
云南花椒黄花病相关病原分析
2024-01-22 01:13余烯宇张丽珍郝佳波呼延丽杜春花陈永对董家红
西南农业学报 2023年11期
余烯宇,吴 阔,张丽珍,郝佳波,呼延丽,杜春花,陆 斌,陈永对,董家红
(1.云南中医药大学中药学院/中药材栽培研究所/云南省南药可持续利用重点实验室,昆明 650500;2.云南省农业科学院生物技术与种质资源研究所,昆明 650205;3.云南省林业和草原科学院,昆明 650204)
【研究意义】花椒为芸香科花椒属植物,世界上有250种,中国有45种以及13个变种,是具较高商业价值的药食同源作物,日常多用于风味调料品使用,其在化妆品市场也有较大发展空间,栽培品种有大红袍、汉源花椒、粤西贡椒等[1-2]。药用部位为干燥成熟果皮,花椒常见功效为温中止痛、杀虫止痒[3]。对花椒的功效深入研究发现其所含生物碱、木脂素、香豆素、甾萜类等成分有抗肿瘤、心脑保护、抗血小板凝聚、降压等功效[4]。随着花椒种植规模扩大,多地建立了花椒种植基地,花椒病害成为危害花椒产业的重要因素,花椒叶、根、枝部都极易受不同病虫害影响,如花椒蚜虫、花椒窄吉丁、花椒鞘锈菌、花椒生理性病害等[5]。近年花椒黄花病在四川、重庆、云南等地发生流行,其主要症状为花朵雄蕊泛黄、雌蕊败育,叶片黄化、斑点,果实产量降低[6],利用高通量测序分析花椒黄花病病原,对后续解决花椒黄花病并保证花椒经济价值具有重要意义。【前人研究进展】有研究对产于重庆的九叶青花椒(Zanthoxylumarmatumvar.novemfolius)测序发现,花椒中5种新病毒,分别为伴生豇豆病毒科(Secoviridae)线虫多面体病毒属(Nepovirus)的四川青花椒线虫多面体病毒(Green Sichuan pepper-nepovirus,GSPNeV),梅奥病毒科(Mayoviridae)悬钩子病毒属(Idaeovirus)的四川青花椒悬钩子病毒(Green Sichuanpepper-idaeovirus,GSPIV),黄症病毒科(Luteoviridae)耳突花叶病毒属(Enamovirus)的四川青花椒耳突花叶病毒(Green Sichuan pepper-enamovirus,GSPEV),弹状病毒科(Rhabdoviridae)胞核弹状病毒属(Nucleorhabdovirus)的四川青花椒发现弹状病毒(Green Sichuan pepper-nucleorhabdovirus,GSPNuV),在青花椒(Z.schinifolium)花椰菜花叶病毒科(Caulimoviridae)的四川青花椒脉明相关病毒(Green Sichuan pepper vein clearing-associated virus,GSPVCaV)病毒[7-8]。也有研究发现四川花椒(Z.bungeanuminSichuan)黄花病中有植原体感染[9]。【本研究切入点】为明确云南昭通竹叶花椒永青1号黄花病的病原,本研究用高通量测序技术(High-throughput sequencing,HTS)对4个感病样品与2个无症样品进行宏基因组测序分析,得到花椒病毒序列,对相关病毒进行分析。【拟解决的关键问题】分析花椒宏基因组测序所得的病毒基因组,结合前人的研究分析,筛选与黄花病症关系较为紧密的病毒,为明确花椒黄花病病原提供理论依据。
