
锡金尾褐蚬蝶Abisara chela线粒体基因组特征及系统发育分析
2024-01-22 05:05:18刘晨阳夏梦玉杜红雨刘晓光
西南农业学报 2023年11期
刘晨阳,夏梦玉,翟 卿,赵 卓,李 祥,杜红雨,刘晓光
(河南省害虫绿色防控国际联合实验室/河南省害虫生物防控工程实验室/河南农业大学植物保护学院,郑州 450046)
【研究意义】蚬蝶是一种小型蝴蝶,全球已知1400多种,其中93%分布在新大陆。蚬蝶形态上具有突出的模仿性、生殖器相似度高,仅从形态特征和生活习性无法确定锡金尾褐蚬蝶的分类地位。昆虫线粒体基因组全序列长度为15~18 kb,包括13个蛋白质编码基因、22个tRNA基因、2个rRNA基因以及1段控制区[1]。昆虫线粒体基因组母系遗传、结构简单、分子量小、易于检测,因此,目前被广泛应用于分子鉴定、DNA条形码、系统发育等研究,且13个蛋白质编码基因已被证实可用于解决昆虫高级阶元系统发育关系分析[2-6]。【前人研究进展】闫振天等[7]和石庆会等[8]认为应将蚬蝶作为科级单位。有研究认为蚬蝶科下分优蚬蝶亚科(Euselasiinae)、古蚬蝶亚科(Nemeobiinae)和蚬蝶亚科(Riodininae)3个亚科[9-11]。其中古蚬蝶亚科分为褐蚬蝶族(Abisarini),包括褐蚬蝶属(Abisara)、莱蚬蝶属(Laxita)、暗蚬蝶属(Paralaxita)和白蚬蝶属(Stiboges)4属;古蚬蝶族(Nemeobiinae),包括红斑蚬蝶属(Hamearis)、沙蚬蝶属(Saribia)、小蚬蝶属(Polycaena)、豹蚬蝶属(Takashia)和藏蚬蝶属(Tibetododona)5属;波蚬蝶族(Zemerini),包括波蚬蝶属(Zemeros)、尾蚬蝶属(Dodona)2属;以及未明确划分族的塔蚬蝶属(Taxila)、迪蚬蝶属(Dicallaneura)和前列蚬蝶属(Praetaxila)3属。近年系统发育分析发现,古蚬蝶亚科内分类系统与传统分类系统存在巨大差异。Espeland等[12]认为应将古蚬蝶亚科分为褐蚬蝶族和古蚬蝶族,将传统的波蚬蝶族拆分为波蚬蝶亚族(Zemeros)和尾蚬蝶亚族(Dodonaina)并入古蚬蝶族;原属于优蚬蝶亚科中的白珂蚬蝶属(Corrachia)和粉蚬蝶属(Styx)整合为白珂蚬蝶亚族(Corrachina)归入古蚬蝶族;原属于褐蚬蝶族中的黄带褐蚬蝶(Abisarafylla)和白带褐蚬蝶(Abisarafylloides)与古蚬蝶族中的白蚬蝶(Stibogesnymphidia)组成白蚬蝶亚族(Stibogina);将塔蚬蝶属、迪蚬蝶属、前列蚬蝶属归入褐蚬蝶族。