
构树源乳酸菌影响其青贮发酵品质
2024-01-11 10:40舒刚钦张云华程建波范彩云陈丽娟
草业科学 2023年12期
舒刚钦,王 慧,张 琛,张云华,张 玲,程建波,范彩云,陈丽娟
(1.安徽农业大学动物科技学院, 安徽 合肥 230036;2.安徽农业大学资源与环境学院, 安徽 合肥 230036)
构树(Broussonetia papyrifera)属于桑科植物,是一种极具开发价值的非常规林业蛋白饲料资源[1]。构树叶中粗蛋白含量丰富,最高可达干物质含量的20%~30%,富含多种氨基酸,其中谷氨酸、天冬氨酸和亮氨酸最为丰富,同时维生素、碳水化合物及微量元素等也颇为丰富。虽然构树枝叶作为营养价值很高的畜禽饲料,但是构树叶直接作为饲料时,蛋白质结构复杂,同时构树叶中的抗营养因子单宁会使蛋白质变性使得其难以被动物消化吸收,导致了高达80%以上营养物质会从粪便中排出,利用效率不高[2-5]。研究表明青贮可以改善构树叶营养价值,提高动物利用率[6-7]。付锦涛等[8]研究表明在全株构树中添加乳酸菌和糖蜜可改善发酵品质和营养成分。黄媛等[9]研究发现,在添加糖蜜的基础上单独或混合添加乳酸菌与纤维素酶能改善构树青贮饲料的发酵品质。刘晓婧等[10]研究表明,在杂交构树中添加乳酸菌可降低青贮pH,提高青贮品质。赵娜等[11]研究表明在构树叶青贮中增加乳酸菌含量可以提高青贮品质,但不同来源的乳酸菌青贮效果存在差异。
青贮是依靠乳酸菌厌氧发酵产生乳酸降低青贮饲料的pH,抑制腐败微生物和致病菌,达到长期保存植物饲料营养价值,提高植物饲料适口性和消化率的目的[12]。目前,植物青贮饲料中的乳酸菌主要有两个来源,一种是植物自身附着乳酸菌,另一种是人为添加的外源乳酸菌[13]。对于植物附着乳酸菌,一般都能适应其原青贮原料,但由于大部分青贮原料表面附着乳酸菌数量小于105,难以满足青贮发酵的要求,通常需添加外源乳酸菌以期迅速降低青贮原料的pH。但不同的青贮原料含有不同的微生物抑制因子,引起乳酸菌对青贮效果存在差异,导致青贮前期乳酸菌不能迅速扩增,青贮环境中pH 不能迅速降低而致使青贮品质下降[14-15]。构树中含有大量单宁,研究表明构树单宁对乳酸菌存在抑制作用,且不同来源的乳酸菌对单宁的耐受性存在差异[16],构树单宁是否影响青贮效果尚未明确。为此,本研究从自然构树青贮中分离乳酸菌,研究构树来源不同乳酸菌对构树叶青贮效果的影响,旨在为调制优质构树叶青贮饲料提供理论参考依据。