1 材料与方法
1.1 花椒样品
于2021年7月取自云南昭通花椒基地,品种为竹叶花椒永青1号(Zanthoxylumarmatumvar. yongqing 1)。
1.2 花椒RNA提取及花椒转录组测序分析
取自云南昭通竹叶花椒永青1号花椒种植基地的无病症花椒共2组,标记为H-1、H-2。选择感病花椒4组,标记为D-1、D-2、D-3、D-4,挑选花椒叶片和花部分,分别用Eastep Super总RNA提取试剂盒(动植物两用型,上海普洛麦格生物产品有限公司)提取总RNA,琼脂糖凝胶电泳测试RNA,保证RNA提取质量较好,将提取的RNA送至上海欧亿生物医药科技有限公司使用SPAdes对质控后数据进行组装,最小contigs片段大小设置为50,通过NR注释选取比对上数据库中病毒序列的contigs,使用PRodigal对virus序列进行基因预测。
1.3 病毒序列分析
用Origin Pro 2021对contigs在12种病毒科组内的占比进行可视化聚类分析,将所有NR注释的FYD花椒与健康花椒所有contigs通过NCBI BLAST匹配到不同的病毒序列,用本地注释文件定位不同contigs于病毒的位点,用DNAMAN将匹配到相同序列号的同类型病毒调整为碱基5′→3′的顺序,按匹配点位进行拼接,得到较完整序列,使用MEME(Multiple Em for Motif Elicitation)分析RNA保守位点,通过 ORF预测蛋白开放阅读框,使用CDD(Conserved domains database)分析蛋白结构域。
2 结果与分析
2.1 感黄花病花椒样品中病毒种类分析
在Illumina HiSeqTM 2500平台上对6个样品进行高通量测序获得总长351 139 315 nt,并拼接获得707 122个基因片段,GC平均占比44.42%,平均contigs长度为335.99 nt,保证拼接数据高质量。对6个花椒样品进行病毒NR注释,发现484个contigs可匹配到12个病毒科15种病毒。在6个样品中均检测出在九叶青花椒上已报道的4种病毒:四川青花椒线虫多面体病毒(GSPNeV)、四川青花椒悬钩子病毒(GSPIV)、四川青花椒耳突花叶病毒(GSPEV)、四川花椒弹状病毒(GSPNuV),contigs占比25.82%;同时发现2种未报道的新病毒,分别命名为永青花椒相关幽影病毒(Yongqing green pepper-associated umbravirus)和永青花椒相关负链RNA病毒(Yongqing green pepper-associated ssRNA negative-strand virus),contigs占比2.52%。从感病样品D-2、D-3、D-4发现乙型线形病毒科(Betaflexiviridae)香石竹潜隐病毒属(Carlavirus)马铃薯M病毒(Potato virus M,PVM)、马铃薯H病毒(Potato virus H,PVH)和马铃薯S病毒(Potato virus S,PVS),同时在D-1样品中发现葡萄茎痘相关病毒(Grapevine rupestris stem pitting-associated virus,GRSPAV)。