
Continent-wide vocal leapfrog pattern in Collared Scops Owls obfuscates species boundaries
2024-01-22 08:24MengYueWuFrankRheindt
Avian Research 2023年4期
Meng Yue Wu, Frank E.Rheindt
aDepartment of Biological Sciences,National University of Singapore,16 Science Drive 4,Singapore,Republic of Singapore
bDepartment of Environmental Sciences,Zoological Institute,University of Basel,Basel,Switzerland
Keywords:Asia Biogeography Species delimitation Taxonomy Bioacoustics
ABSTRACTMuch confusion has surrounded the taxonomy of the Collared Scops Owl (Otus bakkamoena) complex, distributed widely across tropical and subtropical Asia.Often divided into three species, modern taxonomies disagree on its treatment, as a lack of deep mitochondrial DNA divergence is seemingly in contrast with patterns of vocal differentiation.Analysing a dataset of territorial calls of 122 individuals across Asia, we identified a vocal leapfrog pattern in which owl populations at the eastern and western ends of the distribution sound more similar to each other but are separated by different-sounding populations in between.The boundaries between vocal types are not congruent with traditional species boundaries.Our data allow for a re-drawing of species boundaries and reveal that Collared Scops Owls follow a biogeographic pattern rare in Southeast Asian birds.However, this pattern is shared with several other owl complexes in that peninsular Malaysian populations form part of a continental Asian species centred further north in Southeast Asia rather than forming part of a Sundaic species centred in the adjacent Indonesian Archipelago.
1.Introduction
Reproductive isolation is a key factor in speciation, as it prevents interbreeding between populations and allows for the accumulation of differences (Mayr, 1942).While morphological features are implicated in reproductive isolation across most animals, bioacoustic characters can be of primary importance in some bird families (Isler et al., 1998;Rheindt et al., 2008; Ng et al., 2016), especially in nocturnal ones such as owls (Strigidae) in which optical cues are less relevant (Howell and Robbins, 1995; King, 2002; Sangster et al., 2013; Pons et al., 2013; Flint et al., 2015; Krabbe, 2017; Gwee et al., 2017, 2019; Dantas et al., 2021;Movin et al., 2022).
Properly diagnosing species-specific differences in vocalizations is key in correct species delimitation in optically cryptic assemblages such as owls.Making sense of the geographic variation in bioacoustic traits can be challenging when such characters are distributed in complex ways, such as when they form a leapfrog pattern (Remsen, 1984).A leapfrog pattern is one in which two populations at opposing ends of a distribution range resemble each other in certain character traits, but are separated by intervening populations that differ in these traits, either along a geographic continuum or in a non-clinal fashion (Remsen,1984).While most leapfrog patterns documented by evolutionary biologists refer to morphological characters, bioacoustic leapfrog patterns are known from various tropical bird groups (Cadena and Cuervo, 2010;Rheindt et al., 2011).
Little is known about the mechanisms that create leapfrog patterns;they may be implicated in speciation events (Remsen, 1984; Rheindt et al., 2011).The traits of the terminal populations that are geographically distant may reflect the ancestral character state, while populations in the middle may be on their way to becoming a different species (if they have not already speciated), and hence diverge in specific characters from populations on either side.Studying bioacoustic leapfrog pattern could be a promising avenue to provide insights into the mechanisms of speciation in cryptic bird groups such as owls.In this study, we uncover a vocal leapfrog pattern in the Collared Scops Owl(Otus bakkamoena) complex that has long been overlooked, and we shed light on the turn-over of bioacoustic traits in this confusing group, which has been marked by much taxonomic confusion.