1 材料与方法
1.1 试验材料
本试验所采用的新鲜构树叶取自于安徽农业大学大杨店试验基地,新鲜构树叶干物质(dry matter,DM)含量为鲜重的30.15%,粗蛋白质(crude protein,CP)含量为干重的21.87%,中性洗涤纤维(neutral detergent fiber, NDF)含量为干重的28.35%,酸性洗涤纤维(acid detergent fiber, ADF)含量为干重的21.68%,可溶性碳水化合物(water soluble carbohydrate, WSC)含量为干重的6.15%。本试验使用构树单宁由实验室从构树中提取并保存。MRS 肉汤培养基购自海博生物(MRS 培养基,HB0384-1)。
1.2 试验方法
1.2.1 乳酸菌的分离
以自然青贮构树料作为菌株分离源,先从青贮罐中取10 g 构树叶,将其加入到装有90 mL 无菌生理盐水的锥形瓶(250 mL)中,并加入适量玻璃珠,封口膜封口后置于4 ℃冰箱中静置浸提24 h,摇匀后稀释成不同浓度涂布于MRS 固体培养基上并置于37 ℃厌氧培养箱中恒温培养。从上述培养平板中挑选出单菌落划线分离直至得到纯化菌株,随后对纯化菌株进行革兰氏染色、过氧化氢酶和葡萄糖产气试验[17],染色结果呈阳性并且过氧化氢酶接触试验呈阴性的菌株进行扩增并传代。-80 ℃甘油管保存菌种。
1.2.2 乳酸菌的形态学鉴定
观察MRS 固体培养基上单菌落的形态、大小、颜色,并进行镜检,观察菌株的细胞形态。分子鉴定:提取菌株的DNA、采用16S rDNA 通用引物(27F:AGAGTTTGATCCTGGCTC-AG;1492R:GGTTA CCTTGTTACGACTT)进行PCR 扩增。扩增成功后进行测序,在NCBI 上进行BLAST 序列同源性比对并构建系统进化树。
1.2.3 无菌单宁溶液的配制
将构树单宁高温灭菌,用无菌水配制成不同浓度无菌单宁溶液待用。
1.2.4 乳酸菌对构树单宁耐受性的测定
取对数生长期各乳酸菌菌液于4 ℃离心3 min,将沉淀菌体用无菌生理盐水洗涤3 次,然后用无菌生理盐水重悬,调整菌液浓度至OD600= 0.8,吸取上述菌液各2.5 mL 分别添加至单宁浓度为0.05、0.1和0.2 g L-1的50 mL 无菌构树单宁溶液当中,在转速 为150 r·min-1的 摇 床 中 作 用30 min 后 取10 μL菌液进行平板活菌计数,每个处理组重复3 次。
