
鸡经济性状遗传标记挖掘与基因组选择研究进展
2024-01-13 07:09:16李瑞婷李东华牛玉芳陈冰洁王燕星何利洋姬海港王文韬刘树立康相涛李转见
中国畜禽种业 2023年12期
李瑞婷,李东华,牛玉芳,陈冰洁,王燕星,刘 阳,何利洋,姬海港,王文韬,刘树立,康相涛,李转见
(河南农业大学动物科技学院,河南郑州 450002)
家禽是动物蛋白的主要来源,尤其是鸡肉。家禽不仅提供了宝贵的膳食蛋白质供应,而且是发展中国家农村地区的重要收入来源[1]。鉴定与重要经济性状相关的遗传标记是提高鸡生产力和质量的基础,通过挖掘与经济性状相关的遗传标记和基因,我们可以更好地了解鸡的遗传背景,科学家们一直致力于对鸡的经济性状进行遗传标记挖掘和基因组选择。未来,随着技术的不断发展和研究的深入,鸡经济性状遗传标记挖掘与基因组选择将持续取得新的进展,并为畜牧业的可持续发展做出更大的贡献。
1 DNA 分子标记技术概述
分子标记技术是用DNA 序列上的突变来对动物表型进行标记的技术[2]。用该技术进行检测时,不会受个体性别、年龄和环境的影响,还可以标记较多的位点,提供丰富的遗传信息量,并且该技术具有高度的试验重复性,能够产生稳定和可靠的结果[3,4]。经过几十年的快速发展,分子标记技术已经趋于成熟,通过该技术,研究人员能够准确地定位基因,克隆感兴趣的基因,并在遗传育种中进行精确的选择和改良。
在遗传育种领域,DNA 分子标记主要包括:单核苷酸多态性(Single Nucleotide Polymorphism,SNPs)、插入/ 缺失多态性(Insertiondeletion,Indel)、拷贝数变异(Copy Number Variations,CNVs),这些分析标记主要通过以下几种方法进行检测(表1)。
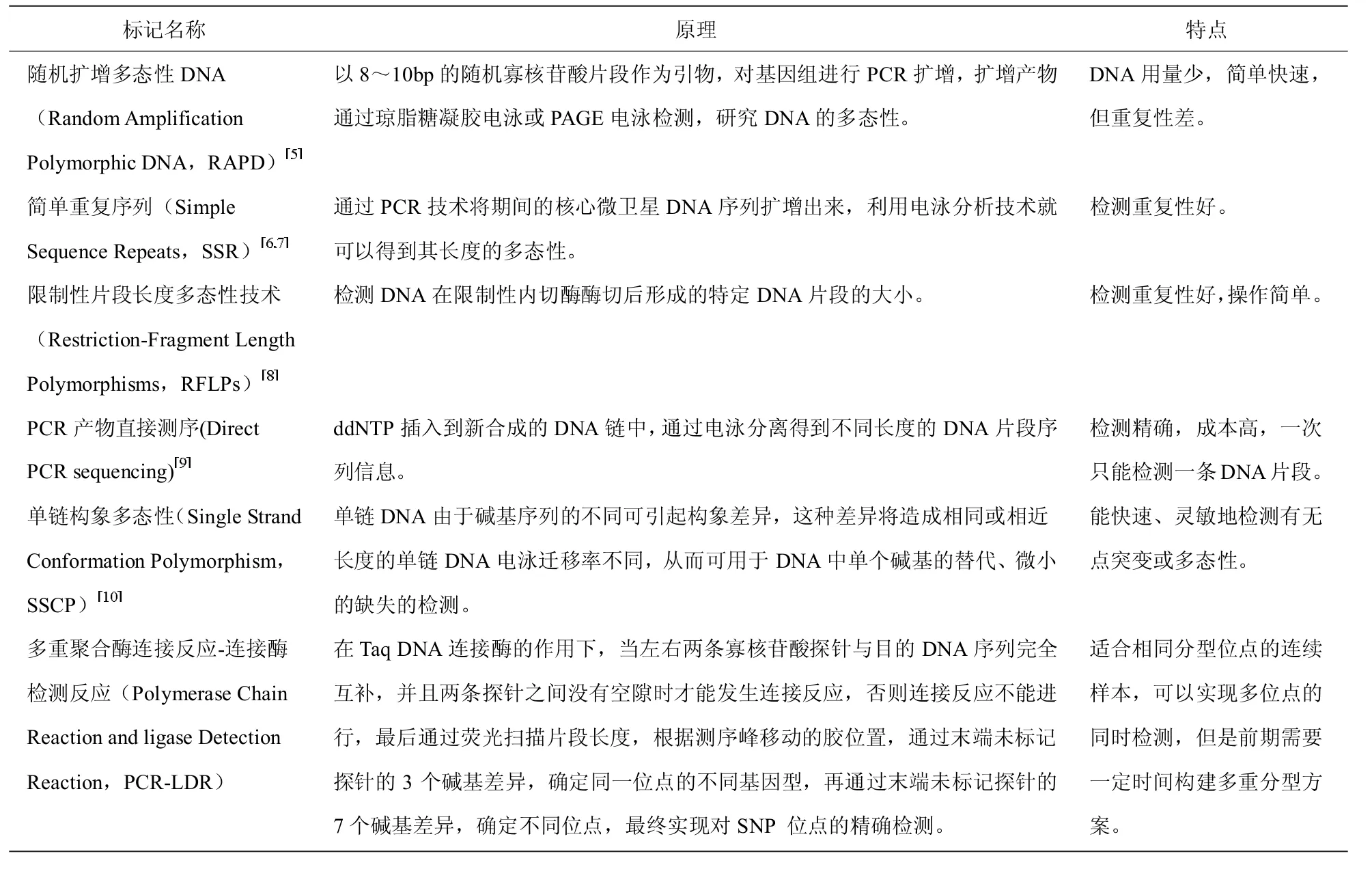
表1 常见的DNA 分子标记检测技术的原理和特点
2 DNA 分子标记技术在鸡经济性状上的应用
2.1 生长性状
研究鸡的生长性状对于育种改良、提高生产效率、增加经济效益和具有重要意义。生长性状主要包括体重、体斜长、胸骨长、胫骨长等多种表型,往往受到多基因的调控[11,12]。Yan 等[13]研究生长激素基因对生长和胴体性状的影响时,通过PCR-RFLP 发现与胴体性状(如胸肌重量、胸肌率、腹部脂肪率有显著关联的DNA 突变位点。杨菊凤[14]对不同周龄鸡的体重进行测定,并对10个DNA 位点进行SSR,筛选出控制体重的主效基因和位点。王强利用PCR-SSCP 技术胰岛素样生长因子-Ⅰ基因中找到了一些与生长发育、体重、龙骨长和胫骨长显著相关的位点。
