
猪NARS2基因变异与硒都黑猪胴体和肉质性状的关联分析
2024-01-08 09:18乔同同张宇李梓芃乔木秦振洋彭先文吴俊静
湖北农业科学 2023年12期
关键词:关联分析
乔同同 张宇 李梓芃 乔木 秦振洋 彭先文 吴俊静

摘要:在硒都黑豬群体中发现猪NARS2基因第9内含子中存在g.101284 C>T突变位点,采用PCR产物直接测序法进行基因分型后,与23个胴体和肉质类性状表型进行关联分析。结果表明,NASR2基因g.101284 C>T位点极显著影响猪胴体皮厚(P<0.01),且显著影响猪肉眼肌面积、滴水损失48 h和肌内脂肪含量性状(P<0.05)。CC基因型个体皮厚极显著低于TT基因型个体(P<0.01);CC基因型个体滴水损失48 h和肌内脂肪含量显著高于其他基因型个体,而TT基因型个体胴体眼肌面积显著小于其他基因型个体(P<0.05)。而这4个性状中,除皮厚与眼肌面积性状不存在显著相关外,其他各性状之间存在一定程度的相关性。其中,滴水损失48 h与肌内脂肪含量性状相关性最高(r2=0.882,P<0.01)。因此,该标记在育种应用中应当结合育种目标合理利用,针对不同主选性状,选留或淘汰不同基因型个体。
关键词:硒都黑猪;胴体性状;肉质性状;NARS2基因;关联分析
中图分类号:S828;S813 文献标识码:A
文章编号:0439-8114(2023)12-0195-05
DOI:10.14088/j.cnki.issn0439-8114.2023.12.034 开放科学(资源服务)标识码(OSID):
Association analysis of NARS2 gene variation and carcass and meat quality traits
in selenium black pig
QIAO Tong-tong, ZHANG Yu, LI Zi-peng, QIAO Mu, QIN Zhen-yang, PENG Xian-wen, WU Jun-jing
(Institute of Animal Science and Veterinary Medicine, Hubei Key Laboratory of Animal Embryo and Molecular Breeding, Hubei Academy of Agricultural Sciences, Wuhan 430064,China)
Abstract: A g.101284 C>T mutation site was found in intron 9 of the NARS2 gene in the selenium black pig population. After genotyping using PCR product direct sequencing, association analysis was conducted with 23 phenotypes of carcass and meat quality traits.The results showed that the g.101284 C>T locus of the NASR2 gene significantly affected the skin thickness of pig carcasses (P<0.01), and significantly affected the eye muscle area, drip loss for 48 hours, and intramuscular fat content traits of pork (P<0.05). The skin thickness of individuals with CC genotype was significantly lower than that of individuals with TT genotype (P<0.01);the 48 hours drip loss and intramuscular fat content of individuals with CC genotype were significantly higher than those with other genotypes, while the carcass eye muscle area of individuals with TT genotype was significantly smaller than that of individuals with other genotypes (P<0.05). Among these four personality traits, there was a certain degree of correlation between other traits except for skin thickness and eye muscle area traits, which were not significantly correlated. Among them, the 48 hours drip loss had the highest correlation with intramuscular fat content traits (r2=0.882, P<0.01). Therefore, this marker should be reasonably utilized in breeding applications in conjunction with breeding objectives, and individuals of different genotypes should be selected or eliminated for different main selected traits.
