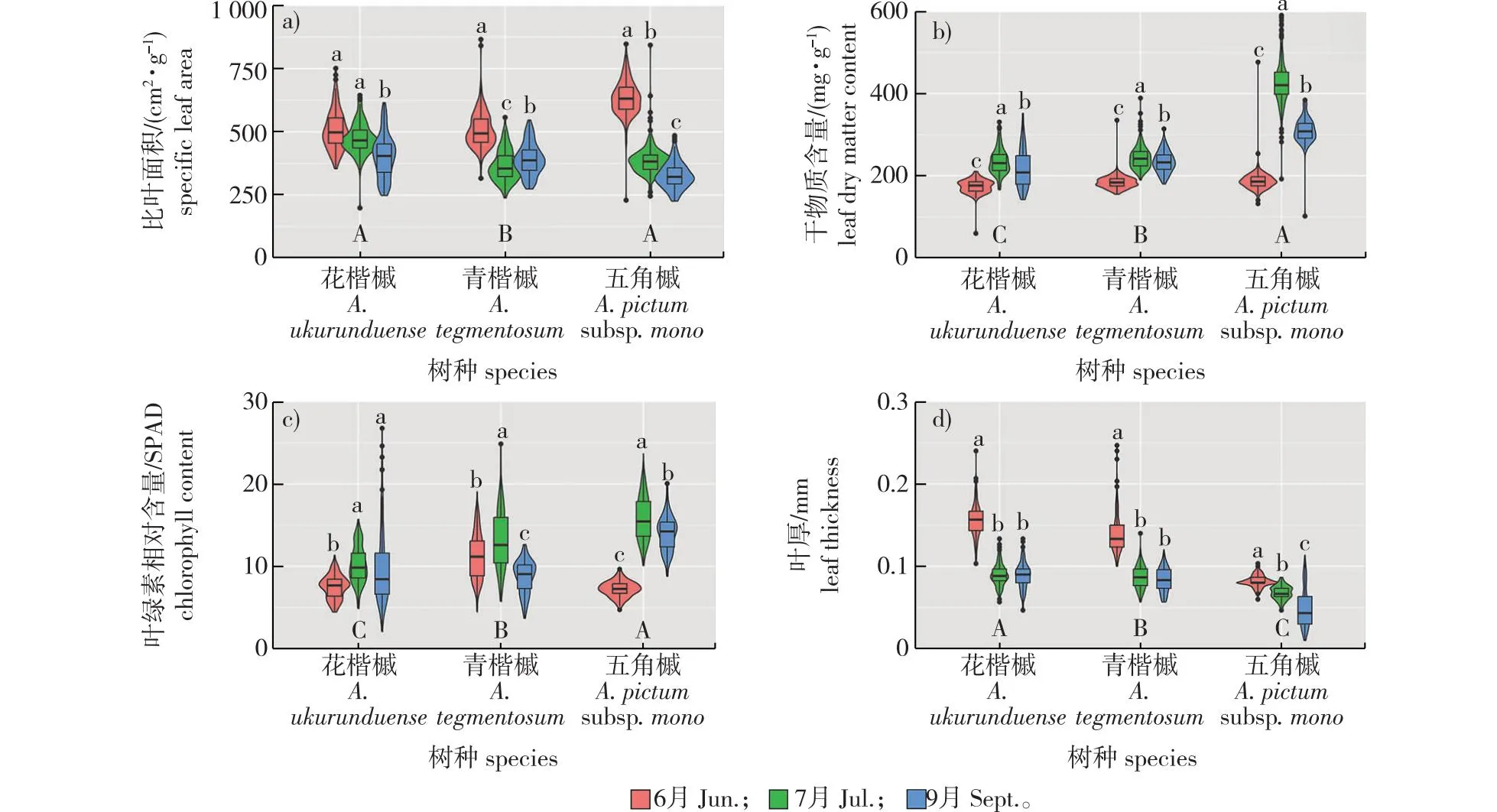
不同大写字母表示不同树种水平上叶性状间存在显著差异(P<0.05),不同小写字母表示同种槭树不同生长期的叶性状间存在显著差异(P<0.05)。Different uppercase letters meant significant difference among leaf traits of different species at 0.05 level, different lowercase letters meant significant difference among leaf traits of different leaf growth periods for each species at 0.05 level.图1 不同生长期3种槭树叶性状的变异Fig. 1 Violin plots for leaf traits of three Acer species in different growth periods

图2 叶性状变异的方差分解Fig. 2 Variance partitioning of leaf traits variations for three Acer species
随叶生长期的变化,3种槭树叶稳定期的叶干物质含量显著高于展叶期和叶凋落期(图1b),而展叶期的叶厚(LT)显著高于其余两个时期(图1d)。花楷槭叶稳定期(7月)和叶凋落期(9月)的叶绿素含量与叶厚(图1c、1d)以及青楷槭叶稳定期和叶凋落期的叶厚差异不显著(图1d);五角枫展叶期的比叶面积和叶厚(图1a、1d),叶稳定期的叶干物质含量和叶绿素含量显著高于其他两个时期(图1b、1c)。3种槭树在展叶期(6月)的叶干物质含量多集中分布于200 mg/g左右(图1b),花楷槭和五角枫在展叶期的叶绿素含量集中分布于7.5 SPAD左右(图1c)。生长期和树种对4个叶性状变异的解释率之和均超过了60%,其中对于叶干物质含量的解释率之和达到了91%(图2)。除叶绿素含量受个体间差异影响较大外(Chl为32%),个体对其他3个叶性状变异的解释率均小于18%(图2)。
2.2 叶性状间的相关性
3个树种各叶性状之间均存在显著或极显著的相关关系。其中:比叶面积与叶干物质含量、叶绿素含量均存在极显著负相关关系(相关系数分别为-0.667**、-0.667**,P<0.01),比叶面积与叶厚存在极显著正相关关系(相关系数为0.515**,P<0.01);叶干物质含量与叶厚存在极显著负相关关系(相关系数为-0.647**,P<0.01),与叶绿素含量存在极显著正相关关系(相关系数为0.761**,P<0.01);叶厚与叶绿素含量存在显著负相关关系(相关系数为-0.421*,P<0.05)。