现在很多研究已经通过DNA 分子标记技术挖掘出与鸡生长性状相关的基因。SH3 domain containing ring finger 2(SH3RF2)基因中CNV 与体重、胸骨长、半净膛重、全净膛重、胸肌重、肌胃重等生长发育指标极显著相关[15]。通过重测序 在Y-box 蛋 白3(Y-box binding protein 3,YBX3)基因的内含子区发现了一个27bp 的插入/缺失突变与鸡的半净膛重、全净膛重、胸肌重、腿重、腿肌重等屠宰性状显著相关[16]。细胞周期蛋白依赖性激酶抑制剂3(cyclin dependent kinase inhibitor 3,CDKN3)基因启动子区indel 突变的多态性与体重、胫长、腿重等生长发育指标显著相关[17]。硫酸肝素-6-O-磺基转移酶3(heparan sulfate 6-o-sulfotransferase 3,HS6ST3)基因上发现了与生长性状、胴体性状和肉质性状相关的43-bp indel 多态性[18]。另外还有很多其他indel 与鸡的生长性状相关,比如谷氨酰胺酰肽环转酶(glutaminyl-peptide cyclotransferase,QPCT)上有52 和224bp 的indel[19],锌指蛋白764-like (Zinc finger protein 764-like,ZNF764L)的第一个外显子区域有一个22bp 的indel[20],钾离子内向整流通道蛋白J 亚单位11 号成员(Potassium voltage-gated channel subfamily J member 11,KCNJ11)基因下游区域有一个新的163bp 的indel[21],催乳素受体(Prolactin receptor,PRLR)上存在80bp 的indel[22],羧基酯脂肪酶(Carboxyl ester lipase,CEL)上存在99bp 的indel[23],动情素受体基因(Motilin receptor,MLNR)基因下游区域有一个86bp 的indel[24]等。
2.2 繁殖性状
鸡的繁殖性能涵盖了受精率、孵化率、产蛋量和雏鸡存活率等指标。由于其经济重要性,越来越多与鸡产蛋性状的SSR 被挖掘出来[25],大部分数量性状基因座(Quantitative Trait Iocus,QTL)主要分布在在1 号,2 号和5 号染色体上[26]。例如Radwan[27]通过使用SSR 检测基因位点来改善埃及地方品种的产蛋量和产蛋质量性状。Su 等[28]通过PCR-LDR 研究与开产体重、开产蛋重和总产蛋量等显著相关的单核苷酸多态性的单倍型。常志伟[29]采用SSR 和RAPD 两种方法,对180 只渝西乌鸡慢羽系个体的基因组DNA 进行了分析,并将分析结果与试验鸡的产蛋性能进行了相关性分析,成功筛选出与产蛋率以及蛋重相关的分子标记。Makhsous[30]通过PCR 扩增和限制性核酸内切酶消化,用RFLP 方法筛选出与鸡产蛋率相关的分子标记。
2.3 肉质性状