目前蝶类较为通用的5科分类系统中,蚬蝶作为灰蝶科下的亚科。【本研究切入点】目前褐蚬蝶族内的分类系统混乱,锡金尾褐蚬蝶(A.chela)归属存在争议,尚无明确定位。【拟解决的关键问题】本文测定并分析锡金尾褐蚬蝶线粒体基因组特征,探究蚬蝶与灰蝶的系统进化关系以及锡金尾褐蚬蝶的分类地位。
1 材料与方法
1.1 实验材料
本研究所采用的7种蚬蝶标本均于2021年7—8月在西藏自治区采集,种类鉴定依据形态特征。具体采集信息如表1所示。
1.2 基因组DNA的提取与测定
试剂盒为TIANamp Fenomic DNA Kit血液/细胞/组织基因组DNA提取试剂盒(离心柱型),由天根生化科技(北京)有限公司提供。挑取-20 ℃冻存标本胸部肌肉,将标本剩余部分标注对应编号保存,按照试剂盒标注步骤提取总DNA。测定浓度,选取高浓度样本完成高通量测序,测序工作由北京贝瑞和康生物技术有限公司完成。
1.3 测定序列的拼接、分析与注释
将测序所得短片段拼接,选择长度在15 000 bp左右的序列,删除重复序列,上传至Mitos网站(http://mitos.bioinf.uni-leipzig.de/index.py)注释。依据注释结果,使用MEGA 6软件,对13个蛋白质编码基因、2个rRNA基因和A+T富含区与其他已测定鳞翅目昆虫比对后,调整起始和终止位置后上传NCBI网站获取GenBank登录号。
利用MEGA 6软件分析13个蛋白质编码基因序列、主要编码链J链上的tRNA基因和次要编码链N链上的tRNA基因的核苷酸组成,使用AT-skew=(A-T)/(A+T)和GC-skew=(G-C)/(G+C)公式计算AT偏斜和GC偏斜,分析13个蛋白质编码基因的氨基酸使用频率和相对同义密码子使用频率(RSCU)。根据上传Mitos网站的注释结果绘制22个tRNA基因的二级结构,并利用OGDraw网站(https://chlorobox.mpimp-golm.mpg.de/OGDraw.html)在线绘制线粒体基因组结构。
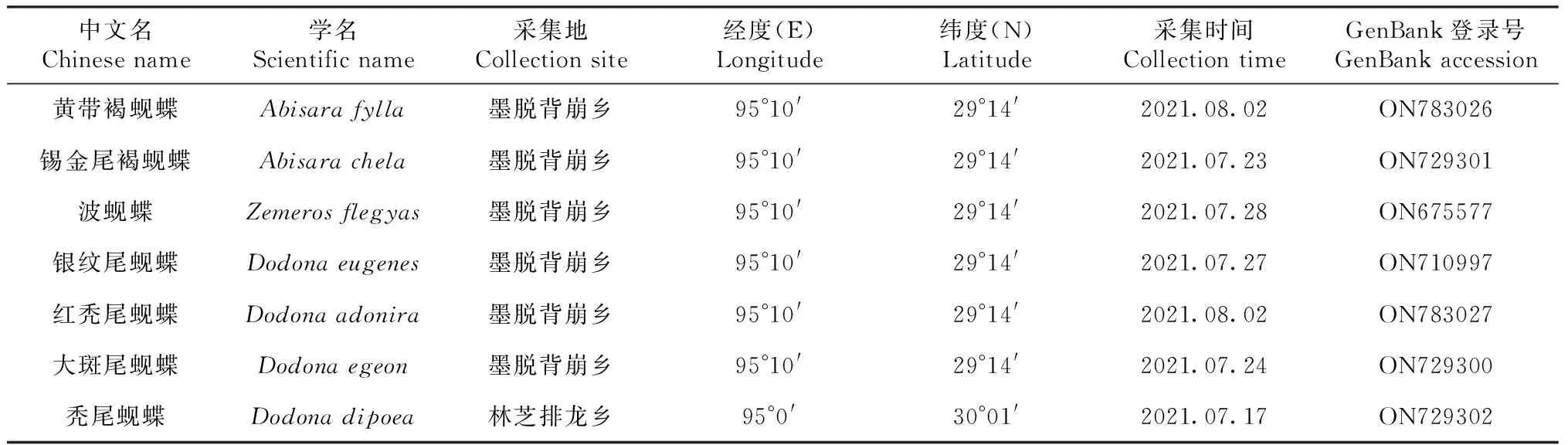
表1 标本采集信息
1.4 系统发育分析