The Collared Scops Owl complex, which has been one of the taxonomically most controversial owl assemblages, ranges across tropical and subtropical Asia.It comprises roughly 15 subspecies-level taxa(Fig.1), often divided into anywhere between 1 and 3 species-level lineages including the Indian Scops Owl (O.bakkamoena) from the Indian subcontinent, the Collared Scops Owl (O.lettia) from the Himalayan foothills and large parts of subtropical East and Southeast Asia, and the Sunda Scops Owl (O.lempiji) from tropical Southeast Asia (Fig.1)(Robson and Allen, 2005; Eaton et al., 2021; Clements et al., 2022;Handbook of the Birds of the World & BirdLife International, 2022).An alternative taxonomic treatment has divided the complex into four species, with the 3-species split outlined above in addition to the separation ofcnephaeusfrom the Malay Peninsula and Singapore into a monotypic species (K¨onig and Weick, 2008).
Vocal variation within the Collared Scops Owl complex has been described in multiple ornithological treatises and studies (Najmi-Hanis et al., 2016; Yee et al., 2016; Eaton et al., 2021), but has thus far not resulted in one unequivocal taxonomic interpretation.In this study, we used a quantitative bioacoustic approach to characterize the territorial vocalisations of the Collared Scops Owl complex across its range.We used these new insights to revise the taxonomy of the entire complex,emulating work on multiple other owl complexes (Howell and Robbins,1995; King, 2002; Rasmussen et al., 2022; Pons et al., 2013; Sangster et al., 2013; Flint et al., 2015; Sadanandan et al., 2015; Krabbe, 2017;Gwee et al., 2019; Dantas et al., 2021; Movin et al., 2022) and other Asian bird species (Ng et al., 2016; Ng and Rheindt, 2016; Prawiradilaga et al., 2018; Rheindt et al., 2011).
2.Materials and methods

Fig.1.Distribution of the three scops owl species of the Collared Scops Owl complex (green:Otus lempiji, blue:O.lettia, orange:O.bakkamoena) according to the present study.Species level boundaries are indicated by the three different colors, with subspecies boundaries overlaid in stippled lines and subspecies names given.The black dots represent localities of vocal recordings used in this study.Representative sonograms of each of the three species shown in lower left, with an additional sonogram of the subspeciescnephaeus, long attributed toO.lempijior considered a monotypic species, but here assigned toO.lettia.(For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
We compiled 213 sound recordings from the online sound library xeno-canto (https://www.xeno-canto.org) filed underO.lempiji,O.lettiaandO.bakkamoena(Appendix Table S1).To ensure measurements of vocal homologs across the complex, we focused only on their territorial call—a single short note.Recordings that did not feature the territorial call were removed.Many birds, includingOtusowls, are known to return to the same breeding site with the same partner for many years(Greenwood, 1980; Linkhart and Reynolds, 2007).To avoid duplicating vocalisations of the same individual, only one recording made by the same person at the same site and time was retained, and we additionally retained only one out of multiple recordings within a five-year period if they were taken at a narrow specific locality.In total we measured 122 recordings of the main territorial call for quantitative vocal analysis,with each recording representing one individual, after removal of poor-quality recordings (Fig.1; Appendix Table S1).
We used Raven Pro 1.5 (Bioacoustics Research Program, Cornell Laboratory of Ornithology, Ithaca, NY, USA) to inspect and measure parameters of each recording.Although different recording equipment was used among recordists, spectrogram inspection revealed that there was negligible equipment bias.All notes made by the same individual were measured up to 20 times, when available, to account for withinindividual variation.With the exception of the window size being set to 1000 to obtain the best resolution across all recordings, default settings of Raven Pro were used.The contrast and brightness of the sonograms were adjusted to ensure measurements were carried out under similar settings across all recordings.We measured a total of six parameters: (1) lowest frequency of the call; (2) highest frequency of the call; (3) duration of the call; (4) peak frequency of the call; (5) change in peak frequency between the first third and second third of the call; (6)change in peak frequency between the second third and the last third of the call.As preliminary inspection of the sonograms revealed distinct element shapes in different populations (Fig.1), we used (5) and (6) to quantify this change.