Key words: selenium black pig; carcass traits; meat quality traits; NARS2 gene; association analysis
中国是全球猪肉生产大国,无论是养殖规模还是消费量都居于世界前列。胴体和肉质是猪育种行业中的重要经济特征,胴体质量及猪肉品质的高低直接影响着养殖场的生产和经济效益。胴体和肉质的大多数性状属于中高等遗传力[1-3],如肌内脂肪含量性状遗传力可达0.49~0.86[4]。目前,人们对影响肉质特征的关键基因了解仍然不足。因此,寻找控制猪肉质量的关键分子遗传标记,并将其应用于分子标记辅助育种,对于提高猪的肉质特征、增加经济效益具有重要意义。
线粒体转运核糖核酸合成酶2(NARS2)是一种酰基tRNA合成酶,其在细胞的蛋白质合成过程中起着重要作用。在人类相关研究报道中发现,NARS2基因变异在联合氧化磷酸化缺陷24(OXPD24)中引起神经退行性和肌病综合征[5,6],与非综合征性常染色体隐性耳聋94(DFNB94)[7]、利氏综合征[8]等疾病相关。而有关该基因在动物中的研究报道较少。
硒都黑猪是以恩施黑猪、梅山猪、湖北白猪等为基础,历经12年培育而成的黑猪新品种。课题组前期利用基因组重测序技术对比分析了硒都黑猪肌内脂肪含量极端差异个体的基因组群体分化信号,发现NARS2基因在高、低肌内脂肪组中存在显著分化位点。因此,本研究扩增了猪NARS2基因序列,利用该序列进行遗传变异位点的筛选鉴定及与胴体和肉质性状进行关联分析,以期获得与猪肉质性状相关的新分子育种标记。
1 材料与方法
1.1 样品采集
275头健康硒都黑猪饲养于湖北华健硒园农牧科技有限公司硒都黑猪育种基地,在体重达100 kg左右时,进行屠宰测定,采集背最长肌(背肌)组织黄豆大小样本,置于2 mL冻存管,液氮保存。同时收集并记录屠宰日龄、屠宰日期、屠宰体重、系谱等基本信息。随机选取少量个体,采集腰肌、比目鱼肌、心脏、肝脏、脾脏、肺脏、肾脏、背脂、腹脂、结肠、回肠、盲肠等组织黄豆大小样本,置于2 mL冻存管,液氮保存。
1.2 胴体和肉质性状检测
屠宰率、胴体长、平均背膘厚、皮厚、肋骨对数、眼肌面积、腿臀比例、皮率、肥肉率、瘦肉率、骨率等胴体性状按照国家行业标准NY/T 825—2004《瘦肉型猪胴体性状测定技术规范》规定方法测定。肉色评分1、肉色评分24、肌肉色值L1、肌肉色值L24、大理石纹评分、pH1、滴水损失48 h、系水力、肌内脂肪、水分、嫩度等肉质性状表型按照国家行业标准NY/T 821—2019《猪肉品质测定技术规程》规定方法测定。
1.3 基因组DNA和RNA提取