选取51种蝴蝶的线粒体全基因组,其中7种蚬蝶自测,44种由NCBI下载,以家蚕(Bombyxmori)和美国白蛾(Hyphantriacunea)作为外群,分别采用最大似然法(Maximum Likelihood,ML)和贝叶斯法(Bayesian inference,BI)构建系统发育树。使用Phylosuite软件中的ModelFinder,筛选出贝叶斯推断最佳核苷酸替换模型GTR+I+F+G4,设置贝叶斯推断中运行4条马尔科夫链蒙特卡洛算法(Markov Chain Monte Carlo,MCMC),运行代数为2000 000代,每100代取一次样,舍去25%的老化样本;筛选出的最大似然法推断的最佳替换模型为GTR+F+R5,系统发育树各分支节点的置信水平选择ultrafast bootstrap,重复抽样10 000次。
2 结果与分析
2.1 锡金尾褐蚬蝶线粒体基因组结构
锡金尾褐蚬蝶线粒体基因组全序列总长度为15 259 bp,由37种基因组成,包括13个蛋白质编码基因、22个tRNA基因、2个rRNA基因和1个A+T富含区,呈闭合双链环状结构。37个基因中有位于J链上的9个蛋白质编码基因(nad2、cox1、cox2、atp8、atp6、cox3、nad3、nad6、cob)和14个tRNA基因(trnM、trnI、trnW、trnL2、trnK、trnD、trnG、trnA、trnR、trnN、trnS1、trnE、trnT、trnS2),N链上有4个蛋白质编码基因(nad5、nad4、nad4l、nad1)和8个tRNA基因(trnQ、trnC、trnY、trnF、trnH、trnP、trnL1、trnV)(图1)。锡金尾褐蚬蝶线粒体基因组中有8处基因重叠,共31 bp,重叠最长的8 bp在trnW和trnC之间,重叠最短的1 bp在nad4和nad4l之间;基因间隔有13处,共146 bp,间隔最长的58 bp在trnQ和nad2之间,间隔最短的1 bp在trnL2和cox2之间;无间隔无重复的16处(表2)。
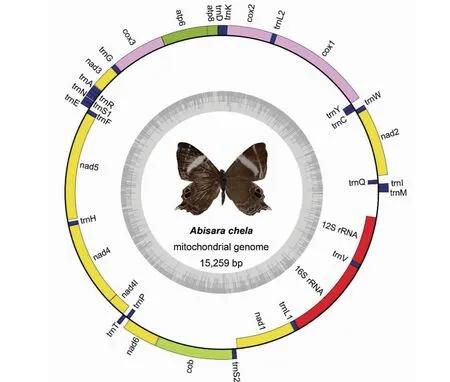
图1 锡金尾褐蚬蝶线粒体基因组结构Fig.1 Structure of the mitochondrial genome of A. chela
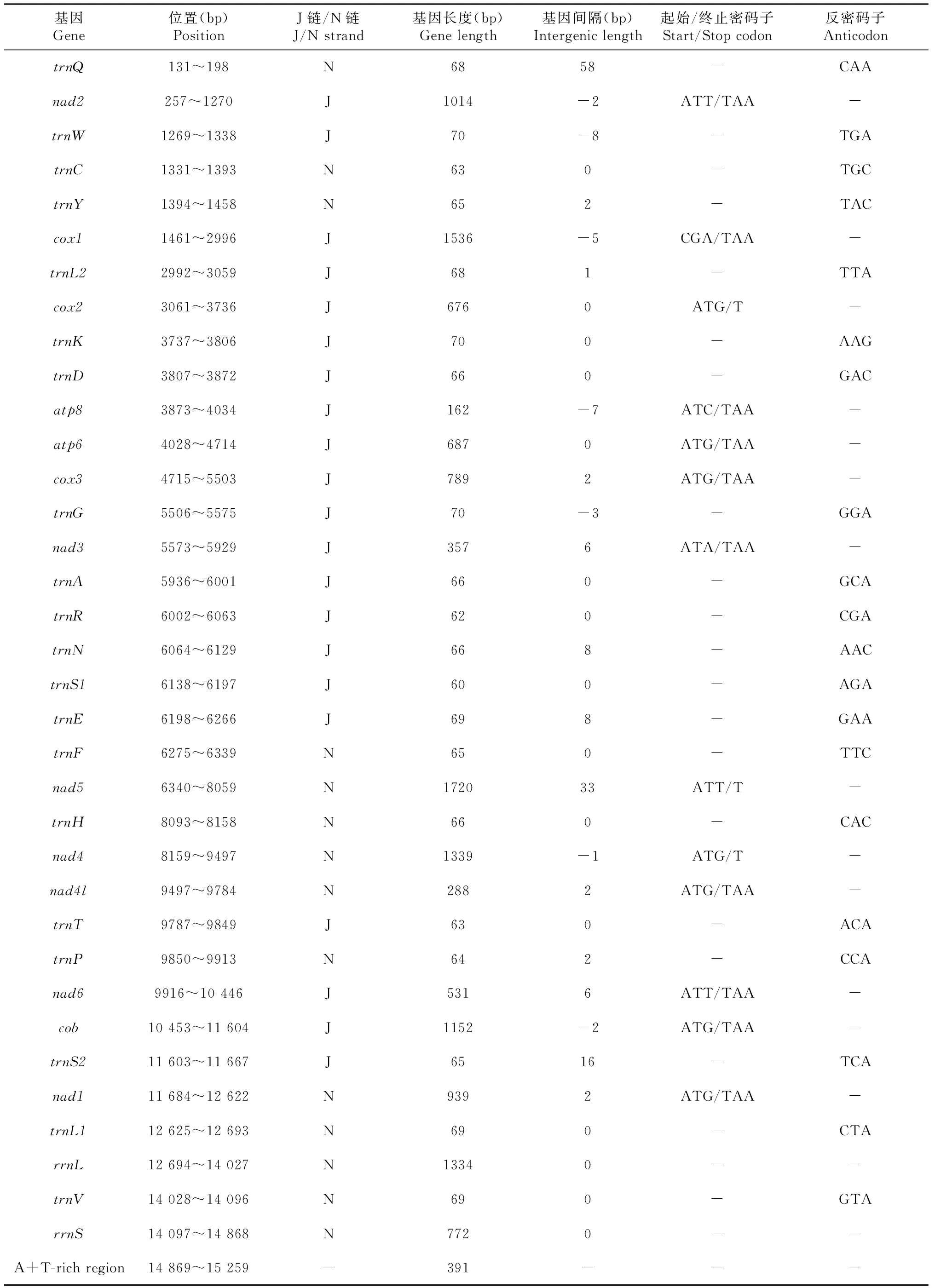
续表2 Continued table 2
2.2 核苷酸组成
锡金尾褐蚬蝶线粒体全基因组中,A、T、G和C含量分别为39.9%、40.6%、7.8%和11.7%,A+T含量为80.5%,G+C含量为19.5%,有明显的AT偏向性。AT偏斜(AT-skew)和GC偏斜(GC-skew)分别为-0.009和-0.200,表明线粒体全基因组中碱基含量T高于A,C高于G(表3)。
2.3 蛋白质编码基因分析