鸡肉是人类营养的主要来源之一,目前禽肉的产量和消费量超过了猪肉,预计在未来10 年,禽肉将占新增肉类产量的近一半,所以研究肉质性状具有很高的经济价值。肉质评价的技术指标有pH 值、脂肪含量、剪切力、氧化稳定性等。Deeb 和Lamont[31]在肉鸡、来航蛋鸡及其F2 代杂交鸡种用SSR 检测到了过氧化物酶体增殖物激活受体α 上的有影响鸡脂肪代谢的QTL 位点。刘琛等[32]采用PCR-SSCP 技术在北京油鸡中筛选出与鸡肉肌内脂肪和肌苷酸含量相关的SNP 位点。李维等[33]采用PCR 产物直接测序技术检测赤水黑骨鸡肌肉生长抑制素基因多态性与肉质性状的关系,找到了可作为检测黑骨鸡的pH 值和剪切力分子标记。
2.4 抗病性状
鸡常见的疾病如禽流感、鸡白痢、马立克氏病等不仅会危及鸡的生命,更会严重影响养殖收益。马立克氏病是家禽业经济损失的主要原因,也是最严重和最持久的传染病问题。Heifetz 等[34]通过使用微卫星标记来鉴定影响MD 耐药性的QTL。有研究发现,RFLP 技术还可以鉴定对马立克氏病具有抗性和易感性的纯合雏鸡,大大提高了育种效率[35]。通过对抗病毒基因进行PCRRFLP 基因分型,发现该基因的cDNA 上有一个非同义多态性突变,而该突变已被证明会影响Mx 分子的抗病毒活性[36]。
3 基因组选择技术概述
一般来说,单个SNP 位点对鸡生长、产蛋等经济性状的影响可能性很小;因此需要大量的QTL 来解释这些性状的遗传变异[37]。鉴于这种遗传结构,仅使用少量DNA 标记来追踪数量有限的QTL 的DNA 分子标记技术的收益可能很小,这促进了使用密集SNP 标记的基因组信息来提高性状选择的准确性,通常也被称为基因组选择(Genomic Selection,GS)[38]。除了可以提高动物幼时选择的准确性外,基因组选择还有望降低每代近亲繁殖率,因为GS 提供了有关选择者的孟德尔遗传信息[39,40]。
基因组估计经济性状的育种值是基因组选择的核心,而基因组选择的实施包括两个主要步骤:首先需要建立一个参考群[42],利用在测试群体中测量的表型和标记基因型,开发基因组预测模型。其次,根据标记基因型预测的育种值,从相关的候选群体中挑选个体。贝叶斯[43]和基因组最佳线性无偏预测模型(Genomic Best Linear Unbiased Prediction,GBLUP)等方法[44,45]都能对QTL数据的基因组育种值进行预测。
4 鸡经济性状的基因组选择研究
随着越来越多测序技术的出现,全基因组重测序(Whole-Genome Sequencing,WGS)应用到GS 成为可能。由于WGS 数据包含大量基因组变异,其中包括与经济性状相关的生长、产蛋、肉质、抗病的基因突变,因此利用WGS 进行GS 有望获得更高的预测能力。使用WGS 比较低蛋壳强度和正常蛋壳强度的基因组,发现与蛋壳性状相关的SNP 和Indel,有助于在GS 时避免因蛋壳损伤导致的经济损失[46]。