1.2.5 构树叶青贮调制
取对数生长期各乳酸菌菌液于4 ℃离心3 min,将沉淀菌体用无菌生理盐水洗涤3 次,然后用无菌生理盐水重悬,调菌浓度为1.0 × 109cfu·mL-1,作为青贮接种菌液。
将新鲜构树叶切碎至2~3 cm,混合均匀后作为青贮原料,设置5 个处理组,分别为对照组(PM0)、清酒乳杆菌组(PMQ)、戊糖片球菌组(PMW)、粪肠球菌组(PMF)与类魏斯氏杆菌组(PML),其中对照组添加5 mL 生理盐水,其他试验组分别添加5 mL 含对应乳酸菌菌液。混合均匀后将样品装入青贮发酵袋(30 cm × 40 cm)中,装袋量每袋500 g,真空封口机密封并使用真空泵抽真空,每个处理组重复3 次,在常温、避光条件下青贮60 d 后开封。
1.2.6 构树叶化学成分和发酵品质的测定
干物质、粗蛋白、中性和酸性洗涤纤维参照《饲料分析及饲料质量检测技术》[18]方法测定,可溶性碳水化合物采用苯酚-硫酸法测定[19]。准确称取20 g青贮样品,加入180 mL 无菌水于4 ℃浸提12 h 后,取浸提液测定发酵品质。pH 参考邵新庆等[20]方法,使用梅特勒FE28 型精密pH 计测定;氨态氮/总氮(NH3-N/TN)参考闫贵龙等[21]方法,采用苯酚-次氯酸钠法利用分光光度计测定;有机酸(乳酸、乙酸、丙酸、丁酸)含量采用高效液相色谱法测定[22]。
1.3 数据统计
试验数据采用Excel 2016 软件整理,利用SPSS 20.0 进行单因素方差分析,并用Duncan 氏法对各组数据进行多重比较,P< 0.05 表示差异显著,数据结果以“平均值 ± 标准差”表示。
2 结果与分析
2.1 构树青贮料中乳酸菌的分离及生理生化鉴定
构树青贮料中共分离出4 株乳酸菌,分别命名为R1、R2、R3、R4。细菌形态学和生理生化鉴定结果表明(表1),固体培养基上R1和R2菌落均呈乳黄色,圆形凸起,表面光滑湿润,R3和R4均呈乳白色,圆形,表面光滑湿润。革兰氏染色均为阳性,其中R1和R4为杆状,R2和R3为球状(图1)。经过氧化氢酶、甘油、明胶液化试验[23]上述菌株均为阴性。葡萄糖产气试验表明R1、R2和R3为阴性,而R4为阳性。通过糖发酵试验发现上述4 株菌均能利用麦芽糖、甘露醇、菊糖、蔗糖、乳糖、马尿酸和葡萄糖,R4不能利用纤维二糖和水杨苷,R1、R2和R3不能利用山梨醇,R3和R4不能利用棉籽糖。