锡金尾褐蚬蝶线粒体基因组中蛋白质编码基因序列总长为11 182 bp,约占线粒体基因组全序列的73.28%。其中nad5基因最长,为1720 bp;atp8基因最短,为162 bp。其A+T含量为79.0%,G+C含量为21.0%,AT偏斜和GC偏斜分别为-0.137和-0.038。
13个蛋白质编码基因中,除cox1以CGA为起始外,其它均以标准密码子(ATN)为起始,其中atp8的起始密码子为ATC,nad3的起始密码子为ATA,nad2、nad5和nad6的起始密码子均为ATT,cox2、atp6、cox3、nad4、nad4l、cob和nad1的起始密码子则为ATG;除cox2、nad5和nad4以T为终止,其余的终止密码子均为TAA(表2)。
共有60组不同的密码子,其中使用频率较高的依次为UUA(Leu,467次)、AUU(Ile,440次)、UUU(Phe,332次)、AUA(Met,259次)和AAU(Asn,252次),均完全由A或U构成(图2-A)。以A/U结尾的氨基酸密码子的使用频率频繁于以G/C结尾的。使用频率较为频繁的氨基酸依次为Leu(552次)、Ile(469次)、Phe(355次)、Ser(348次)和Met(301次),使用频率较低的氨基酸依次为His(65次)、Gln(61次)、Asp(59次)、Arg(54次)和Cys(27次)。
UUA的相对同义密码子使用频率最高,RSCU值为5.08。Ser和Leu的RSCU值分别为8.01和6.01,Val、Pro、Thr、Ala、Arg和Gly的RSCU值均为4,其它12个氨基酸的RSCU值均为2(图2-B)。
2.4 tRNA基因分析
基因组注释结果显示,22个tRNA基因长度均在60~70 bp。J链上的tRNA基因序列A+T含量为79.8%,G+C含量为20.2%,AT偏斜和GC偏斜分别为-0.010和0.426;N链上的tRNA基因序列A+T含量为82.1%,G+C含量为17.9%,AT偏斜和GC偏斜分别为0.028和0.017。
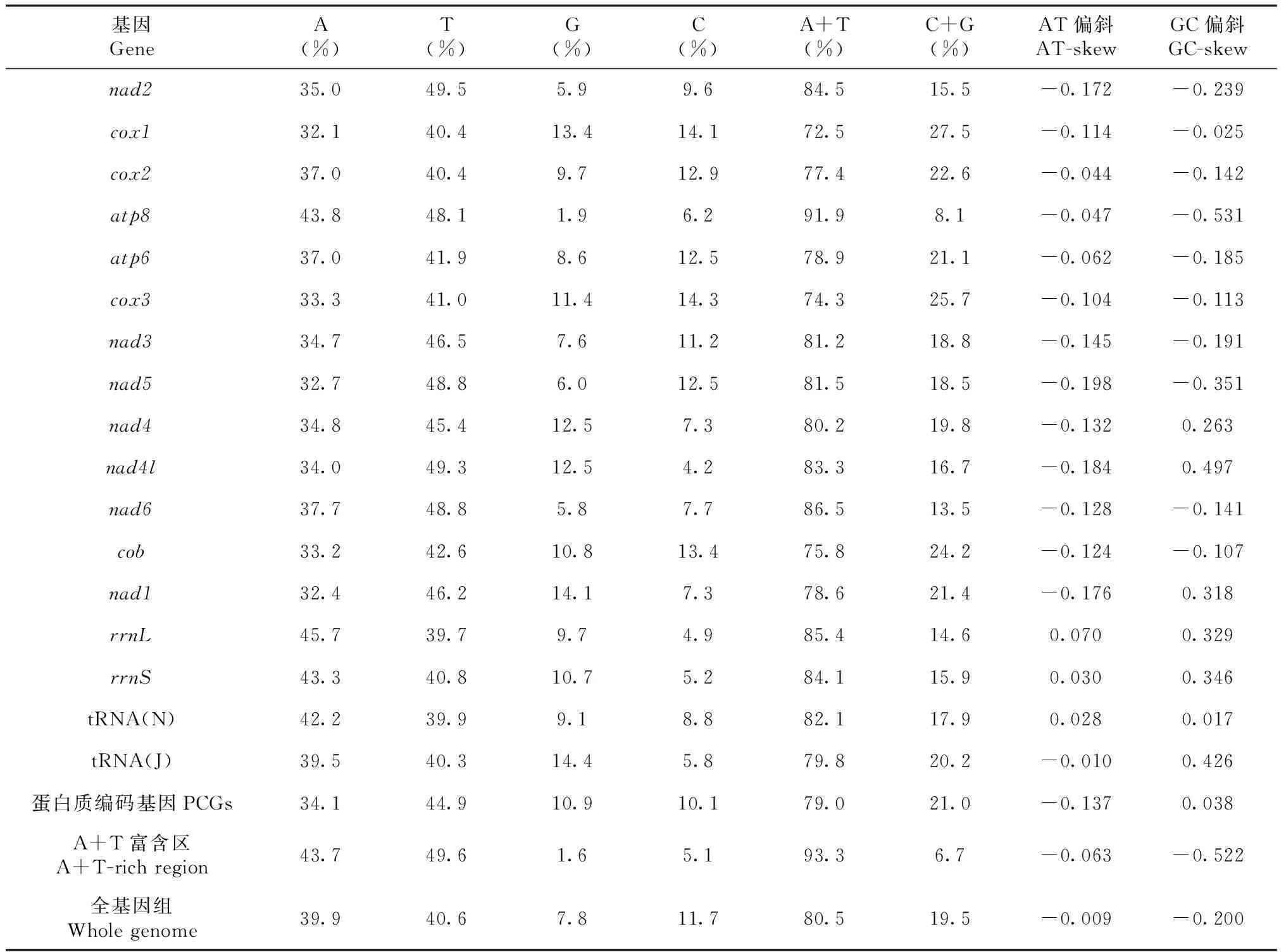
表3 锡金尾褐蚬蝶线粒体基因组核苷酸组成
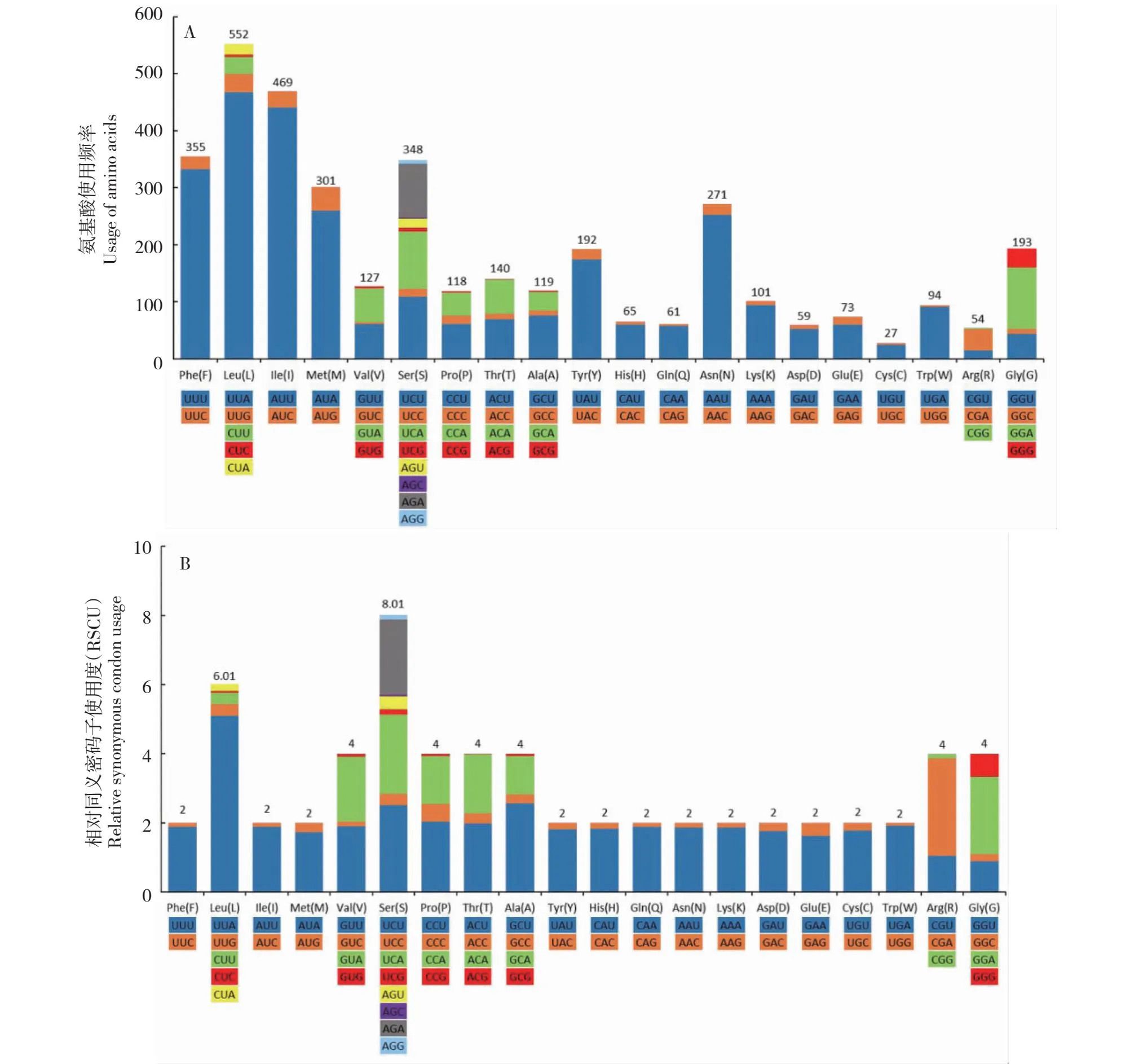
图2 锡金尾褐蚬蝶蛋白质编码基因的氨基酸使用频率(A)和相对同义密码子使用度(RSCU)(B)Fig.2 Usage of amino acids (A) and the relative synonymous codon usage (B) of protein-coding genes of mitochondrial genome in A. chela
