
湖南省湘潭市花榈木野生种群资源调查
2024-01-10 03:49:48周洁尘朱天才肖亚琴吴红强李学信
湖南林业科技 2023年6期
周洁尘,朱天才,肖亚琴,吴红强,段 翔,李学信,彭 皎,方 亮,文 虹
(1.湘潭市林业科学研究所,湖南 湘潭 411206;2. 湖南省林业科学院,湖南 长沙 410004)
花榈木(Ormosiahenryi)为豆目(Fabales)蝶形花科(Papilionaceae)红豆属(Ormosia)常绿乔木,又名花梨木、红豆树,主要分布在我国安徽、浙江、福建及两湖两广等亚热带地区,越南和泰国等国也有分布[1-2],集材用、景观和药用多种价值于一身[3],因其具有很高的利用价值,被大量采伐利用。花榈木种子种皮覆有蜡质,坚硬致密,透水、透气性较差,休眠期长,不易发芽,在自然传播繁殖情况下,种子1~2 a才能萌发,自然更新能力弱[4-5],致使现存野生资源稀缺[6],被列为国家二级重点保护植物,纳入《国家储备林树种目录》重要资源树种[7],世界自然保护联盟评估其濒危等级为易危(VU)。珍稀植物是生物多样性不可或缺的组成部分,随着花榈木野生资源的日益减少[8-9],花榈木野生种群的合理保护与可持续利用问题逐渐引起人们的重视[10-12]。
本研究通过走访、实地调查等方式,对湘潭地区花榈木野生种群资源状况进行详细调查,分析其自然种群地理分布、林龄结构及种群分布格局,以期为花榈木的保育、可持续开发与利用及今后的深入研究提供参考。
1 研究区概况
调查区域位于湖南省中部偏东地区湘潭境内,地理坐标为111°58′—113°05′E,27°21′—28°05′N;北连宁乡市、望城区、长沙县,南与衡东县、衡山县、株洲县接壤,东接株洲城区、株洲县,西与双峰县、涟源县交界,总面积5015km2。属亚热带季风性湿润气候,年平均日照为1507.7~1570.5h,年平均温度在16.9~17.4℃,平均无霜期269~281d;年均降水量1389.4~1437.8mm,年均蒸发量900~1000mm,年平均降雪日10d。植被主要是常绿阔叶次生林和马尾松(Pinusmassoniana)林、湿地松(Pinuselliottii)林、火炬松(Pinustaeda)林、杉木(Cunninghamialanceolata)林、油茶(Camelliaoleifera)林、毛竹(Phyllostachysheterocycla)林和灌木林等。
2 研究方法
>于2021年6月—2022年12月,对湘潭市区、湘潭县、湘乡市和韶山市可能有花榈木野生种群的地区,通过走访调查、实地调查等方式进行详细地调查。
2.1 走访调查
广泛走访林区农民、乡镇林业站工作人员、林场技术员等,了解到可能有花榈木野生种群的分布点、分布范围等,进行信息处理和资料汇总后,确定野外实际调查区域方案。
2.2 调查路线
根据初步走访,对分布点位进行统计,按照确定的调查区域逐一进行实地踏查。对花榈木野生种群采用实测法进行调查统计,确定湘潭地区花榈木野生种群的分布点,对其详细信息进行记录和整理。
2.3 实地调查
在野生花榈木分布点进行每木调查,调查统计采用实地踏查、形态观测、树体测量等方式进行[13]。详细记录花榈木每株的树高、冠幅、胸径及其种群所处的生境信息,包括分布点的群落组成、经纬度、海拔、郁闭度、坡度、坡向、土壤、干扰情况和保护管理现状等数据[14-15]。参考《中国植物志》[16]对种群分布点植物物种进行判定。
2.4 数据分析
2.4.1 分布图制定
根据调查发现的花榈木经纬度坐标应用ArcGIS 10.2软件投影到1∶10000地形图,编制花榈木种群点位分布图。
2.4.2 特定时间生命表编制
以胸径径级代替年龄级[17],将花榈木进行以下5个龄级的划分:Ⅰ级幼苗(D<2.5cm);Ⅱ级小树(2.5cm≤D<5cm);Ⅲ级中树(5 cm≤D<10cm);Ⅳ级成年树(10cm≤D<20cm);Ⅴ级老树(D≥20cm)。根据特定时间生命表编制的原理方法,计算生命表的各项参数,编制特定时间生命表。