图1 青贮构树分离菌株显微形态Figure 1 Micromorphology of the lactic acid bacteria isolated from Broussonetia papyrifera silage
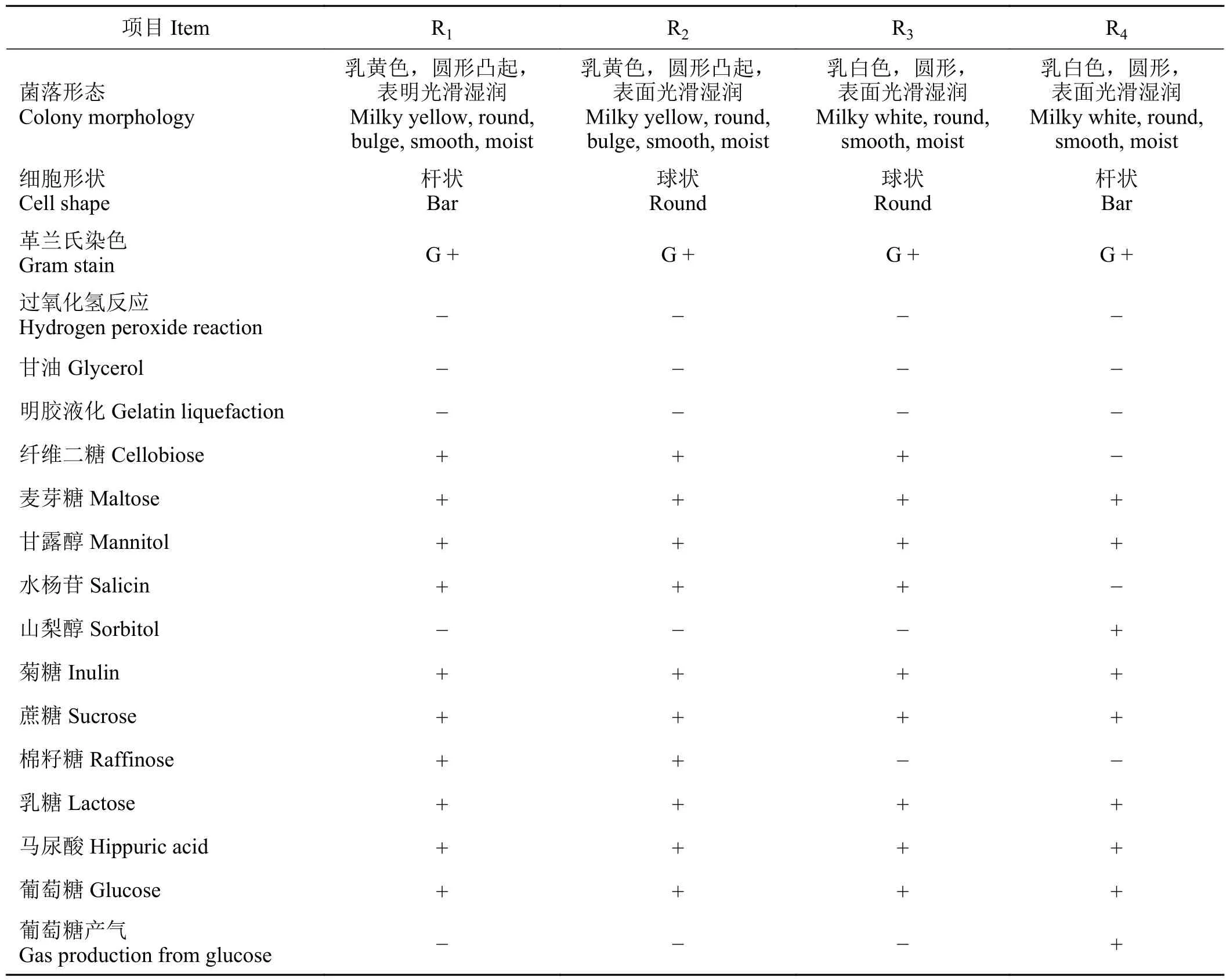
表1 构树青贮料中分离菌株形态及生理生化特性Table 1 Morphological, physiological, and biochemical identification of the strains from Broussonetia papyrifera silage
2.2 构树青贮料分离乳酸菌的分子生物学鉴定
将16S rDNA 序列结果与NCBI 数据库中序列数进行同源性比对发现,R1与清酒乳杆菌(Lactobacillus sakei)基因序列同源性为100%,R2与戊糖片球菌(Pediococcus pentosaceus)基因序列同源性为100%,R3与粪肠球菌(Enterococcus faecalis)基因序列同源性为100% ,R4与类肠膜魏斯氏菌(Weissella paramesenteroides)基因序列同源性为99.17% (图2)。结合生理生化鉴定结果确定构树枝叶中分离出4 株菌均为乳酸菌,其中R1为清酒乳杆菌,R2为戊糖片球菌,R3为粪肠球菌,R4为类肠膜魏斯氏菌。
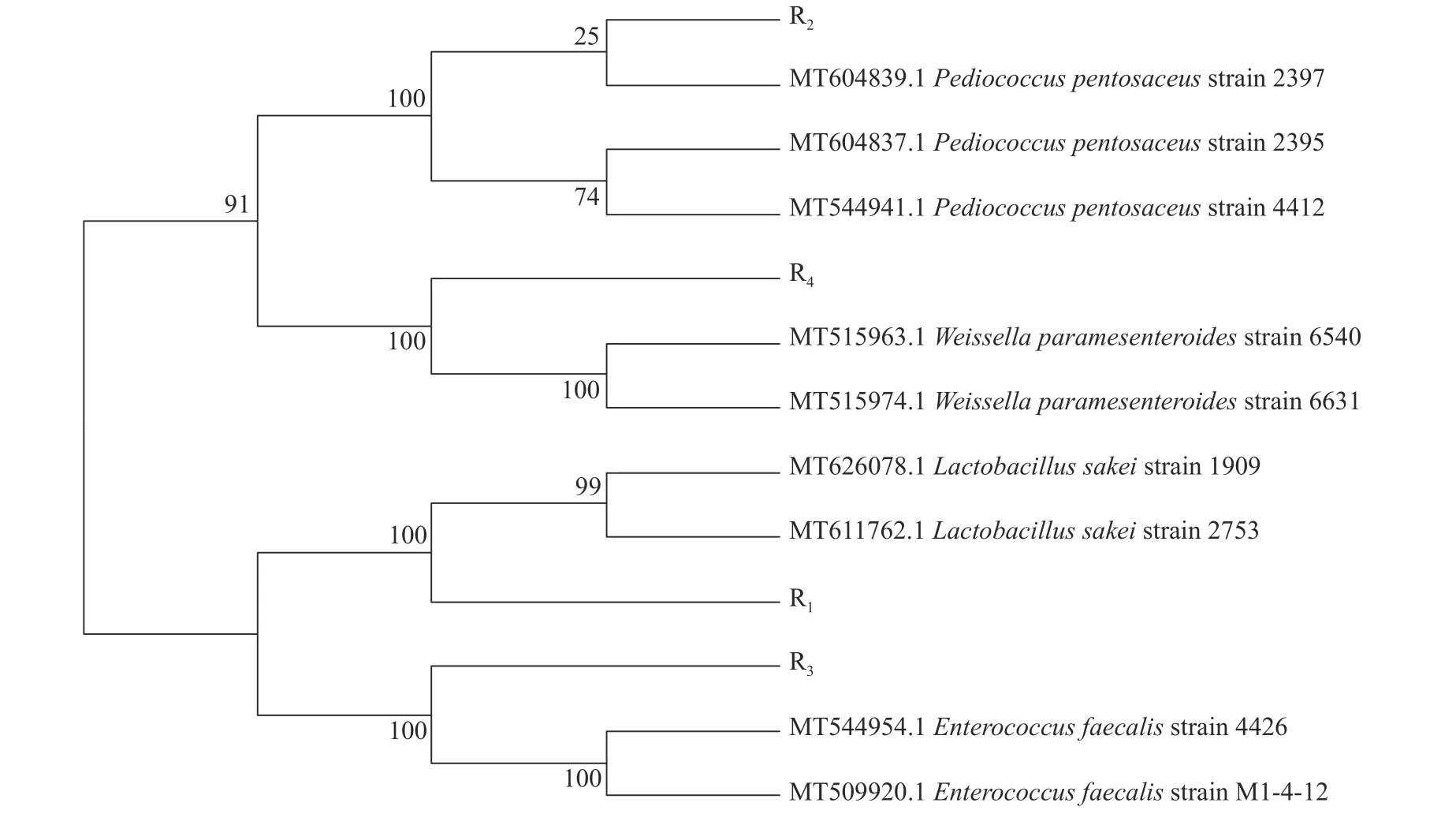
图2 青贮构树分离乳酸菌进化树Figure 2 Phylogenetic tree of the lactic acid bacteria isolated from Broussonetia papyrifera silage
2.3 构树青贮料中分离乳酸菌对构树单宁耐受性
构树青贮料中分离乳酸菌对构树单宁耐受性结果表明(表2),R2在同一浓度构树单宁作用下的存活率显著高于其他菌株(P< 0.05),R3显著高于R1和R4(P< 0.05),R4存活率最低;随着单宁浓度的增加,乳酸菌的存活率均显著降低(P< 0.05)。
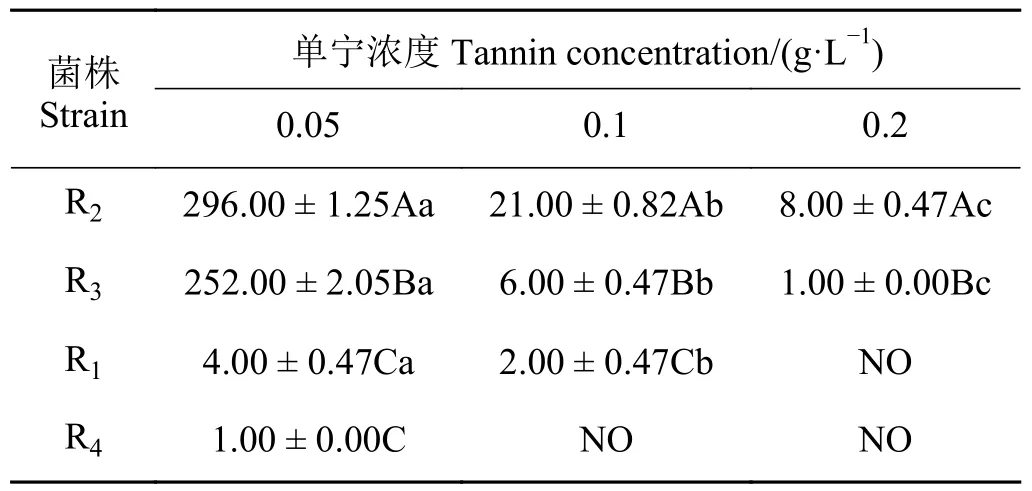
表2 乳酸菌对构树单宁耐受性Table 2 Tolerance of lactic acid bacteria to the Broussonetia papyrifera tannins cfu·mL-1
2.4 构树青贮料中分离乳酸菌对构树青贮营养水平的影响