从图1可知,Betaflexiviridae病毒仅存在于显病组,且为独立类别。整理不同测序组样品中已报道的4种新病毒及未报道的2种新病毒及乙型线形病毒科(Betaflexiviridae)的病毒contigs范围,通过NCBI BLAST确定病毒种类(表1)。已被报道的GSPNeV、GSPIV、GSPEV、GSPNuV病毒在每组花椒中均有发现,contigs范围大且氨基酸相似性在86.16%~100%;其中未报道的2种新病毒也能从所有测序花椒样品中检测到。依据NCBI BLAST比对结果以其最相近病毒为参考将2种新病毒初步命名为永青花椒相关幽影病毒(Yongqing green pepper-associated umbravirus)和永青花椒相关负链RNA病毒(Yongqing green pepper-associated ssRNA negative-strand virus),而上述6种病毒contig长度在无症组与感病组无明显差异。通过DNAMAN对病毒序列进行拼接得到较长的永青花椒相关幽影病毒和永青花椒相关负链RNA病毒序列,永青花椒相关幽影病毒序列长2900~3080 nt,永青花椒相关负链RNA病毒序列长7306 nt。通过BLAST发现FYD组花椒病毒39个contigs比对到乙型线形病毒科(表2),其中D-2、D-3、D-4中存在马铃薯M病毒(Potato virus M,PVM)、马铃薯H病毒(Potato virus H,PVH)、马铃薯S病毒(Potato virus S,PVS)3类病毒,此3种病毒均属于乙型线形病毒科(Betaflexiviridae)香石竹潜隐病毒属(Carlavirus)[10],而中D-1组仅有凹陷病毒属(Foveavirus)的葡萄茎痘相关病毒(Grapevine rupestris stem pitting-associated virus)2个短片段contigs。
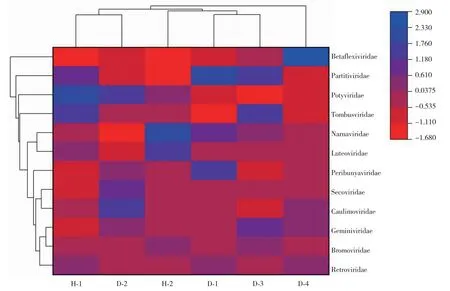
图1 依据12个病毒科contigs占比的热图聚类分析Fig.1 Heatmap cluster analysis based on the percentage of contings in 12 virus families
2.2 永青花椒相关幽影病毒分析
由表1可知,从6个样品得到长度为302~3080 nt的永青花椒相关幽影病毒(Yongqing green pepper-associated umbravirus)contigs,对不同核苷酸序列进行两两比对发现相似性为88.1%~100.0%, 其中H-2、D-1、D-3、D-4分别得到长为3025、3022、2900、3080 nt的病毒序列,4条序列的相似性在92.1%~97.8%,通过NCBI氨基酸BLAST发现与幽影病毒属(Umbravirus)的仙人掌幽影病毒(Opuntia umbra-like virus, GenBank:AXG65483.1)最相似,覆盖率为50%~53%,相似性为49.90%~50.56%,核酸序列相似性为51.9%~52.1%。