在树种水平上,3种槭树除叶绿素含量和叶厚之间存在共同斜率(-0.964;P=0.452,图3 a)外,其他叶性状间相关关系的各斜率之间均差异显著(P<0.001),即各叶性状之间的相关关系随树种不同而产生变异。其中:比叶面积随叶厚的变异速度为青楷槭>五角枫>花楷槭(图3b);比叶面积随叶干物质含量的变异速度为青楷槭>花楷槭>五角枫(图3c);比叶面积随叶绿素含量变异速度为五角枫>青楷槭>花楷槭(图3d)。3种槭树的叶干物质含量与叶厚、叶绿素含量之间分别呈极显著负相关和极显著或显著正相关关系,且回归斜率绝对值均为五角枫>青楷槭>花楷槭(图3e、3f)。
花楷槭除比叶面积和叶绿素含量之间斜率在3个叶生长期间差异显著(P<0.001;图4a1)外,剩余性状间相关关系的斜率在不同叶生长期间均差异不显著(P>0.05)。青楷槭除叶厚和叶绿素含量之间的斜率在3个叶生长期间差异不显著(P>0.05)外(图4b2),其他性状间均差异显著。其中叶干物质含量和叶绿素含量在叶稳定期和叶凋落期表现为正相关,且回归斜率为叶稳定期>叶凋落期;而在展叶期则表现为负相关(图4b3)。五角枫叶性状之间相关关系的斜率在不同生长期均差异显著(P<0.001),即表现出不同的异速生长关系(图4c)。其中,比叶面积和叶厚在展叶期和叶稳定期表现为负相关,且回归斜率的绝对值为展叶期大于叶稳定期,在叶凋落期则表现为正相关(图4c4)。

P<0.05代表叶性状间相关关系随生长期不同发生显著变化。叶性状间的显著关系以回归线段表示,若性状间无显著相关关系,则不显示该回归线。P<0.05 means significant difference in trait-trait relationships for different growth periods of three species. The significant relationship among leaf traits are represented by regression segments. If there is no significant relationship between leaf traits, the regression line is not shown.图4 3种槭树叶性状相关关系在不同生长期的变异Fig. 4 Variations for trait-trait relationships between different growth periods of three Acer species
主成分分析的结果表明,不同树种和叶生长期PC1和PC2合计方差贡献率分别为86.2%和92.7%(图5)。叶厚、比叶面积、叶绿素含量和叶干物质含量均与PC1轴有较强的相关性;叶厚和叶绿素含量与PC2轴有较强的相关性,说明4个叶片性状均可作为叶性状分化的指标。