基因组DNA的提取采用北京百泰克生物技术有限公司生产的基因组DNA提取试剂盒(按该试剂盒说明书进行操作)提取;组织总RNA利用Invitrogen TRIzol和氯仿提取。通过1%琼脂糖凝胶电泳检测核酸质量,并用Nanodrop 2000型分光光度计检测核酸浓度。
1.4 猪NARS2基因荧光定量检测
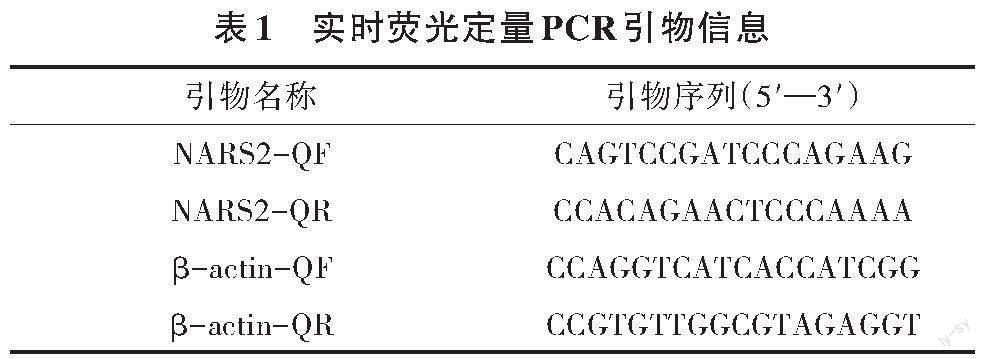
取1 μg的RNA样本,利用Thermo ScientificTM RevertAidTM First Strand cDNA Synthesis试剂盒反转录cDNA。采用THUNDERBIRD SYBR qPCR Mix试剂盒,使用QuantStudio 6荧光定量PCR仪(Applied Biosystems公司)进行qPCR检测。以β-actin基因为内参,采用2-ΔΔCt法计算相对表达量,每组试验做3个平行,每个试验重复3次。实时荧光定量PCR引物信息见表1。采用20 μL反应体系,反应程序为95 ℃预变性5 min,95 ℃变性15 s,56 ℃退火15 s,72 ℃延伸15 s,循环40次;熔解曲线:95 ℃ 15 s, 60 ℃ 1 min,96 ℃ 1 s。
1.5 猪NARS2基因多态性检测
以40头硒都黑猪的DNA样本混合池为模板,进行猪NARS2基因第9内含子片段序列PCR扩增,上游引物:5′-TGTGCATTTATGTGTATTTCGGGG-3′,下游引物:5′-GGTCAAATGACGATGCTGCT-3′。PCR反应体系(50 μL):100 ng模板DNA、10×Buffer (含Mg2+) 4 μL、上下游引物各0.5 μmol/L、2.5 μmol/L dNTPs、1 U Taq DNA聚合酶。PCR运行程序:95 ℃预变性2 min;94 ℃变性20 s,65~55 ℃退火40 s(每个循环降低0.5 ℃),72 ℃延伸60 s,共20个循环;94 ℃变性20 s,55 ℃退火30 s,72 ℃延伸60 s,共15个循环;72 ℃延伸10 min;4 ℃保存。PCR产物用1%琼脂糖凝胶电泳检测后,采用Gel Extraction Kit试剂盒进行纯化。纯化产物直接送到北京奥科公司进行DNA测序,对比筛选基因变异位点。
1.6 基因分型
利用上述引物分别扩增硒都黑猪DNA,PCR产物经Sanger测序分析后进行基因分型。
1.7 统计分析
采用SPSS 26.0统计软件中一般线性模型GLM进行基因型与性状表型值的关联分析。一般线性模型GLM的表達式如下:
Yijklm=μ+Gi+Aj+Xk+Sl+eijklm (1)