开放阅读框(ORF)搜索发现其含有3个ORF,ORF1长594 nt并编码197个氨基酸(2027~2620,197 aa),ORF2 长1188 nt并编码395个氨基酸(903~2090,395 aa),未获得完整的ORF3。ORF1编码蛋白与无花果幽影病毒(Fig umbra-like virus)的P3蛋白相似性最高,氨基酸序列覆盖率为82%,相似性为36.97%,ORF2编码RNA依赖性RNA聚合酶(RNA-dependent RNA polymerase)与幽影病毒属大多病毒覆盖率在68%~99%,相似性42.47%~51.12%,与仙人掌幽影病毒(Opuntia umbra-like virus, GenBank:AXG65483.1)相似性最高,为51.12%。如图2所示,使用最大似然法对ORF2构建系统发育树,发现永青花椒相关幽影病毒与仙人掌幽影病毒(Opuntia umbra-like virus)及玉米相关幽影病毒(Maize-associated umbra-like virus)等病毒进化关系较近,幽影病毒属大部分为正义阳性单链RNA,有4个开放阅读框,由于其基因组不编码结构蛋白,受幽影病毒属感染的植物通常伴有对应的某一种黄症病毒(Iuteovirus)复合侵染,黄症病毒对幽影病毒进行基因组衣壳化并通过蚜虫作为介体传播[11-12],虽然序列不全,但对获得的3个开放阅读框分析发现该病毒与幽影病毒属病毒关系密切,且6个样品测序刚好检测出属于黄症病毒科的四川青椒耳突花叶病毒,幽影病毒的辅助病毒为黄症病毒科成员,两者或有关联。
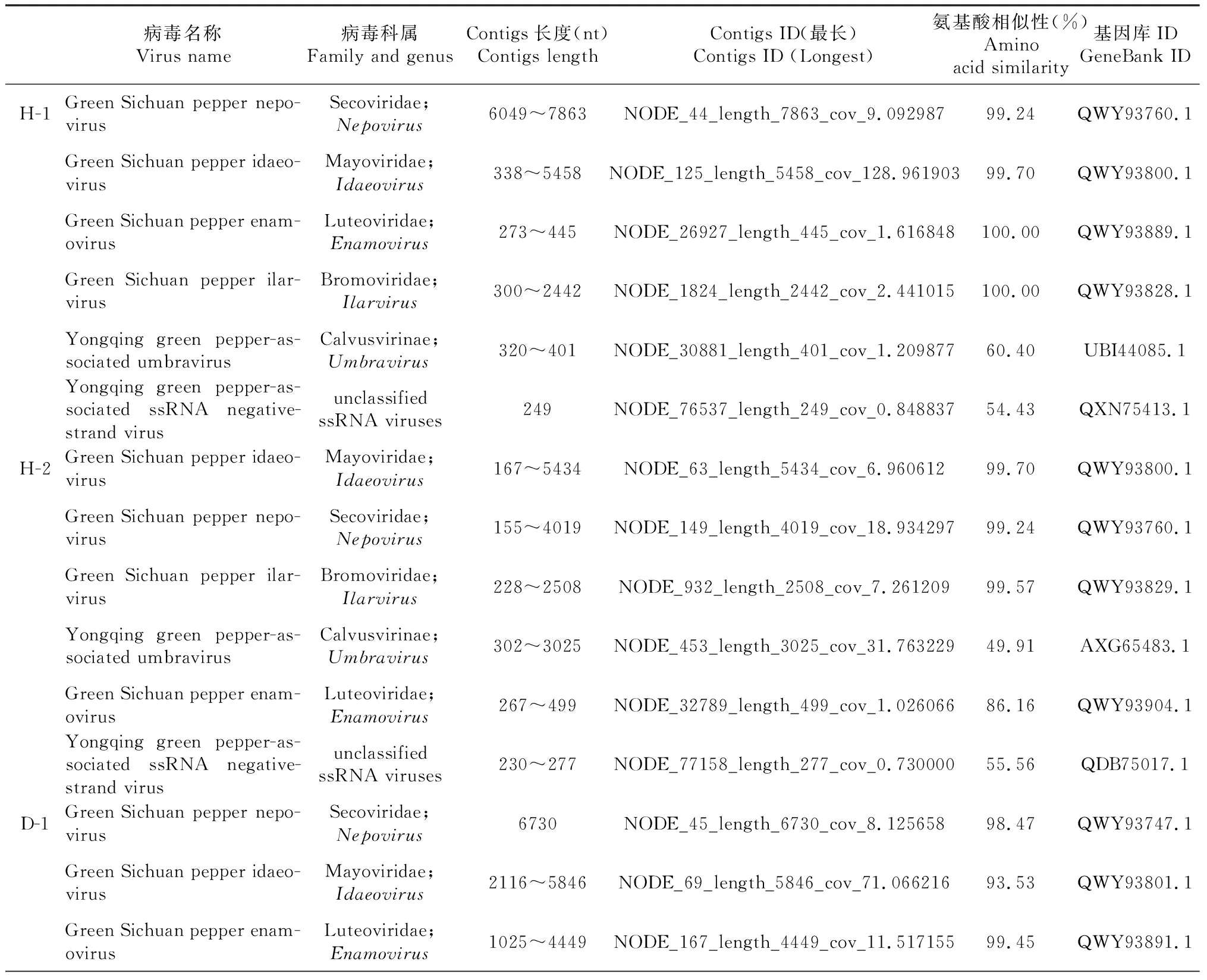
表1 不同样品中的主要病毒种类

续表1 Continued table 1

表2 感病样品中Betaflexiviridae病毒种类
2.3 永青花椒相关负链RNA病毒分析
由表1可知,6个样品中得到长度为30~7306 nt的永青花椒相关负链RNA病毒(Yongqing green pepper-associated ssRNA negative-strand viruse)contigs,其中D-3得到最长读数片段长为7306 nt,与其余短片段核酸进行两两比对发现相似性为94.6%~100.0%,通过NCBI BLASTX发现与枝孢霉菌负链RNA病毒1(Cladosporium cladosporioides negative-stranded RNA virus 1,GenBank:QDB75017.1)最接近,覆盖率为99%,相似性为66.58%,DNAMAN比对核酸序列相似性为42.2%。开放阅读框检索发现含有1个ORF,长7041 nt,编码2346个氨基酸,111~7151 nt, 2346 aa),主要为RNA依赖性RNA聚合酶,与其余未分类单链负义RNA病毒(Unclassified ssRNA negative-strand viruses)覆盖率42%~99%,相似性23.25%~64.49%。如图3所示,使用最大似然法对ORF进行系统发育分析,绘制系统发育树发现永青花椒相关负链RNA病毒与枝孢霉菌负链RNA病毒1系统发育最为接近,其余相近病毒也皆为真菌传播病毒,认为该病毒或为枝孢霉菌相近的真菌病毒,该菌种及病毒有待验证。