除trnS1基因缺少DHU臂外,锡金尾褐蚬蝶的其他21个tRNA基因的二级结构均为典型的三叶草型,与目前已知鳞翅目昆虫的tRNA基因二级结构一致[19-21]。有19对G=U错配分别出现于trnA、trnC、trnF、trnG、trnH、trnI、trnL1、trnP、trnQ、trnT、trnV、trnW基因的氨基酸接受臂、TψC臂、二氢尿嘧啶臂和反密码子臂;4对U=U错配出现在trnA、trnL2、trnS2基因的氨基酸接受臂和反密码子臂上(图3)。
2.5 rRNA基因分析
锡金尾褐蚬蝶线粒体基因组中有2个rRNA基因,rrnL(1334 bp)基因位于trnL1和trnV之间,rrnS(772 bp)基因位于trnV和A+T富含区之间,都有较高的A+T含量,分别为85.4%和84.1%。
2.6 A+T富含区分析
A+T富含区又称为控制区,是昆虫线粒体中的非编码序列,锡金尾褐蚬蝶线粒体基因组中的A+T富含区位于rrnS和trnM之间,长度为391 bp,其A+T含量高达93.3%,明显高于线粒体基因组中其它部分A+T含量。
锡金尾褐蚬蝶的A+T富含区端部存在一个由保守序列“ATAGA”引导的19 bp的多聚T结构(图4);中部存在2个6 bp的串联重复序列(TATATT)、2个8 bp的串联重复序列(TTAATTAT)和1个微卫星片段(TA)6;尾部存在一个10 bp的多聚A结构。
2.7 系统发育分析
基于线粒体基因组13个蛋白质编码基因和2个rRNA基因的串联序列,以家蚕(B.mori)和美国白蛾(H.cunea)作为外群分别采用贝叶斯法(BI)和最大似然法(ML),分析51种蝴蝶的系统发育关系,构建BI树(图5)和ML树(图6)。结果表明,2种系统发育分析方法得到相似的树拓扑结构,均显示建树所用蝶类分为5个大分支:蚬蝶与灰蝶聚为一支,BI后验概率(BPP)和ML自举值(MLBP)分别为1、100;蛱蝶、珍蝶、眼蝶、环蝶、斑蝶和喙蝶聚为一支(BBP=1,MLBP=100);粉蝶单独为一支(BPP=1,MLBP=100);弄蝶单独为一支(BPP=1,MLBP=100);凤蝶和绢蝶聚为一支(BPP=1,MLBP=100)。其中蚬蝶、灰蝶、眼蝶、斑蝶、粉蝶和弄蝶为单系群;蚬蝶与灰蝶互为姐妹群(BPP=1,MLBP=100)。黄带褐蚬蝶(A.fylla)和白带褐蚬蝶(A.fylloides)聚为一支(BPP=1,MLBP=100),但锡金尾褐蚬蝶(A.chela)独立于褐蚬蝶属(Abisara)之外(BPP=1,MLBP=100)。相较于其它类群,眼蝶和环蝶的亲缘关系更近(BPP=1,MLBP=100)。