通过将五花黄鸡WGS数据和鸡参考基因组比较,成功发现在与鸡的代谢、免疫相关的基因上有众多SNP、Indel、结构变异和拷贝数变异,这个结果有助于我们更好地了解白羽肉鸡的遗传特征,并有助于未来通过GS 选育鸡生长、抗病性状[47]。对巴西F2 鸡资源群体进行WGS,获得的SNP 数据制作使用600 K高密度SNP 芯片,对来自鸡资源群体进行了基因分型,最终鉴定出与腹部脂肪和胴体脂肪含量性状的QTL,可以用来估计脂肪沉积相关性状的基因遗传率[48]。
全基因关联分析(Genome-Wide Association Studies,GWAS)是揭示基因组与表型组关系的重要工具,GWAS 结果不仅可以提高基因组预测的准确性[49],还能筛选出能用于GS 的SNPs 集[50]。已有多项针对鸡的生长[51,52]、体重[53]、胴体[54]、和肉质性状的GWAS 报告,这些结果不仅提供了许多可用于预测和基因组选择的分子标记,而且有助于解释这些性状的遗传机制。有学者利用鸡60k 高密度单核苷酸多态性阵列开展了一项GWAS,确定了与鸡细胞介导免疫相关的基因组区域,为后续利用GS 实现抗病性育种奠定了基础[55]。
目前针对鸡各种经济性状研发的芯片都可应用于GS。例如河南农业大学康相涛团队基于前期鸡泛基因组研究成果与公共数据,创制了40K、5K 地方鸡基因组系列芯片——“神农1 号”,囊括了生长、繁殖、屠体等性状GWAS 的功能位点,有利于地方鸡的种质资源鉴定、精准评价和品种保护[56]。根据典型地方品种和商业品系开发出的55K 基因组芯片上包括胸肌和肌内脂肪发育、体脂代谢、抗病等性状的SNPs 具有广泛的潜在应用价值,如基因组选育、经济性状的GWAS 分析以及不同鸡种品种多样性的研究[57]。
鸡群应用GS 后经济性状得到有效提升的例子有很多。有研究发现基于GS 选育出的禽类的体重、产蛋率、产蛋数等表型优于传统选育[58]。杂交育种被广泛用于提高植物和动物的生产率,有研究使用60K SNP 芯片进行基因分型,然后与传统的最佳线性无偏预测(Best Linear Unbiased Prediction,BLUP)方法和GS 中的GBLUP 等方法的预测能力和选择差异进行比较,发现GS 在鸡不同时期的体重、胸肌重、腿肌重、翅重和平均日增重等18 个性状上的准确性均高于传统BLUP[47]。此外,当利用GS 来缩短育种间隔时,还可以带来额外的遗传进展和经济效益[59]。基因组选择利用贝叶斯回归模型和半参数方法提升了肉鸡的饲料转化率预测的准确性[60]。使用GBLUP估算鸡新城疫病毒抗体反应和禽流感病毒抗体反应,发现基因组选择能提升这两个抗病性状的预测准确率,减少鸡群死亡率和农户经济损失[61]。选择和使用对沙门氏菌有更强抵抗力的鸡品系,可以减少沙门氏菌在表面健康的鸡身上的传播以及随之而来的食品安全问题[62]。
5 利用遗传标记与基因组选择应用的挑战
5.1 数据分析难度