2.4 乙型线形病毒科病毒分析
将测序文库与NCBI数据库比对,获得与FYD相关且仅在D组检测的乙型线形病毒科的PVM、PVH、PVS的序列号,使用NDAMAN拼接不同病毒序列,得到病毒近全长序列。从表2可知,GRSPaV仅在D-1中发现,其他感病样品中均未发现,所匹配的contigs片段仅为434 和294 nt,无法拼接出全长,NCBI BLASTX序列比对与GRSPaV isolate NV-57G 分离物的复制酶蛋白位点相似性为99.25%,可确定为该病毒。
D2~D4样品中多为PVM、PVH、PVS 2种病毒或3种病毒复合侵染,通过DNAMAN序列拼接得到总长分别为8436、8353、8510 nt的PVM、PVH、PVS病毒的近全长序列,提交至NCBI进行序列比对,与PMV 分离物M57(GenBank: AY311395.1)、PVH 分离物YN(GenBank: JQ904630.10)、PVS 分离物BY(GenBank: MF033144.1)核苷酸相似性分别为96.5%、96.6%、97.9%以及98.6%。PVM、PVH、PVS均归属于香石竹潜隐病毒属(Carlavirus),大多为蚜虫非持久性传播,在马铃薯上常表现为混合感染,其基因组特征为有6个ORF[13],PVM、PVH、PVS的ORF1(~5900 nt)编码甲基转移酶(321 aa,pfam01660)、C23肽酶(87 aa,pfam05379)、病毒解旋酶1(253 aa,pfam01443)、RNA依赖性RNA聚合酶(RdRp,389 aa,pfam00978),重叠的ORF2、3、4(~1200 nt)为负责胞间运动的3个基块(TGB)[14],ORF5、ORF6分别编码香石竹潜隐病毒属特异性CP蛋白(304 aa,pfam08358)、carlaviruses推定的核酸结合蛋白(114 aa,pfam01623),ORF序列比对分析表明本研究所获得匹配到PVM、PVH、PVS的contigs与相应代表性分离物具有较高相似性。
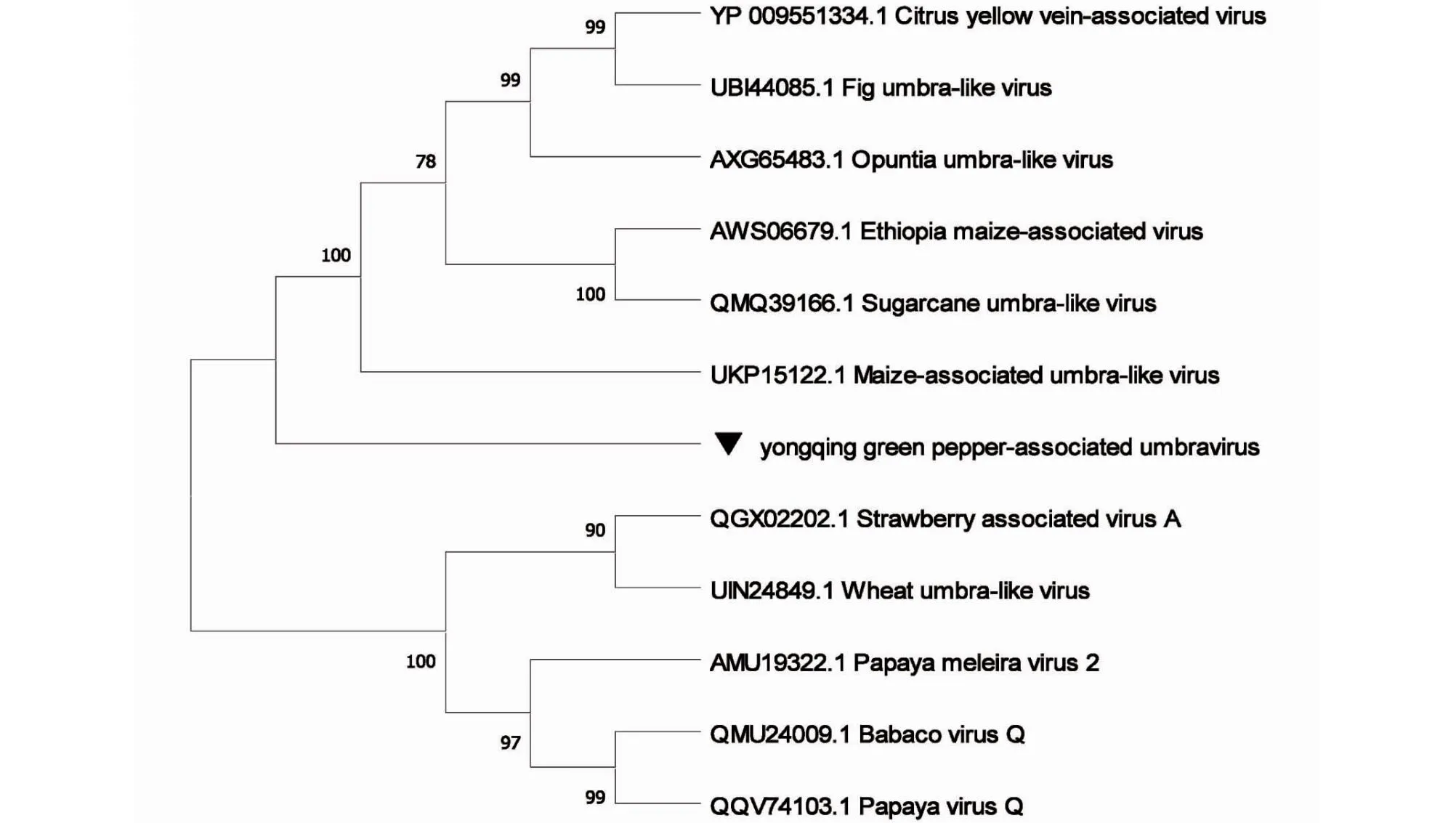
图2 基于ORF2核苷酸序列的永青花椒相关幽影病毒系统发育树Fig.2 Phylogenetic tree of Yongqing green pepper-associated umbravirus based on the nucleotide sequences of ORF2

图3 基于全长核苷酸序列的永青花椒相关负链RNA病毒系统发育树Fig.3 Phylogenetic tree of Yongqing green pepper-associated ssRNA negative-strand virus based on the complete nucleotide sequences
3 讨 论