BI树显示,本研究中各类群间关系为蚬蝶种间关系为(A.chela+((A.fylloides+A.fylla)+(Z.flegyas+(D.dipoea+(D.adonira+(D.eugenes+(D.maculosa+D.egeon))))))),其中波蚬蝶属(Zemeros)和尾蚬蝶属(Dodona)聚为一支(BBP=0.54)。苎麻珍蝶(Acraeaissoria)先与蛱蝶中的斐豹蛱蝶(Argynnishyperbius)和嘉翠蛱蝶(Euthaliakardama)聚为一支(BPP=1),然后与大紫蛱蝶(Sasakiacharonda)和柳紫闪蛱蝶(Apaturailia)的集合互为姐妹群(BPP=1)。

图4 锡金尾褐蚬蝶线粒体基因组A+T富含区结构Fig.4 Structure of A + T-rich region of A. chela mitochondrial genome
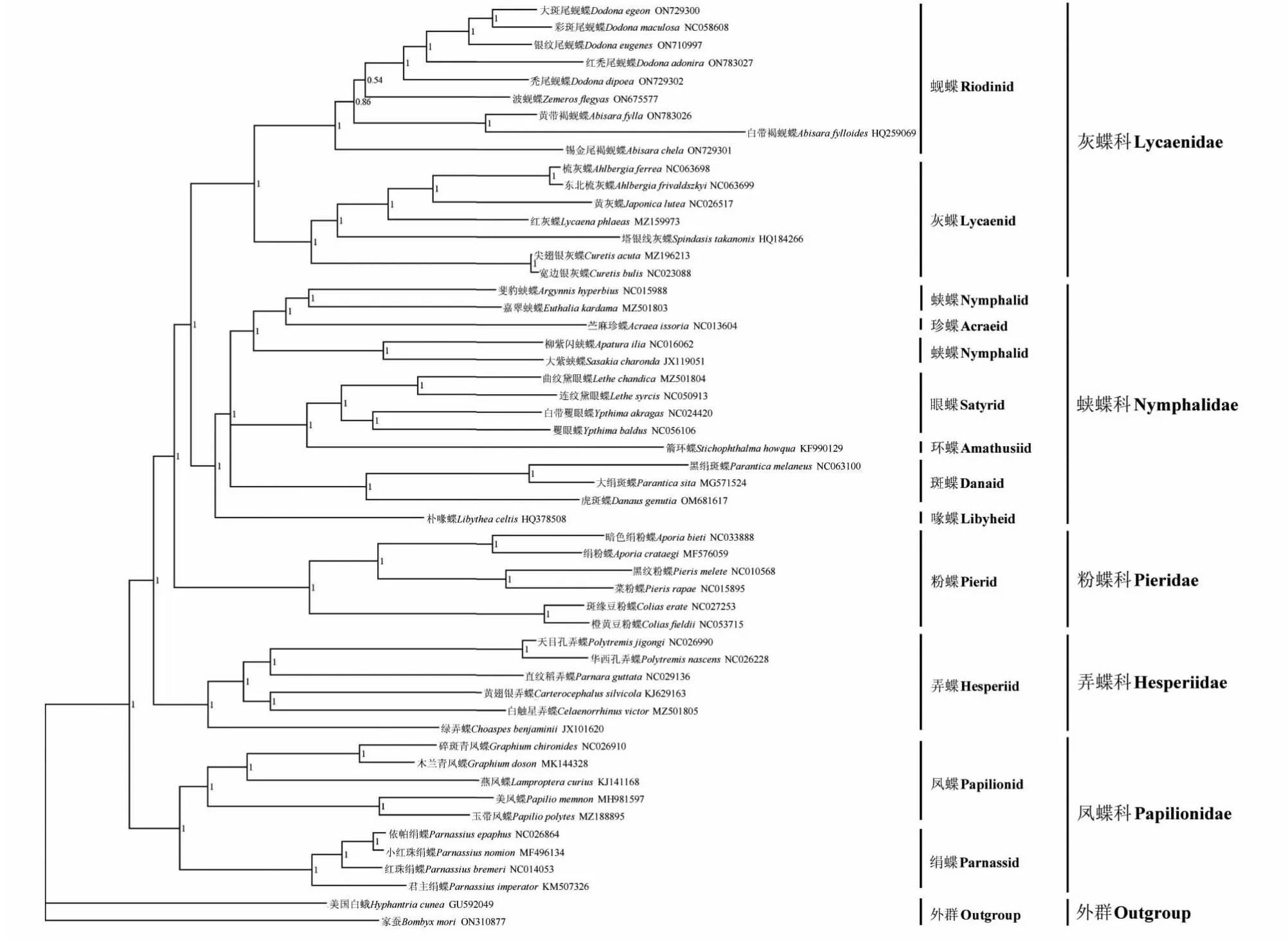
图5 基于13个蛋白质编码基因和2个rRNA基因序列构建的蝶类贝叶斯法系统发育树Fig.5 Bayesian phylogenetic tree of butterflies based on the sequences of 13 protein-coding genes and 2 rRNA genes of mitochondrial genome

图6 基于13个蛋白质编码基因和2个rRNA基因序列构建的蝶类最大似然法系统发育树Fig.6 Maximum likelihood phylogenetic tree of butterflies based on the sequences of 13 protein-coding genes and 2 rRNA genes of mitochondrial genome
ML树显示,蝴蝶类群间关系为((Papilionid+Parnassiid)+(Hesperiid+(Pierid+((Libyheid+(Danaid+((Amathusiid+Satyrid)+(Acraeid+Nymphalid))))+(Lycaenid+Riodinid)))))。蚬蝶种间关系为(A.chela+((Z.flegyas+(A.fylloides+A.fylla))+(D.dipoea+(D.adonira+(D.eugenes+(D.maculosa+D.egeon)))))),其中黄带褐蚬蝶、白带褐蚬蝶和波蚬蝶(Z.flegyas)聚为一支(MLBP=77),然后与尾蚬蝶属互为姐妹群(MLBP=88%)。苎麻珍蝶先与蛱蝶中的斐豹蛱蝶聚在一起(MLBP=69),然后与嘉翠蛱蝶聚为一支(MLBP=100),之后与大紫蛱蝶和柳紫闪蛱蝶互为姐妹群(MLBP=99)。