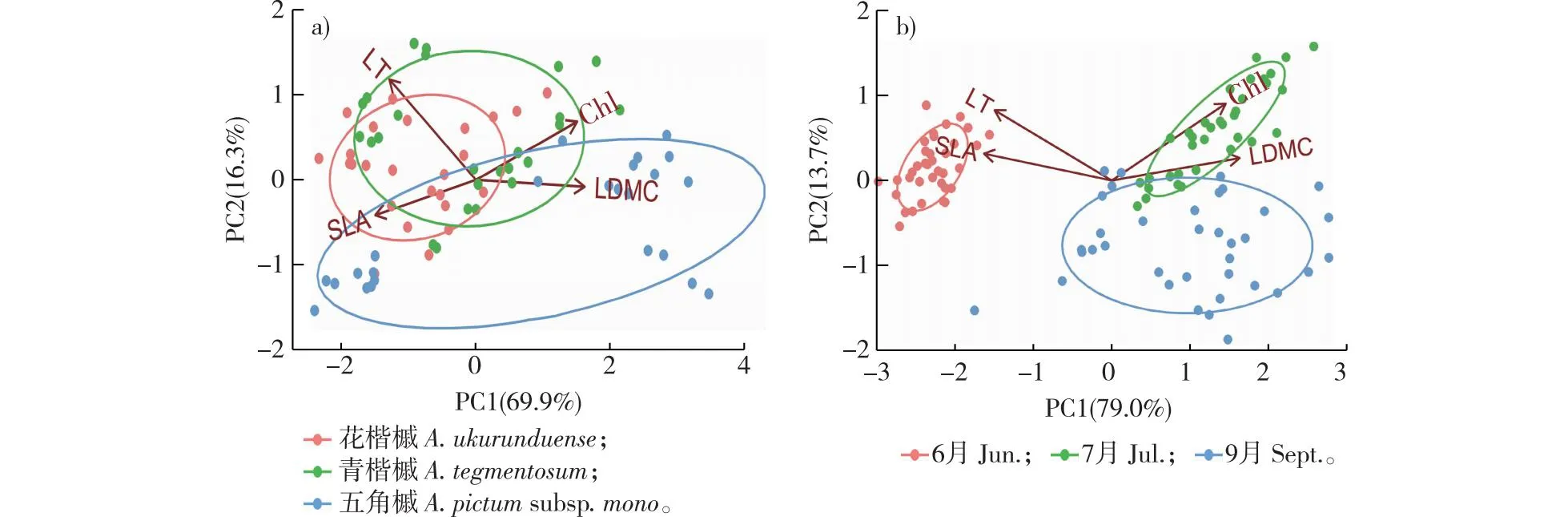
图5 不同树种及叶生长期叶性状的主成分分析Fig. 5 Principal component analysis for leaf traits of different species and leaf growth periods
在树种水平上,不同种槭树的生存策略存在差异。例如花楷槭表现为高比叶面积、低叶干物质含量和叶绿素含量,倾向于选择高生长速率的“快速投资-收益”生存策略(图4a);五角枫则表现为低比叶面积、高叶干物质含量和叶绿素含量,倾向于低生长速率的“缓慢投资-收益”型策略(图5a)。
在叶生长期水平上,展叶期3种槭树有较大的比叶面积和较小的叶干物质含量,表现为高生长速率的“快速投资-收益”型(图5b);而在叶稳定期和叶凋落期表现为较大的叶干物质含量和较小的比叶面积,表现为低生长速率的“缓慢投资-收益”型策略,不同之处在于叶稳定期的叶绿素含量较高(图5b)。
3 讨 论
3.1 叶性状的变异
1)叶性状随生长期的变异。本研究主要分析了3种槭树叶性状在不同叶生长阶段的变异情况,结果显示叶性状随叶生长期变化均存在显著差异,但对于同一叶性状,3种槭树的变异情况又存在一定相似性,如3种槭树的叶厚随叶生长发育均呈减小趋势,叶干物质含量则均先增加再减小。Basnett等[31]认为某些近缘物种由于受到遗传因素和分化时间短的限制会表现出相似的性状特征;Albert等[32]发现随叶片生长,叶绿素含量显著增加。这与本研究结果相符,即3种槭树的叶绿素含量从6月(展叶期)到7月(叶稳定期)均呈显著增加趋势,有利于光合作用的进行。相对而言,到了叶凋落期,由于养分再活化过程的进行,气温降低和叶片衰老等原因,导致叶绿素含量迅速下降[16]。7月展叶过程基本完成,叶片受到来自其他林木叶片的遮盖变多而获得的光照减少,相较在光照充足条件下叶片会变薄[33-34]。这也说明了3种槭树的叶厚在叶稳定期均显著降低。在本研究中的展叶期,即6月初,各树种的比叶面积较大。因此这个时期养料和叶片干物质投资的回报时间较短,叶片生长速率快,有利于展叶的进行[35]。而在炎热干燥的条件下,植物为减少水分散失更倾向于降低比叶面积[18, 36],所以青楷槭和五角枫在7月的比叶面积显著减小。花楷槭和五角枫的比叶面积在9月(叶凋落期)进一步显著降低,可能因为这个时期叶片进入衰老阶段,水分散失加快,使比叶面积减小[10, 37]。展叶期的叶干物质含量比较低,这是因为幼嫩的叶片含水量更高,能够满足展叶阶段各种旺盛生理活动对水分的需求[38]。相反,叶干物质含量高的叶片能有效防止其内部的水分扩散至叶片表面,对水分的保留能力增强[39-40];所以3种槭树在7月(叶稳定期)的叶干物质含量显著增大。