植物病害已成为影响经济效应最显著因素之一,因此需要在最短的时间内找到有效检测工具对病原进行快速且特异性的检测,而对植物病害的预防和管理在很大程度上取决于对病原的准确识别。现阶段HTS比起从前测定的方法更具有全面性,可作为常规病原体筛查的一部分,已成功应用于发现许多农作物病毒、类病毒[15],如使用HTS对葡萄总RNA进行测序鉴定出4种病毒和3种类病毒,揭示多重感染的病毒之间可能有复杂的相互作用[16]。Cao等[17]用HTS技术从九叶青花椒黄花病样品中发现4种新病毒,这4种RNA病毒在同种植物混合感染中具有水平传播的共性,线虫多面体病毒属病毒主要是长线虫非持久性传播,种子/花粉传播也是常见传播方式。耳突花叶病毒属通过花粉和种子传播,未见生物媒介传播的报道[18],悬钩子病毒属病毒由蚜虫以持续循环和机械接种的方式传播[19]。核型弹状病毒属由节肢动物或壶菌传播,有报道称叶蝉或蚜虫可传播此类病毒[20 -21]。4个新病毒中,GSPNeV在宿主中具有高表达的特性且传播更广泛,且后续实验发现与FYD相关的是GSPNeV及satGSPNeV,统计学显著性分析证明两者之间存在密切的共感染能力,又因其在患病植物有较高读数占比,证明高病毒滴度对植物宿主正常生理活性可产生一定程度的影响,从感病花椒未显症枝条与显症枝条的HTS测序发现GSPNeV几乎仅存在于显症枝条中,认为GSPNeV及sat GSPNeV与FYD密切相关,但从无症状树也检测到GSPNeV,预示此类病毒可能存在临床潜伏期,又或跟病毒滴度含量低于植株出现感病症状的阈值而未显症有关,不排除感染GSPNeV的无症树木可能是对病毒感染具有某方面抗性的栽培品种,在仅考虑感病花椒的前提下认为GSPNeV可作为侵染植体的优势病毒[7,22],但仍然未完成病毒病原的柯赫氏法则检验。本研究使用HTS对6个永青花椒样品进行了测序,在无症、感病组花椒中均发现有Cao等[7]从九叶青花椒黄花病样品中发现的4种新病毒,再一次证明了花椒黄花病与这4个病毒的相关性,也说明该类病害的普遍性。本研究还发现2种新病毒,暂命名为永青花椒相关幽影病毒和永青花椒相关负链RNA病毒。另外,本研究的花椒黄花病样品也发现乙型线型病毒科的几种病毒。仅样品D-1中检测出与GRSPaV相似的contigs片段,可能该病毒在宿主内浓度滴度较低,测序时未能测出完整的病毒序列,而 GRSPaV 被验证出在宿主植物葡萄藤的生长发育中具有降低宿主光合作用率和防御机制的能力[23],受GRSPaV感染的花椒或可能会因该病毒使花椒防御机制减弱而引发黄花病症状。
从样品D-2、D-3、D-4等感染黄花病花椒可以测得的PVM、PVH、PVS完整序列,并且contigs读数较高,表明植物宿主内病毒浓度滴度在较高水平。其中D-4检测出的香石竹潜隐病毒属病毒contigs读数最多,病毒种类丰度最高,D-3、D-4同时受PVM、PVH、PVS 3种病毒混合感染。PVM被验证拥有“双重策略”以抵抗宿主植物的抗病毒沉默,其编码的富含蛋胱氨酸的CRP蛋白可在单细胞水平抑制RNA沉默和沉默扩散,作用效应发挥于局部,还会增加病毒RNA积累,而TGB1除促进细胞运动外仅在沉默扩散时发挥抑制RNA沉默的效应[24],除此之外,携带香石竹潜隐病毒属CRP蛋白的马铃薯X病毒(PVX)发现可改变PVX侵染的症状表型,接种叶片原PVX感染表征从无症感染及轻度花叶转变为坏死的局部损伤和顶端叶片坏死[25]。