3 讨 论
3.1 线粒体基因组特征分析
锡金尾褐蚬蝶线粒体基因组中,37个基因包括2个核糖体基因(rrnL和rrnS)、22个转运RNA基因和13个蛋白质编码基因。13个蛋白质编码基因包括1个细胞色素b(cob)、2个ATP酶亚基基因(atp6和atp8)、3个细胞色素氧化酶亚基基因(cox1、cox2和cox3)、7个NADH降解酶基因(nad1、nad2、nad3、nad4、nad5、nad6和nad4l),cox1基因以CGA为起始,其余均以ATN为起始;cox2、nad5和nad4基因以T为终止,其余终止密码子均为TAA。锡金尾褐蚬蝶线粒体基因的排列顺序和编码方向与已知大多数鳞翅目昆虫一致[13-17]。
锡金尾褐蚬蝶线粒体基因全序列具有AT偏向性,符合昆虫线粒体基因中含有较高的A+T含量的特征[18]。tRNA基因二级结构中,除trnS1基因缺少DHU臂外,均为标准的三叶草结构,与鳞翅目中已测定的线粒体基因组tRNA基因表现一致[19-21]。22个tRNA基因二级结构中有19对碱基错配。A+T富含区中的多聚T结构,目前已被证实为复制起点[22-24]。
3.2 系统关系分析
蚬蝶与灰蝶各为单系群,互为姐妹群,支持Cong等[25]将蚬蝶和作为亚科列入灰蝶科的观点。珍蝶、蛱蝶、眼蝶、环蝶、斑蝶和喙蝶归入蛱蝶科,其中眼蝶与环蝶的亲缘关系相对于其它类群较近。粉蝶和弄蝶均为单系群,认可粉蝶和弄蝶分别为科级单元的观点。凤蝶和绢蝶并列为凤蝶科的亚科。波蚬蝶属(Zemeros)和尾蚬蝶属(Dodona)的亲缘关系相较于褐蚬蝶属(Abisara)较近,这一点与Campbell等[11]观点一致,但是黄带褐蚬蝶(A.fylla)和白带褐蚬蝶(A.fylloides)未与锡金尾褐蚬蝶(A.chela)聚为一支,而是与波蚬蝶属和尾蚬蝶属亲缘关系更近。依据2种系统发育树结果,认可Espeland等[12]将黄带褐蚬蝶和白带褐蚬蝶归入古蚬蝶族(Nemeobiinae)的观点;锡金尾褐蚬蝶仍属于褐蚬蝶族(Abisarini),与比较形态学分类地位一致。黄带褐蚬蝶、白带褐蚬蝶和锡金尾褐蚬蝶的学名厘定还需要另做研究。