生命表各特征值及其定义:
存活个体的百分数:
lx=nx/n1
(1)
式中:x为年龄级;nx为存活个体数。
x到x+1龄级间隔期内的死亡率:
qx=dx/nx
(2)
式中:dx为从x到x+1龄级间隔期内死亡个体数。
x到x+1龄级平均存活个体数:
Lx=(nx+nx+1)/2
(3)
从x龄级至更高龄级的平均个体总数:
Tx=∑Lx
(4)
x龄级开始时的平均生命期望值:
ex=Tx/nx
(5)
2.4.3 存活曲线编制
以花榈木各龄级为横坐标,标准化存活数lg(Lx)为纵坐标,制定存活曲线。数据采用Office 2010软件整理并绘制图表。
3 结果与分析
经野外实地调查,花榈木野生种群调查结果见表1。

表1 花榈木野生种群基本信息Tab.1 Basic information of wild populations of Ormosia henryi编号调查点E/(°)N/(°)海拔/m平均胸径/cmD≥2.5 cmD<2.5 cm株数/株株数/株总株数/株1湘潭县古塘桥乡天白村谢家湾112.833 35827.726 23839.6220.601012湘潭县石潭镇洪旺村槽门屋场112.775 13027.724 83269.754.55417213湘潭县石潭镇同庆村羊古塘112.752 44927.734 82391.724.241221334湘潭县石潭镇同庆村羊古塘112.749 70027.735 70075.007.6376125湘潭县石潭镇同庆村羊古塘112.749 00027.734 80071.8513.101786湘潭县分水乡长丰村王家山112.573 60027.559 10065.859.77128207湘潭县分水乡大冲垅村麦子坪112.604 00027.540 80060.3924.05230328湘潭县梅林桥镇黄竹村红旗组112.943 10027.669 30054.2612.62613199湘潭县梅林桥镇黄竹村天心组112.698 10027.913 50049.586.167172510湘乡市白田镇仁厚村老屋里112.364 11027.944 275138.1613.880811湘乡市白田镇仁厚村老屋里112.363 70027.944 400110.0010.276142012湘乡市白田镇仁厚村老屋里112.364 20027.944 300124.0014.93318118413湘乡市白田镇上麓村玄武冲112.366 81127.880 62571.9224.820214湘乡市白田镇白田村沙坪组112.402 44927.929 45771.4224.1020215湘乡市白田镇白田村桐子坪112.404 66027.918 59099.48 9.25 20216韶山市清溪镇永义村枫林冲112.553 70027.884 80045.3947.83101合计74316390
3.1 空间分布点位
从图1中可知,花榈木野生种群在湘潭地区不同市县的分布格局不一致。本次调查共发现花榈木野生种群分布点16个,大部分属于零散分布。根据湘潭地区地理位置划分,位于湘潭县的分布点有9个,位于湘乡市的分布点有6个,位于湘潭市的分布点为0个,位于韶山市的分布点有1个。湘潭市区没有野生花榈木,可能与其城市建设有关,在城建过程中,没有对野生种群进行保护。根据分布点所处位置,所有分布点均位于海拔35~140m之间,位于海拔35~60m的有4个,位于海拔60~110m的有8个,位于海拔110~140m的有2个(表1)。
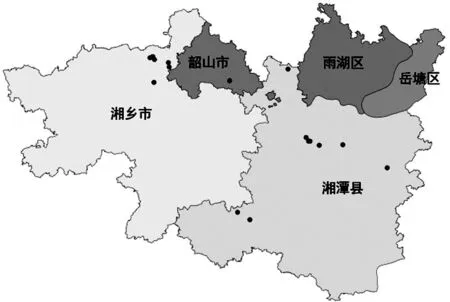
图1 湘潭市花榈木野生种群分布图Fig.1 The profile of wild populations of Ormosia henryi in Xiangtan City
3.2 林龄结构