构树青贮料分离乳酸菌青贮构树叶对其营养水平的影响结果表明(表3),对照组DM 含量显著低于各试验组(P< 0.05),各试验组间差异不显著(P> 0.05),但PMW 和PMF 组 均高于PMQ 和PML组;试验组的CP 含量均显著高于对照组(P< 0.05),PMW 和PMF 组显著高于PML 组(P< 0.05);PMQ、PMW 和PMF 组的NDF 含量显著高于对照组(P<0.05),各试验组之间差异不显著(P> 0.05);对照组ADF 显 著 高 于PMW 和PMF 组(P< 0.05),试 验组ADF 差异不显著(P> 0.05);各试验组WSC 含量差异不显著(P> 0.05),但均显著高于对照组(P< 0.05)。
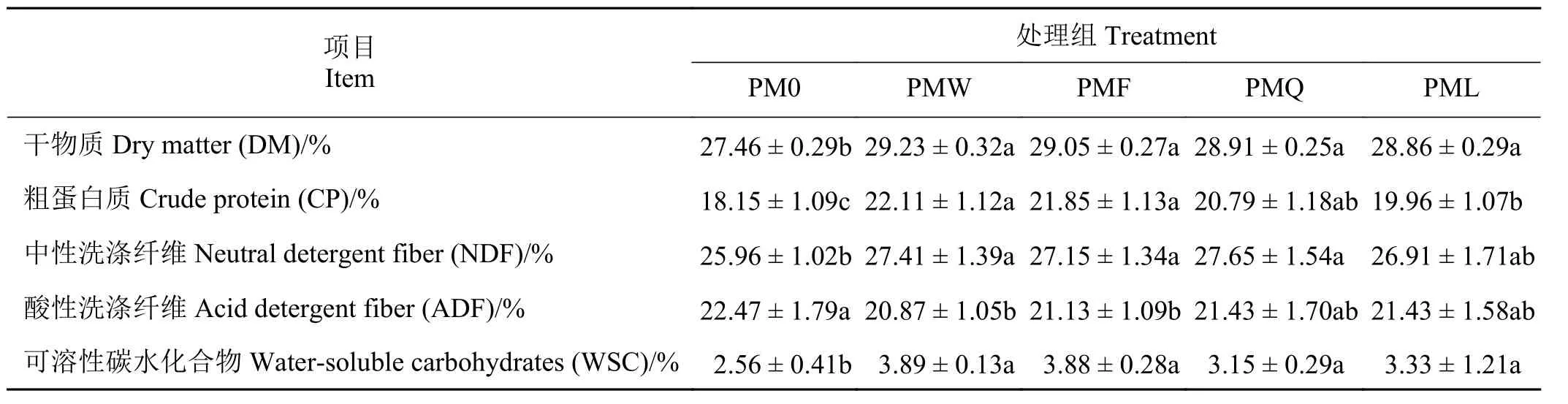
表3 青贮构树分离乳酸菌对构树叶青贮营养水平的影响Table 3 Effects of lactic acid bacteria on the nutrient levels of Broussonetia papyrifera leaves silage
2.5 青贮构树分离乳酸菌对构树叶青贮发酵品质的影响
青贮构树分离乳酸菌对构树叶青贮发酵品质影响结果表明(表4),添加乳酸菌试验组的pH 均显著低于对照组(P< 0.05),其中PMW 和PMF 组差异不显著(P> 0.05),PMQ 和PML 组差异不显著(P>0.05);所有试验组乳酸含量均显著高于对照组(P<0.05),试验组PMQ、PMW 和PMF 组的乳酸含量均显著高于PML 组(P< 0.05);PML 组乙酸含量显著高于其他试验组(P< 0.05);对照组的丙酸含量显著高于试验组(P< 0.05),PML 组丙酸含量显著高于其他试验组(P< 0.05);对照组和PMW 组检测到少量丁酸,其他各组均未检测出丁酸;对照组和PML 组氨态氮含量显著高于其他组(P< 0.05),PMW 和PMF 组氨态氮含量显著低于其他试验组(P< 0.05)。
3 讨论
3.1 青贮构树乳酸菌的分离与鉴定
乳酸菌在青贮发酵过程中起着重要的作用,其种类和数量对青贮发酵品质影响较大。制备青贮饲料的过程中必须具备数量多、活力强的乳酸菌,才能迅速降低pH,从而有效提高饲料青贮品质[24]。自然条件下,植物表面附着各种微生物,除了乳酸菌外,还包括厌氧的梭状芽孢杆菌属、兼性厌氧的泛菌属[25]、需氧的芽孢杆菌属、蓝细菌和大肠菌群[26-27]。好氧性细菌和大肠杆菌的数量多,而有益的乳酸菌含量较低。因而需通过添加外源乳酸菌加快发酵进程,迅速产生大量乳酸,使pH 下降,抑制有害微生物的活动,保存和提高青贮原料的营养价值。