一方面,处理大规模遗传标记数据的复杂性是GS 面临的一个挑战。随着高通量测序和SNP芯片技术的发展,可以获取大量的遗传标记数据,如全基因组测序和SNP 芯片数据。但处理和分析这些庞大的数据集是非常复杂的,涉及到数据预处理、统计分析、关联分析等。另一方面,考虑到遗传背景的复杂性、多基因性状的影响和环境因素,进行数据解读和解释也是一个挑战。
5.2 标记选择与验证
选择和验证具有预测和鉴别力的遗传标记也是一个关键的挑战。在标记选择方面,需要考虑标记覆盖范围、关联程度和遗传多样性等因素。通常使用关联分析和群体遗传学统计方法来筛选与经济性状相关的标记。然而,由于测序数据中的统计相关性和连接关系的复杂性,标记选择仍然存在一定的不确定性。在标记验证方面,可以使用功能基因组学方法,包括功能验证试验、表达谱分析和过量表达等来验证标记与经济性状之间的功能联系。此外,可以通过克隆和家系试验等种群遗传学方法来验证遗传标记的效果。
6 利用遗传标记与基因组选择应用的展望
为了提高种鸡选择的准确性和效率,可以利用遗传标记挖掘和基因组选择的方法,快速筛选出具有高经济效益的鸡种,并提高鸡群的产量、质量和适应力。首先,通过分析大规模的SNP 或基因测序数据,可以找到与经济性状密切相关的遗传标记,基于这些标记进行鸡种选择,可以更准确地预测鸡种的表现,提高鸡的经济性状和品质。其次,基因组选择可以加快鸡种选择的速度。通过分析标记与表型的关联性,可以快速评估个体的遗传优势,从而提前选出具有良好经济性状的个体作为父母本,大大缩短鸡种选择的周期,节约时间和资源成本。准确性和效率是鸡种选择中至关重要的因素,它们可以确保选择的鸡种具有良好的遗传潜力和经济性状,提高养殖业的盈利能力,还可以加快遗传改良的速度,适应市场需求的变化,以及更好地应对疾病和环境压力。
7 小结
利用DNA 分子标记挖掘和基因组选择,可以通过定位与经济性状密切相关的遗传标记,提高育种质量,加快育种速度,提高养殖业的收入。因此应该改善、提高和优化DNA 分子标记挖掘和基因组选择在鸡育种上的应用,克服技术上的困难,为养殖业发展提供保障。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
今日农业(2021年11期)2021-08-13 08:53:24
意林(2019年17期)2019-10-07 12:28:52
上海农业学报(2019年4期)2019-09-18 06:46:48
浙江农业学报(2017年4期)2017-04-26 02:28:22
浙江农业学报(2016年7期)2016-06-15 20:30:08
西南农业学报(2016年6期)2016-04-16 05:12:47
法医学杂志(2015年4期)2016-01-06 12:36:36
遗传(2014年3期)2014-02-28 20:58:49
世界科学(2014年8期)2014-02-28 14:58:31