R 3.3.2 (R Core Team, 2009) was used to conduct principal component analysis (PCA).We also computed Isler et al.’s (1998) vocal diagnosability criterion, commonly known as the Isler criterion.This criterion has been used in bioacoustic species delimitation across a wide variety of Asian birds, including owls (Gwee et al., 2017, 2019; Movin et al., 2022).The Isler criterion uses the standard deviation of the sample points rather than the taxon mean, and as such is substantially more discriminating than thet-test or the Mann-WhitneyUtest (Isler et al.,1998; Rheindt et al., 2011; Ng and Rheindt, 2016; Gwee et al., 2017).
3.Results
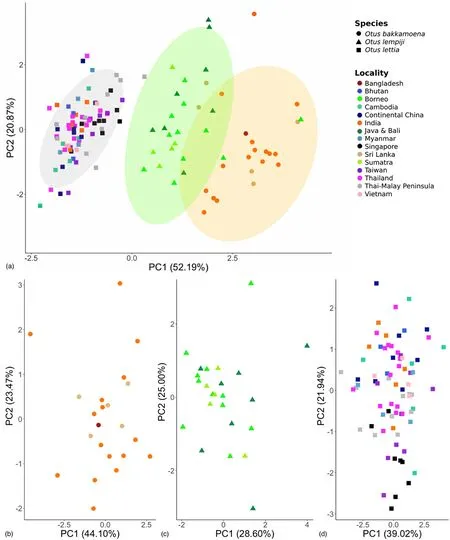
Fig.2.Principal component analysis plots based on the six vocal parameters measured.(A) With all samples (n=122) included, the first principal component (PC1)chiefly differentiates the three species level taxa within the complex.Colored ellipses denote the 95% confidence intervals forOtus lettia(grey),O.lempiji(green) andO.bakkamoena(orange).(B) Samples within theO.bakkamoenacluster (n=22) do not separate into distinct groups.(C) Samples within theO.lempijicluster (n=24)equally do not separate into distinct groups.(D) Samples within theO.lettiacluster (n=76) appear to form a geographic cline from south to north along PC2.(For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
On the basis of the six vocal parameters analysed herein, we found three independent PCA clusters that correspond largely but not entirely with the existing species level division of the complex intoO.bakkamoena,O.lempijiandO.lettia(Fig.2A).The first and second principal components explained 52.2% and 20.9% of data variation,respectively (Fig.2A).The greatest difference to the traditional taxonomic arrangement related to the taxon from the Malay Peninsula and Singapore,cnephaeus, long classified underO.lempiji, but in our analysis grouping with the continental Southeast Asian speciesO.lettiainstead.Our bioacoustic data also revealed that the two geographically disjunct speciesO.bakkamoenafrom the Indian subcontinent andO.lempijifrom the Indonesian Archipelago (Fig.1) are more similar to each other as compared to the vocally more divergent but geographically intervening speciesO.lettia(Fig.2A).
A closer bioacoustic analysis within each species cluster did not reveal additional vocal population structure inO.bakkamoenaandO.lempiji(Fig.2B and C).WithinO.lettia, on the other hand, a geographical cline appeared to emerge along PC2 from the south of the range (Singapore,Thai-Malay Peninsula) towards the north (Himalayan foothills, continental China) (Fig.2D).This result was consistent with conclusions from qualitative vocal inspections showing that the element shape of the territorial call slowly changes from a flat note in the southern Malay Peninsula to a rising inflection as one moves north (Fig.1).
Means and standard deviations of all six vocal measurements varied among taxa (Fig.3).The measurements betweenO.bakkamoenaandO.lettiawere mostly non-overlapping, whileO.lempijiwas often intermediate between the other two in vocal parameter measurements(Fig.3).Pairwiset-tests comparing all six parameters between the taxa demonstrated significant differences (p≤0.05) for most comparisons,with the exception of comparisons betweenO.lettiaandO.lempijiin lowest frequency as well asO.bakkamoenaandO.lempijiin highest and peak frequency (Appendix Table S2).However, under the Isler criterion,shown to be more discriminating than the commont-test (Isler et al.,1998), no diagnosable traits were found in any pairwise comparisons.