分析年龄结构能有效探索种群发展和动态。根据花榈木生活史特点,通过实地调查,共发现野生花榈木390株,其中老树(D≥20cm)10株,成年树(10cm≤D<20cm)21株,中树(5cm≤D<10cm)22株,小树(2.5cm≤D<5cm)24株,幼苗(D<2.5cm)313株。有5个分布点无幼苗存在,最多的1个分布点也仅有幼苗181株,种群自然更新能力不足。从分布区域来看,位于湘乡市的种群个体数量也最多,其中幼苗数量最多。调查研究发现,绝大多数的野生花榈木只作为群落伴生种存在,能形成优势种的分布点极少;在开荒、采集、种植等人为活动强度较大的区域,花榈木种群在其分布点中数量较少,而且各成长期的个体数不均,尤其是幼苗在发育过程中极易死亡。
3.3 特定时间生命表
从表2可知,各龄级个体数都小于后一龄级个体数,死亡率为正;随着龄级增加死亡率先减小后增大;Ⅰ龄级死亡率达0.923,说明种群前期生存力极低,存活率低。
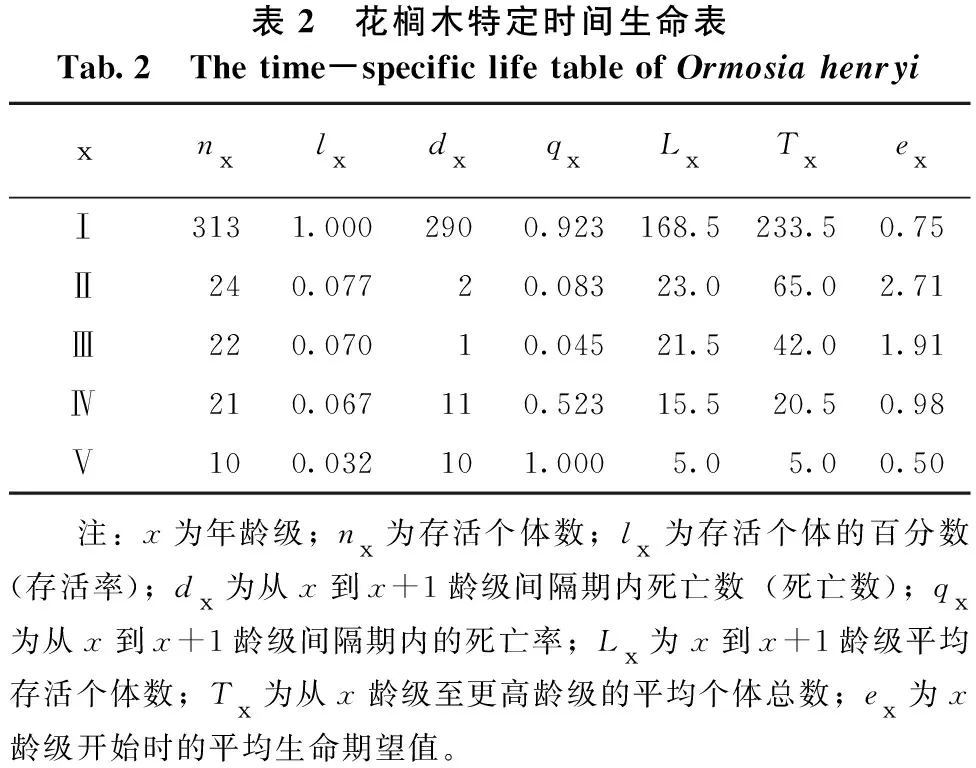
表2 花榈木特定时间生命表Tab.2 The time-specific life table of Ormosia henryixnxlxdxqxLxTxexⅠ3131.0002900.923168.5233.50.75Ⅱ240.07720.08323.065.02.71Ⅲ220.07010.04521.542.01.91Ⅳ210.067110.52315.520.50.98Ⅴ100.032101.0005.05.00.50 注:x为年龄级;nx为存活个体数;lx为存活个体的百分数(存活率);dx为从x到x+1龄级间隔期内死亡数(死亡数);qx为从x到x+1龄级间隔期内的死亡率;Lx为x到x+1龄级平均存活个体数;Tx为从x龄级至更高龄级的平均个体总数;ex为x龄级开始时的平均生命期望值。
生命期望值能在一定程度上体现种群个体生存能力强弱,从表2中期望寿命(ex)一栏中可以看出,湘潭地区整体花榈木种群的ex值均较低,存在一定相差值。ex期望值表现出随着龄级的增加,先上升到2.71后又下降,说明种群个体在持续发展过程中,生存不稳定。幼苗的数量较多,但生命期望值比较低;小树、中树、成年树数量较少且相对稳定,其中小树生命期望值最高,表现出较强的生存能力;老树的数量很少,且生命期望值较低。这表明花榈木种群具有幼苗期生长不稳定,幼龄树和中龄树稳定,老龄树衰退的特点。
3.4 存活曲线
从图2中可以看出,调查区域内的花榈木存活率最高在Ⅰ龄级,之后存活率随着龄级的增加而下降,表明幼苗数量虽相对较多,但存活到下龄级的生存力较低,幼苗期死亡率较高,个体存活数量急骤减少。整体上存活曲线接近于Deevey-Ⅲ型,表示种群生长发育过程中早期死亡率很高,但生活到一定的年龄阶段,死亡率变得较低而且稳定。
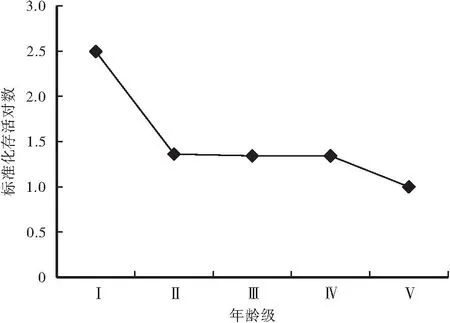
图2 花榈木种群存活曲线图Fig.2 Survival curve chart of wild populations of Ormosia henryi
3.5 群落结构
湘潭地区花榈木野生种群分布点树种组成丰富,成层明显,可分为乔木层、灌木层和草本层,层间植物较少,多样性较高,优势物种不明显,常绿物种数量较多,落叶阔叶成分并存,属常绿落叶阔叶混交林。由表3可知:分布点调查到乔木层物种16种,其中常绿物种9种,落叶物种7种。乔木层主要树种为青冈(Cyclobalanopsisglauca)、樟树(Cinnamomumcamphora)、苦槠(Castanopsissclerophylla)、枫香(Liquidambarformosana)、山矾(Symplocossumuntia)、苦楝(Meliaazedarach)、石栎(Lithocarpusglaber)、马尾松(Pinusmassoniana)等,其树型饱满,枝繁叶茂,盖度较大。灌木层物种45种,主要由青冈(Cyclobalanopsisglauca)、油茶(Camelliaoleifera)、檵木(Lespedezabicolor)、胡枝子(Lespedezabicolor)、柃木(Euryajaponica)、刚竹(Phyllostachyssulphurea)、盐肤木(Rhuschinensis)等物种组成。草本层物种较少,仅5种,层高0.5m以下,主要为淡竹叶(Lophatherumgracile)、铁线蕨(Woodwardiajaponica)、麦冬(Ophiopogonjaponicus)等。