已有研究表明不同青贮饲料中分离乳酸菌存在差异,郭刚等[28]从玉米(Zea may)秸秆青贮饲料中分离出粪肠球菌、植物乳杆菌和蒙氏肠球菌;李世丹等[29]从黑麦(Secale cereale)青贮饲料中分离得到植物乳杆菌、清酒乳杆菌、弯曲乳杆菌和屎肠球菌;刘晗璐[30]从禾本科牧草披碱草(Elymus dahuricus)和老芒麦(E.sibiricus)表面分离筛选出植物乳杆菌和蒙氏肠球菌;张敏[31]从西藏青贮饲料中分离出戊糖片球菌和乳酸片球菌。本研究从构树青贮饲料中共分离出清酒乳杆菌、戊糖片球菌、粪肠球菌、类肠膜魏斯氏菌4 种菌,与上述研究的植物中分离的乳酸菌存在一定差异,表明不同植物表面或青贮饲料中所分离出的乳酸菌种类差异较大。这种植物分离乳酸菌的差异与自然环境、植物种类、植物生育期、植物化学成分等有很大相关性[32-35]。
3.2 构树青贮料中分离乳酸菌对构树单宁耐受性差异的影响
单宁对微生物的抑制作用主要是因为单宁能与主要蛋白质如酶等形成氢键作用[36],植物单宁分为水解单宁和缩合单宁,其中水解单宁的分子量小于其缩合单宁[37],而单宁分子量大小决定其对微生物毒性的大小,一般小分子量单宁与蛋白质形成氢键,嵌入到微生物的活性部位,抑制微生物[38],但单宁聚合体与水解单宁相比,其对微生物的毒性反而降低[39]。上述理论预示同一单宁对不同微生物的抑制作用差异性可能不大,但本研究结果显示构树来源的不同乳酸菌对同一种构树单宁的耐受性差异较大,其中戊糖片球菌对单宁的耐受性能最强,这种差异可能与某些乳酸菌可以分泌特定单宁酶对目标单宁结进行降解,从而减少单宁对该乳酸菌的抑制作用[40],至于本研究中所获得的乳酸菌是否分泌单宁酶需要进一步验证。当单宁有一定关系同时随着构树单宁浓度的逐渐升高,乳酸菌对单宁的耐受性呈线性下降趋势。该结果表明不同微生物对由于其微生物蛋白的组成差异导致微生物对单宁的耐受性存在差异,从而影响微生物在生产中的活性,特别是青贮微生物浸入到含单宁的青贮原料中时,从而影响青贮效果。
3.3 青贮构树乳酸菌对构树青贮营养成分的影响