4.Discussion
We provide a quantification of territorial call variation of the members of the Collared Scops Owl complex across its natural range(Fig.1).We identified three call shapes—‘inflected’, ‘deflected’ and‘level’ (Fig.1)—using vocal parameters that had been used to successfully delimit other owl species (Gwee et al., 2019; Movin et al., 2022),partly in conjunction with genomic results (Gwee et al., 2017).
4.1.Vocal leapfrog pattern in the Collared Scops Owl complex
The three commonly accepted species-level taxa of the Collared Scops Owl complex occur along an east-west axis from the Indian subcontinent across continental Southeast Asia down to the western Indonesian archipelago (Fig.1).We show that the two terminal species,O.bakkamoenain the west andO.lempijiin the east, are vocally closer to each other than towards the geographically interveningO.lettia(Fig.2).In many respects,O.lempijiwas intermediate betweenO.bakkamoenaandO.lettiain measurements of six vocal parameters, with a few exceptions (Fig.3; Appendix Table S2).We identified distinct element shapes in territorial calls diagnosable even by the human eye and ear for each species-level taxon (Fig.1).The fact that the geographically centralO.lettia, with its vigorous and often deflected call note, stood out within the complex from the two peripheral forms characterized by a more inflected call note points to a bioacoustic leapfrog pattern (Cadena and Cuervo, 2010; Rheindt et al., 2011).
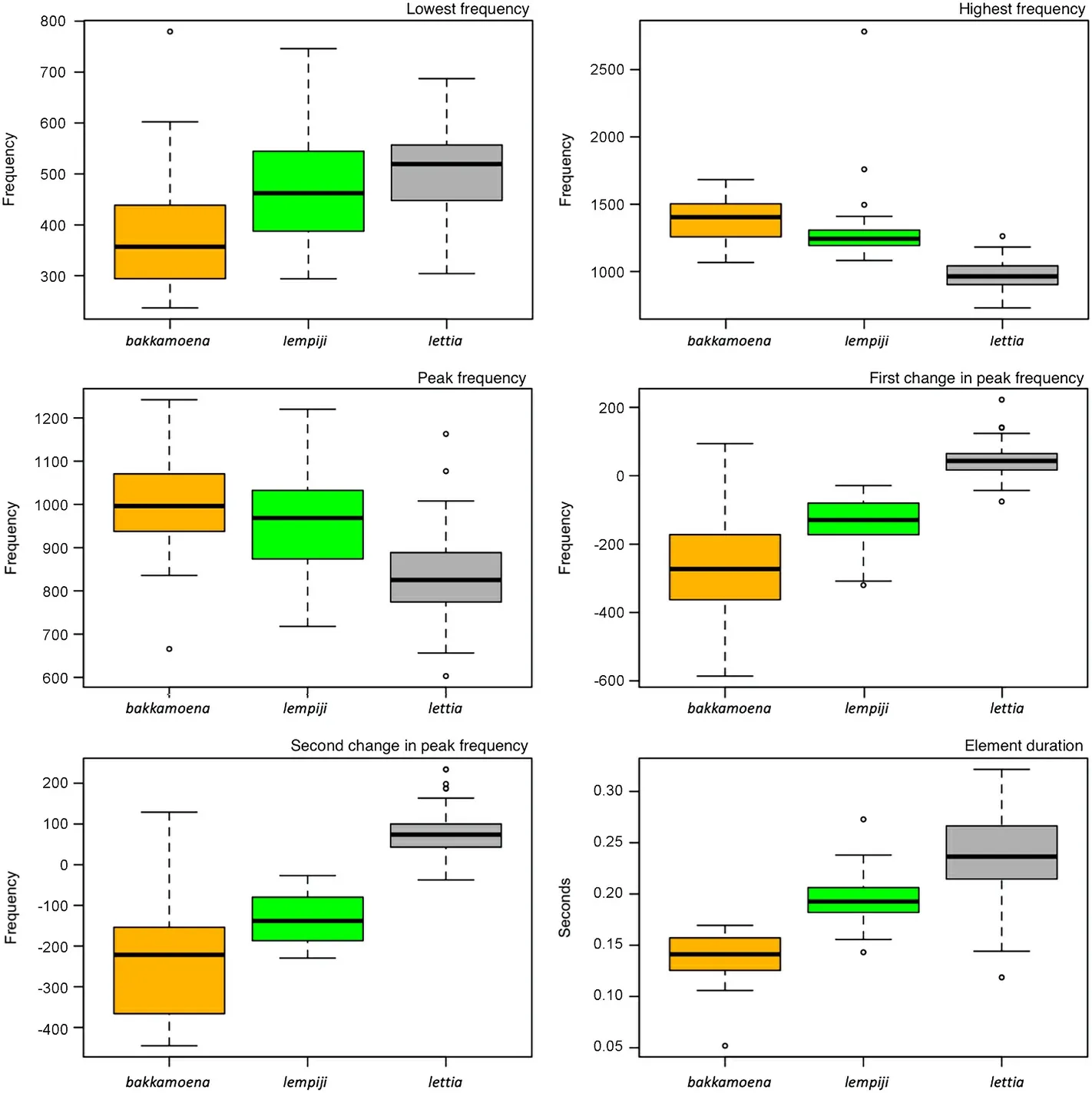
Fig.3.Boxplots for all six vocal parameters for the three species as here defined.