从测序花椒样品中可以发现,显症组D-2、D-3、D-4与正常组H-1、H-2最大的区别就是均遭受香石竹潜隐病毒属病毒侵染, FYD或由协同侵染花椒宿主的香石竹潜隐病毒属病毒通过抑制宿主花椒抗病毒沉默以及增强某些病毒毒性从而导致花椒宿主显症。同原理的现象有蚕豆萎蔫病毒2号(Broad bean wilt virus 2,BBWV-2)编码的VP53,VP37和LCP抑制宿主基因沉默所产生的siRNAs的积累,使PVX致病性增强[26],但此推测有待通过对正常花椒植株接种PVM、PVS、PVH混合病毒加以验证。
在昭通花椒种植基地观察发现,FYD显症花椒植株受蚜虫大面积覆盖感染,研究表明Carlavirus属病毒可通过蚜虫以非持久方式传播[10],蚜虫也是幽影病毒的传播介体,但花椒黄花病症是否由蚜虫传播病毒引起,有待后续实验验证。
随机抽查FYD花椒样品进行植原体验证并未检测到植原体侵染,无法解释FYD与植原体感染有直接关联,发现了与真菌病毒——枝孢霉菌负链RNA病毒1序列相似的永青花椒负链RNA病毒,但该病毒的真菌寄主菌种有待确定,由于不同宿主对同类病原感染效应不同,该真菌是否导致花椒产生黄花病也需要后续对正常花椒单独接种该菌进行验证分析。
本研究测序选择的样品相对少,对花椒FYD相关病原分析具有一定局限性,无法准确说明FYD疾病爆发的因素是否与上述病毒具有潜伏期有关,也无法排除测序样品中包含个别对引发FYD直接相关病毒具有低敏感性而不显症的植株的可能,但从感病样品中检测出了高表达的香石竹潜隐病毒属病毒,该结果或可视为引发FYD症状的因素之一。Cao等报道的4个病毒,通过田间现场调查、HTS分析、RT-PCR验证,结果显示GSPNeV在宿主中具有高表达特点,GSPEV传播更广泛,但其并未表述直接导致病症的病毒,且其余病毒是否相关仍需要开展严格的病毒鉴定(如柯赫氏法则分析等)才能明确其病原[7-8]。因此,初步分析PVM、PVS、PVH与FYD症状可能相关,基于测序结果发现未报道的永青花椒幽影病毒、永青花椒负链RNA病毒也需要后续研究其在单独感染花椒时是否会造成黄花病症,同时不排除病毒协同效应导致花椒黄花病症的发生。
4 结 论
本研究利用HTS技术从昭通竹叶花椒永青1号中均发现已报到的四川青花椒病毒(GSPIV、GSPNeV、GSPEV、GSPNuV)和未报道的永青花椒幽影病毒、永青花椒负链RNA等多种病毒,其中PVM、PVS、PVH等乙型线形病毒科病毒仅从感病组中发现,通过RT-PCR验证了GSPNeV、PVM等病毒与本研究样品的相关性,推测高表达的香石竹潜隐病毒属病毒或可诱发花椒FYD症状。本研究为后续明确花椒黄花病的病原奠定了理论基础。
猜你喜欢
今日农业(2022年14期)2022-09-15
新民周刊(2022年27期)2022-08-01
今日农业(2022年3期)2022-06-05
基层中医药(2021年8期)2021-11-02
传染病信息(2021年6期)2021-02-12
今日农业(2020年15期)2020-12-15
黄河之声(2020年16期)2020-11-05
今日农业(2020年16期)2020-09-25
作文通讯·高中版(2017年11期)2017-12-20
海峡姐妹(2017年9期)2017-11-06