青贮料干物质含量是评定青贮饲料品质的重要因素,青贮料干物质的减少意味着原料青贮后营养成分的损失[41]。本研究中,对照组DM 含量显著低于试验组,表明试验组中构树源乳酸菌的添加有利于构树青贮DM 保存,减少其损失。CP 是饲料中重要营养物质,是衡量青贮饲料发酵品质的重要指标。研究发现,在饲料青贮过程中添加乳酸菌制剂有利于青贮料CP 保存[42-44]。在本研究中,添加乳酸菌的试验组CP 含量均高于对照组,主要是由于构树青贮发酵外源乳酸菌的添加,快速降低构树青贮发酵环境的pH,有利于抑制有害微生物的繁殖和对青贮料CP 的分解,从而减少其的损失[45-46]。同样是乳酸菌,添加粪肠球菌和戊糖片球菌组的CP 含量显著高于PMQ 和PML 组,这可能与粪肠球菌和戊糖片球菌对构树中单宁耐受性或其他化学成分的适应性有一定相关性,特别是青贮早期非构树来源植物乳酸菌繁殖速度难以适应青贮环境,导致CP 的损失。青贮前后NDF 的含量受青贮微生物的影响,已有研究表明参与NDF 降解的微生物主要是真核微生物,而细菌一般不参与NDF 的降解[47]。本研究发现对照组NDF 显著低于PMQ、PMW 和PMF组,表明青贮过程中有真核微生物参与了构树叶NDF 的降解。同时由于对照组的WSC 被有害微生物大量消耗,对照组中的微生物需要营养物质,如木霉属和曲霉属等能产生阿魏酸酯酶,促进半纤维素和木质素等水解,从而利用了NDF 中的结构性碳水化合物,进而导致NDF 降低[48],而试验组由于有外源乳酸菌的介入,加快青贮环境的pH 降低,减少NDF 降解。WSC 作为青贮发酵的底物,会被乳酸菌分解产生乳酸,从而使青贮饲料的pH 降低[49]。试验组的WSC 含量显著高于对照组,这可能是因为在青贮前期对照组的好氧微生物大量繁殖,消耗了大量的WSC,而试验组由于乳酸菌的添加导致有害微生物的减少,从而减少了WSC 的损失。
3.4 青贮构树乳酸菌对构树青贮发酵品质的影响
pH 和乳酸含量是衡量青贮饲料发酵品质的重要指标。由于低pH 可以抑制有害微生物,所以一般青贮饲料的品质越好,其pH 越低[50-51]。本研究结果表明,各试验组pH 均显著低于对照组,表明构树源乳酸菌的添加有利于构树叶青贮环境pH 降低,其中添加戊糖片球菌和粪肠球菌的PMW 和PMF组pH 显著低于PMQ 和PML 组,这种现象表明前两种乳酸菌更能适应构树青贮环境,有利于其青贮初期迅速增殖。本研究中,所测的各处理组青贮后的pH 均在4.3 以上,高于常规牧草青贮时获得的优质品质的pH 4.2,这是由于构树叶WSC 含量低,蛋白含量高,缓冲能值较高,限制构树叶的发酵,不利于pH 降低[52-54],但结合不同乳酸菌对构树单宁的耐受性结果表明,构树单宁对乳酸菌的影响与构树青贮pH 降低效果存在相关性。