In a classical study, Remsen (1984) concluded that leapfrog patterns primarily evolve in regions characterized by geographic linearity,topographic relief and high species riches.Meanwhile, bioacoustic leapfrog patterns in particular have also been detected in insular settings(Rheindt et al., 2011).Here we showcase a vocal leapfrog pattern across the continental scale in a bird group with a broad elevational tolerance,supporting the notion that this pattern may be much more pervasive in birds than currently assumed.
In owls vocalisations are innate and play an important role in mate selection; they are thus thought to be under strong selective pressure(K¨onig and Weick, 2008).Vocal differentiation can act as a mating barrier between populations, potentially leading to speciation.In Collared Scops Owl vocalizations, the geographically terminal phenotype, i.e., the inflected call note ofO.lempijiandO.bakkamoena, is likely plesiomorphic (=ancestral) while the central populations (O.lettia)evolved a novel phenotype, a deflected call note, possibly promoted by character displacement near the range limits between the three evolving species to prevent hybridisation in contact zones.This observation particularly applies to the contact zone betweenO.lettiaandO.bakkamoena, whose deflected and inflected call notes, respectively,meet quite abruptly along a narrow ecotone in the Himalayan foothills of the northern Indian subcontinent (Rasmussen and Anderton, 2005).
4.2.Mapping turn-over between O.lettia and O.lempiji
The contact zone betweenO.bakkamoenaandO.lettiaruns along the foothills of the Himalayas, a common suture zone between species pairs in birds, e.g., between Red (Gallus gallus) and Grey Junglefowl(G.sonneratii) (McGowan and Kirwan, 2020a, 2020b), or between Blyth’s (Terpsiphone affinis) and Indian Paradise-Flycatcher (T.paradisi)(Moeliker et al., 2020; Rasmussen et al., 2022).The vocal turn-over between the two scops owls here is sharp and occurs across only a few kilometers (Rasmussen and Anderton, 2005).