2)树种间的叶性状差异。在树种水平上,较大的比叶面积往往与叶片更强的捕获光照能力有关,而花楷槭和五角枫的比叶面积均显著高于青楷槭,因此在生长环境情况相似的条件下,青楷槭叶片对光照的获取能力可能弱于其余两种槭树。3种槭树的叶厚也存在显著差异,花楷槭的叶厚显著高于青楷槭和五角槭,其叶片有更高的物理强度,对强光的适应能力也增强[12-13, 41]。
3.2 叶性状间相关性在不同生长期的变化
在自然条件下生长的植物能利用的资源是有限的,因此各叶性状间存在普遍的权衡关系[1,6, 43]。本研究中各叶性状之间均存在显著或极显著的正相关或负相关关系,如比叶面积和叶干物质含量间存在极显著负相关关系,这与Wilson等[44]关于英国769个物种的叶性状相关关系的研究结果一致。这主要源于拥有低比叶面积、高叶干物质含量的植物更容易度过资源匮乏及高温等不良环境[10, 12]。本研究中比叶面积和叶厚呈显著正相关关系,这与Qi等[45]的研究结果相反。而田俊霞等[46]关于东北温带针阔混交林的研究表明,叶片通过减小比叶面积和增加叶厚来适应强光和干旱的条件。但本研究中3种槭树在叶稳定期的比叶面积和叶厚均显著降低,这充分体现出不同种植物为适应环境变化所选择的生存策略存在较大差异。
朱弘等[47]的研究表明叶性状的分化会受到遗传和环境因素的共同影响。3种槭树生长在近乎相同的环境中,因此不同种槭树遗传信息的区别可能是决定叶性状分化的主要原因,遗传信息的不同使基因表达产生差异,最终导致不同种槭树叶性状的协作关系产生差异,即花楷槭主要倾向于“快速投资-收益”型生存策略而五角枫则偏向“缓慢投资-收益”型策略。不同叶生长期叶性状分化的原因则可能是遗传信息的不同和各生长期环境条件的差异所共同决定的。在展叶期,叶片对光照的需求量大,因此它们需要在短时间内形成足够的叶面积以获取更多光照[48],大部分能量被用于叶片生长,采取“快速投资-收益”型策略。到叶稳定期,此时叶片相对生长速率很小,叶面积基本无变化,因此在巩固叶片结构上的投资会增多,以便它们更好地抵抗环境压力(高温、风、冰雹等),抵抗病虫害的能力也进一步增强[12, 49],最终达到延长叶片寿命的目的。具体表现为叶干物质含量、叶绿素含量显著增加、比叶面积显著降低,此时植物采取“缓慢投资-收益”型策略。而在9月气温开始下降,植物会选择相对保守的生存策略[1, 50],因此和7月一样也倾向于“缓慢投资-收益”型策略。