乳酸菌在青贮中通过产生大量有机酸来降低青贮环境pH,有机酸主要包括乳酸、乙酸、丙酸等。本研究发现添加构树来源的类肠膜魏斯氏菌的PML 组和对照组乳酸含量显著低于其他试验组,这可能与类魏斯氏杆菌属于异型发酵乳酸菌,在青贮中会将乳酸菌分解产生乙酸有关[55],PML 中的乙酸含量显著高于其他组也证明了这个结果。各试验组丙酸含量均显著低于对照组,表明试验组外源乳酸菌降低了发酵环境的pH,抑制了有害微生物的增殖[56]。丁酸是由腐败菌分解蛋白质、乳酸所产生的,丁酸含量越低,饲料的发酵品质越好[57]。本研究中对照组和PMW 组检测出少量丁酸,表明富集后的构树源乳酸菌添加可降低构树青贮料受有害微生物污染的机率。氨态氮是反映青贮过程中蛋白质降解程度的指标,其含量越高表明青贮过程中蛋白质降解越多,发酵品质越差[58]。本研究中,PMF 和PMW 组的氨态氮含量显著低于对照组,表明构树来源粪肠球菌和戊糖片球菌有利于构树青贮,避免青贮过程中蛋白质被梭菌等有害微生物过多降解[59]。
4 结论
本研究从青贮构树中共分离出4 种乳酸菌,其中R1为清酒乳杆菌(Lactobacillus sakei),R2为戊糖片 球 菌(Pediococcus pentosaceus),R3为 粪 肠 球菌(Enterococcus faecalis),R4为 类 肠 膜 魏 斯 氏菌(Weissella paramesenteroides),这4 种构树来源乳酸菌分别青贮构树叶均有利于构树叶青贮品质提高,但构树源不同乳酸菌对构树单宁耐受性不同,同时对构树叶青贮品质影响存在差异,本研究中戊糖片球菌和粪肠球菌更有利于构树青贮品质提高。
猜你喜欢
现代畜牧科技(2021年8期)2021-10-13
——“单宁”
山东国资(2020年6期)2020-07-09
绿色中国(2019年16期)2019-11-26
湖南饲料(2019年5期)2019-10-15
中国林业产业(2019年3期)2019-03-25
猪业科学(2018年4期)2018-05-19
中国扶贫(2017年16期)2017-08-16
西南学林(2016年2期)2016-11-08
应用化工(2014年11期)2014-08-16
当代畜禽养殖业(2014年10期)2014-02-27