In contrast, the geographic boundary betweenO.lettiaandO.lempijiis much less well understood.Most current taxonomic treatments assign the taxoncnephaeusfrom the Malay Peninsula toO.lempiji, which would place the boundary between the two species somewhere along the Isthmus of Kra between the Thai-Malay Peninsula and the main Southeast Asian continent (Holt et al., 2020a, 2020b).However, our bioacoustic data place samples ofcnephaeusfrom Singapore and the Thai-Malay Peninsula firmly withinO.lettia, suggesting that the actual turnover zone is across the Singapore Straits that cut between Sumatra and the Peninsula (Figs.1 and 2).
Qualitatively, the southern peninsular and Singaporean populations of the taxoncnephaeushave a unique vocal signature, characterized by a fairly level-pitched call, which gradually and clinally shifts into a deflected call note as one moves north along the Peninsula until one reaches populations near the Isthmus of Kra, which are the first to sound unequivocally deflected like other populations ofO.lettia(Fig.1).Conversely, when moving from the southernmost tip of the Peninsula across the Singapore Straits to Sumatra, there is a sharp turn-over from the level-pitched call of the taxoncnephaeusto the distinctly inflected call note ofO.lempiji, which is reflected in the distant placement of Sumatran and Singaporean samples in the bioacoustic PCA (Fig.2A).
The taxoncnephaeusfrom the Thai-Malay Peninsula and Singapore has previously been identified as distinct from other surrounding taxa(Deignan, 1950), and has even been mooted to warrant recognition at the species level due to its smaller body size, other morphological traits and its distinct level-pitched call (Wells, 1999; K¨onig and Weick, 2008).Our more comprehensive bioacoustic inquiry now reveals that this level call, however, is connected to more deflected call notes to the north through a gradual geographic cline, supporting subspecies status ofcnephaeusunderO.lettia, as reflected in bioacoustic PCA (Figs.1 and 2).
4.3.The biogeographic boundary between Sundaland and the continental monsoon region may differ between owls and other birds
Many tropical Asian bird species complexes are divided into an equatorial rainforest species centred around the western Indonesian Archipelago and a closely related monsoon-forest inhabiting species from the Asian mainland (Eaton et al., 2021).Whenever birds display such a biogeographic constellation, the Sundaic species virtually always extends north to the Thai-Malay Peninsula and reaches its northern limit at the Isthmus of Kra, where the range of the continental monsoon species takes over.Owls of the Collared Scops Owl complex were long thought to follow this geographic pattern, withcnephaeusfrom the Thai-Malay Peninsula invariably assigned toO.lempijifrom Indonesia.However, our bioacoustics-based reassignment ofcnephaeustoO.lettiasuggests that these scops owls defy this biogeographic pattern, as the continentalO.lettiaextends south all the way to Singapore (Fig.1).
Collared Scops Owls may not be the only owl group that departs from this biogeographic rule.In another recent taxonomic revision based on bioacoustic characters, Gwee et al.(2019) divided the Collared Owlet(Glaucidium brodiei) into two species, a continental one reaching south to the Thai-Malay Peninsula and an archipelagic one restricted to Sumatra and Borneo in the Indonesian Archipelago.A third such biogeographic arrangement is not universally accepted and relates to the Brown Wood Owl (Strix leptogrammica) from tropical and subtropical Asia, which was recently separated into a mainland species (Brown Wood OwlS.indranee) and an Indonesian archipelagic species (Sunda Wood OwlS.leptogrammica) based on pronounced differences in call notes (Eaton et al., 2021).The reasons why owls defy the usual biogeographic pattern and have Malay peninsular populations that are related to Asian mainland species rather than Indonesian archipelagic ones is puzzling and will require future inquiry.