Savage[51]证明了不同季节叶性状间的相关关系会发生显著变化,与本研究结果相符。本研究中青楷槭和五角枫的叶干物质含量和叶厚之间的关系在3个生长期均不显著。造成这种现象的原因一方面在于两者受到不同的机制驱动,如叶厚与叶肉细胞的大小以及层数有关,并很大程度上受光照强度的影响[13, 33];叶干物质含量则受到植物所处生境水分的限制,因此受到气孔导度等叶片水力特性的影响更多[41-42],这种相对独立的关系有助于叶片更好地根据环境变化调整其功能。另一方面在于叶片生长时期变化的时间相对较短,叶性状能够变化的范围比较窄[22]。青楷槭的叶干物质含量与叶绿素含量在6月表现为负相关,而在7月和9月表现为正相关,且7月二者相关关系的斜率更大。这可能因为对青楷槭而言,它的叶片在展叶期的气孔导度小,蒸腾所散失的水分减少,叶干物质含量保持在较低水平[16, 23]。同时叶绿素却处于不断合成的过程,两者表现为负相关。叶片在7月通过增加叶绿素含量来维持因自身受其他叶片遮盖所导致的受光面积减小而减弱的光合能力[33],此时叶片基本稳定,相对生长速率较小,且为减少水分散失而保持较高的叶干物质含量[12, 39],两者呈正相关关系;而在9月,叶片处于即将凋落的时期,植物会减少对叶片的投入以提高资源利用率,叶片中的养分也开始向其他组织转移[52]。因此叶片不需要维持较高的叶干物质含量,这可能是7月变异速率大于9月的主要原因。Martinez等[48]认为叶片可以通过适应可利用光的变化而做出性状调整,即叶片的可塑性,它扩大了植物的生态位宽度,提高了它们的生存能力。不同叶生长期,植物经历了不同的环境和自然选择压力,这可以解释在不同生长期各叶性状分配模式的改变[7, 53]。例如对于青楷槭和五角枫,虽然它们的比叶面积和叶干物质含量在3个生长期均表现为显著负相关,但相关关系的斜率在不同生长期之间存在显著差异。
总之,本研究对不同生长期小兴安岭3种槭树叶性状变异和相关性变化的分析结果表明,随生长期变化,3种槭树的叶性状均发生了显著变化。与此同时,每种槭树叶性状间的相关关系随生长期不同均发生了不同程度的显著变化。叶性状及其相关关系在树种水平上也普遍存在显著差异。植物在不同生长期可通过叶性状变异和调整叶性状间的相关关系从而形成最适合自身的生存策略。这些研究结果有助于解析落叶植物的叶生活史策略。
猜你喜欢
中国药学药品知识仓库(2022年10期)2022-05-29
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
现代园艺(2020年12期)2020-12-20
新疆农业科学(2020年1期)2020-02-14
现代园艺(2018年13期)2018-07-11
大众摄影(2018年6期)2018-06-19
中国麻业科学(2015年5期)2015-12-28
卫生职业教育(2014年8期)2014-02-16
绿色科技(2013年8期)2013-08-15
中医研究(2013年9期)2013-03-11