表3 花榈木野生种群分布点群落组成Tab.3 Distribution point community composition of wild populations of Ormosia henryi层次物种乔木层青冈(Cyclobalanopsis glauca)、樟树(Cinnamomum camphora)、苦槠(Castanopsis sclerophylla)、枫香(Liquidam-bar formosana)、山矾(Symplocos sumuntia)、苦楝(Melia azedarach)、马尾松(Pinus massoniana)、石栎(Litho-carpus glaber)、杉木(Cunninghamia lanceolata)、小叶栎(Quercus chenii)、黄檀(Dalbergia hupeana)、冬青(Ilex chinensis)、南酸枣(Choerospondias axillaris)、蓝果树(Nyssa sinensis)、白栎(Quercus fabri)、毛竹(Phyllostachys edulis)灌木层青冈(Cyclobalanopsis glauca)、油茶(Camellia oleifera)、檵木(Lespedeza bicolor)、胡枝子(Lespedeza bicolor)、柃木(Eurya japonica)、刚竹(Phyllostachys sulphurea)、杉木(Cunninghamia lanceolata)、苦槠(Castanopsis sclero-phylla)、枫香(Liquidambar formosana)、盐肤木(Rhus chinensis)、白栎(Quercus fabri Hance)、黄檀(Dalbergia hu-peana)、冬青(Ilex chinensis)、杨桐(Adinandra millettii)、棕榈(Trachycarpus fortunei)、吴茱萸(Tetradium ruti-carpum)、山橿(Lindera reflexa)、野蔷薇(Rosa multiflora)、荚蒾(Viburnum dilatatum)、木樨(Osmanthus fra-grans)、女贞(Ligustrum lucidum)、杜鹃(Rhododendron simsii)、槲栎(Quercus aliena)、秤星树(Ilex asprella)、栀子(Gardenia jasminoides)、虎刺(Damnacanthus indicus)、野漆(Toxicodendron succedaneum)、野茉莉(Styrax ja-ponicus)、六月雪(Serissa japonica)、楤木(Aralia chinensis)、山莓(Rubus corchorifolius)、南烛(Vaccinium bractea-tum)、乌药(Lindera aggregata)、山胡椒(Lindera glauca)、檫木(Sassafras tzumu)、菝葜(Smilax china)、土茯苓(Smilax glabra)、忍冬(Lonicera japonica)、海金沙(Lygodium japonicum)、南蛇藤(Celastrus orbiculatus)、中华猕猴桃(Actinidia chinensis)、络石藤(Trachelospermum jasminoides)、铁芒萁(Dicranopteris dichotoma)、络石(Tra-chelospermum jasminoides)、大青(Clerodendrum cyrtophyllum)草本层铁线蕨(Woodwardia japonica)、淡竹叶(Lophatherum gracile)、麦冬(Ophiopogon japonicus)、爬山虎 (Partheno-cissus tricuspidata)、异叶蛇葡萄(Ampelopsis heterophylla)