4.4.Taxonomic treatment of the Collared Scops Owl complex
The results of our vocal analysis of the Collared Scops Owl species complex allow for taxonomic conclusions.A genetic study detected limited divergence in this complex (Fuchs et al., 2008), which has tempered many taxonomists’ desire to divide the species complex based on bioacoustic differences that have been known for decades.However,the genetic signature was dominated by the information signal of mitochondrial DNA, which can be subject to biases, especially as relates to shallow divergences caused by genetic introgression events masking deeper genomic divergence (e.g., Rheindt and Edwards, 2011).Our analyses demonstrate deep bioacoustic differences among the three main species-level groups within the complex, albeit with a reassignment ofcnephaeusfromO.lempijitoO.lettia.Vocal turn-over is steep wherever the three species come close to a neighbour, and our vocal PCA dispels the notion of any bioacoustic clinality among the three species.The only vocal cline detected was withinO.lettiaas here circumscribed,including populations with a deflected call note over most of the range gradually turning to a more level-pitched call note on the Thai-Malay Peninsula within the range ofcnephaeus.Our lack of vocal material for the taxoncondorensis, confined to Con Son Island off the southern tip of Vietnam, precludes a certain assignment to either species.But given the location of this island on the Sunda Shelf at the northern limit of a former rainforest bloc of subcontinental proportions, and based on the results of recent genomic analyses shedding light on the affinities of taxa on nearby islands (Garg et al., 2022), we provisionally retain the most widely-followed treatment ofcondorensisas underO.lempiji.
The following is our proposed taxonomic treatment of the Collared Scops Owl (O.bakkamoena) complex.Distributions of all the taxa involved are shown in Fig.1:
●Otus bakkamoenaPennant, 1769: Indian Scops Owl
○O.b.bakkamoenaPennant, 1769: southern India and Sri Lanka
○O.b.gangeticusTicehurst, 1922: northern India along the Ganges drainage
○O.b.marathaeTicehurst, 1922: northern peninsular India
○O.b.deserticolorTicehurst, 1922: arid northwestern India and Pakistan
●Otus lettia(Hodgson, 1836): Collared Scops Owl
●O.l.lettia(Hodgson, 1836): central Himalayan foothills through Myanmar to Indochina
●O.l.plumipesHume, 1870: western Himalayan foothills
●O.l.erythrocampe(Swinhoe, 1874): southern Chinese mainland,northern Vietnam
●O.l.glabripes(Swinhoe, 1870): Taiwan Island
●O.l.umbratilis(Swinhoe, 1870): Hainan Island
●O.l.cnephaeusDeignan (1950): Thai-Malay Peninsula, Singapore
●Otus lempiji(Horsfield, 1821): Sunda Scops Owl
●O.l.lempiji(Horsfield, 1821): Java, Bali, southern Sumatra,southern Borneo
●O.l.hypnodesDeignan (1950): northern and central Sumatra
●O.l.lemurumDeignan, 1950: northern Borneo
●O.l.kangeanusMayr, 1942: Kangean Islands
●O.l.condorensisKloss, 1930: Con Son Island (= Condor Island,Vietnam)
Funding
This work was supported by a Singapore National Research Foundation (NRF) Investigatorship (WBS A-8001397-00-00) to FER.
CRediT authorship contribution statement
Meng Yue Wu:Data curation, Methodology, Writing – original draft,Writing – review & editing.Frank E.Rheindt:Conceptualization,Methodology, Writing – review & editing.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
We are indebted to the recordists who have submitted their sound material to xeno-canto sound archive (https://www.xeno-canto.org).We thank Geraldine Lee for her preliminary exploration of the vocalizations, and Movin S/O Nyanasengeran for general assistance.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.avrs.2023.100141.

- Avian Research的其它文章
- Selecting the best: Interspecific and age-related diet differences among sympatric steppe passerines
- Morphology and morphometry of two hybridizing buntings at their hybrid zone in northern Iran reveal intermediate and transgressive morphotypes
- Quiet in the nest: The nest environment attenuates song in a grassland songbird
- Characteristics of cross transmission of gut fungal pathogens between wintering Hooded Cranes and sympatric Domestic Geese
- Fecal DNA metabarcoding reveals the dietary composition of wintering Red-crowned Cranes (Grus japonensis)
- Short-term night lighting disrupts lipid and glucose metabolism in Zebra Finches: Implication for urban stopover birds