2种系统发育树在波蚬蝶属和珍蝶属(Acraea)存在差异。BI树中,波蚬蝶属和尾蚬蝶属聚为一支,然后与黄带褐蚬蝶和白带褐蚬蝶互为姐妹群。ML树中,黄带褐蚬蝶、白带褐蚬蝶和波蚬蝶属聚为一支,然后与尾蚬蝶属互为姐妹群。2种系统发育树中波蚬蝶属处的节点支持率分别为0.54%、77.00%,需要增加样本并与其它特征联合分析才能确定其准确系统发育关系。BI树中,蛱蝶中的斐豹蛱蝶(Argynnishyperbius)和嘉翠蛱蝶(Euthaliakardama)的亲缘关系相较于苎麻珍蝶(Acraeaissoria)更近。ML树中,苎麻珍蝶和蛱蝶中的斐豹蛱蝶的亲缘关系相较于嘉翠蛱蝶更近。虽然斐豹蛱蝶、嘉翠蛱蝶和苎麻珍蝶在2种系统发育树的结果不同,但聚在一起后均与大紫蛱蝶(Sasakiacharonda)和柳紫闪蛱蝶(Apaturailia)互为姐妹群。本研究认为,珍蝶应作为蛱蝶科下的族或属,归入蛱蝶亚科,这与邹方振等[26]认为应将珍蝶作为亚科归入蛱蝶科的观点不一致,还需进一步研究确认。
古蚬蝶族中分类系统研究较多,目前较为通用的是5亚族系统:古蚬蝶亚族(Nemeobiina)、白蚬蝶亚族(Stibogina)、波蚬蝶亚族(Zemerina)、尾蚬蝶亚族(Dodonaina)和白珂蚬蝶亚族(Corrachina),但褐蚬蝶族下阶元分类尚无定论。由于已公布古蚬蝶亚科昆虫线粒体基因组数据数量较少,尚不能确定锡金尾褐蚬蝶在褐蚬蝶族中的位置,因此,无论是比较形态学还是基因组学,在后续研究中都有必要增加研究种类。
4 结 论
锡金尾褐蚬蝶线粒体基因组结构特征符合鳞翅目和蝶类的基本规律;基于线粒体基因组特征构建的贝叶斯树和最大似然树均支持目前较为通用的蝴蝶5科系统;锡金尾褐蚬蝶归入古蚬蝶族,黄带褐蚬蝶和白带褐蚬蝶保留古蚬蝶族的地位,锡金尾褐蚬蝶的地位在获得更多证据支撑前,先暂时保留在古蚬蝶族。
猜你喜欢
幼儿100(2020年25期)2020-10-22 05:25:20
武夷科学(2019年2期)2019-12-20 08:39:16
小学生必读(低年级版)(2018年4期)2018-08-01 09:27:54
百姓生活(2018年3期)2018-03-26 08:19:52
人生与伴侣·共同关注(2017年9期)2017-09-13 17:13:49
环球时报(2017-08-08)2017-08-08 12:01:25
小学生必读(低年级版)(2017年11期)2017-03-15 07:53:40
武夷科学(2017年0期)2017-03-10 10:03:31
天然产物研究与开发(2014年8期)2014-04-27 14:16:30
中国青年(1996年4期)1996-08-28 03:47:20