4 结论与讨论
(1)我国物种丰富,但诸多原因致使某些植物濒临灭绝。湖南省属于典型的亚热带季风性湿润气候,植物物种资源丰富,但许多珍稀易危植物存在数量少、分布散等特点,常有老百姓误伐,因此珍稀易危植物野生种群资源调查及保护迫在眉睫。对湘潭地区珍稀易危植物花榈木野生种群进行了系统地调查分析,结果表明湘潭地区花榈木野生种群分布较少,分布点集中在湘潭县、湘乡市等生境良好、林分郁闭度适中、林内透光率较好的林缘边、山坡、溪谷旁等退化的生境区域。城市化建设进程较快的岳塘区、雨湖区没有分布;韶山市仅有1处分布,为城市化建设过程中留存下来的大树,附近未见其他幼苗或小树。城市化建设进程可能很大程度上制约了花榈木野生种群的自然更新和扩散,造成野生资源大量被采挖或砍伐。
(2)湘潭地区发现野生花榈木数量仅有390株,其中313株为幼苗,小树仅24株。存活曲线接近于Deevey-Ⅲ型,种群生长发育过程中早期死亡率很高,幼苗存活到下龄级的生存力较低,存活数量急骤减少。小树的数量对种群自然更新具有极为重要的意义,野生花榈木分布点小树数量极为有限,大部分也是集中生长,急需对其进行移栽和保护;否则,对种群的更新和扩散有着严重的制约作用。
(3)从野生花榈木种群年龄结构、静态生命表和存活曲线来看,花榈木野生种群整体上仍处于一个较为稳定的状态,在现有条件下可维持种群的持续发展,但更新能力不强。野生花榈木主要分布形式为聚集分布,可能与其种子传播方式和生境变化有密切关系。花榈木果为荚果,每荚果种子8粒左右,近圆形,受重力作用,大部分花榈木种子成熟后自然散落在母树周围。在自然环境中,种子散布在母树周围,幼苗表现出更高的聚集强度。但随着林分的发展,由于光照、养分等生长因子的供给限制,弱小的更新苗逐渐被淘汰,导致花榈木野生种群聚集强度随年龄增长而减弱。
(4)湘潭地区花榈木野生种群群落树种组成丰富,为常绿落叶阔叶混交林。群落中乔木物种种类丰富但富集种少,林分郁闭度较高,林隙较小,灌木层和草本层分布不均;灌木层主要是由乔木层物种更新苗和一些生态适应性较强的广布种组成;草本层物种种类稀少,但同种株数较多。绝大多数的花榈木只作为群落伴生种存在,野生种群数量相当匮乏。调查发现,湘潭地区花榈木野生种群基本零星分布在林缘边、山坡、溪谷旁等退化的生境区域。因此,减少人为干扰的影响对花榈木野生种群的保护极为重要。
(5)近年来,随着林业部门加大保护力度,部分天然条件下萌生的花榈木幼树得以自然生长。但调查中也发现存在人为盗伐盗采现象,亟需加大对花榈木野生种群保护投入,宣传生态保护政策,将发现的野生种群分布点作为重点巡护和监测对象,进一步掌握其种群数量、生境变化情况,在加强保护的同时,开展花榈木人工培育,实施迁地保育,加大推广力度,不断扩大花榈木种群数量。
猜你喜欢
湘潮(上半月)(2022年8期)2022-12-12 03:45:40
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
今日农业(2021年19期)2021-11-27 00:45:49
河南农业大学学报(2020年2期)2020-05-22 02:56:30
音乐教育与创作(2019年10期)2019-12-26 06:59:50
山西林业科技(2019年1期)2019-05-27 12:24:52
活力(2019年21期)2019-04-01 12:16:10
流行色(2017年12期)2017-10-26 03:08:22
初中生(2017年2期)2017-02-07 01:46:31
湖南农业(2016年3期)2016-06-05 09:37:34