式中,Yijklm表示性状表型值;μ表示群体均值;Gi表示基因型效应;Aj表示年季效应;Xk表示性别效应;Sl表示父本效应;eijklm表示随机误差,结果以最小二乘均值±标准误表示。
利用SPSS 26.0统计软件中的相关分析模型计算表型性状的皮尔逊相关系数。P<0.05判定为显著差异,P<0.01判定为极显著差异。
2 结果与分析
2.1 猪NARS2基因组织表达谱
利用qPCR方法检测了猪NARS2基因组织表达谱情况,发现该基因在肌肉、脂肪、各内脏器官等组织中广泛表达,在肝脏组织中表达量最高,其次是背肌、腰肌、比目鱼肌等肌肉组织,在盲肠组织中表达量最低(图1)。
2.2 猪NARS2基因多态性检测结果
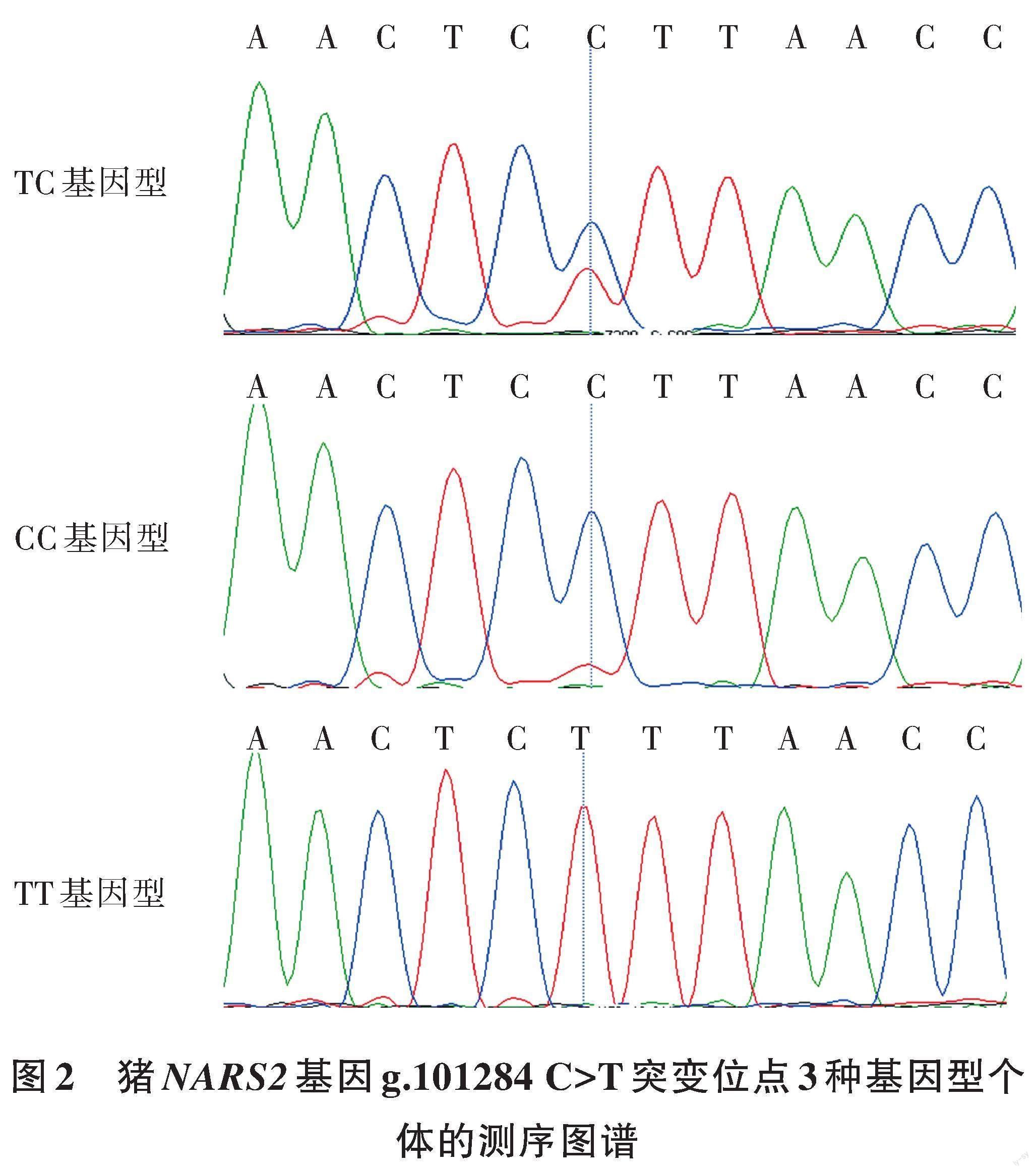
通过对40个硒都黑猪DNA混合样本池进行NARS2基因片段扩增测序,发现猪NARS2基因第9内含子中存在1个单碱基突变位点,即位于该基因序列第101 359 bp处的C>T碱基突变(g.101284 C>T),如图2所示。
2.3 猪NARS2基因变异与胴体和肉质性状的关联分析
将275头硒都黑猪NARS2基因g.101284 C>T基因型与其胴体和肉质等23个表型性状进行关联分析,结果如表2所示。NASR2基因g.101284 C>T位点极显著影响猪胴体皮厚,CC基因型个体皮厚极显著低于TT基因型个体(P<0.01)。同时,该位点还显著影响猪肉眼肌面积、滴水损失48 h和肌内脂肪含量性状,CC基因型个体48 h滴水损失和肌内脂肪含量显著高于其他基因型个体,而TT基因型个体胴体眼肌面积显著小于其他基因型个体(P<0.05)。
2.4 显著相关性状表型值相关性分析
由于猪NARS2基因g.101284 C>T突变显著影响多个胴体和肉质性状,因此将该位点显著影响的皮厚、眼肌面积、滴水损失48 h、肌内脂肪含量4个性状的表型值相关性进行进一步分析,结果如表3所示。滴水损失48 h和肌内脂肪含量分别与皮厚存在显著负相关(P<0.05);滴水损失48 h和肌内脂肪含量分别与眼肌面积存在极显著负相关(P<0.01);滴水损失48 h与肌内脂肪含量存在极显著正相关(P<0.01)。
3 小结与讨论
氨基酰化反应主要由氨基酰化tRNA合成酶家族酶催化[9],参与生物蛋白质合成。NARS2最先在酵母中被发现[10],属于II类氨基酰基-tRNA合成酶,是一种由细胞核编码,但是可以到达线粒体发挥功能的线粒体型天冬酰胺酰tRNA合成酶[11]。研究发现NARS2基因突变可导致tRNA天冬氨酸水平降低,线粒体复合物I、III和IV的活性受损,并对成纤维细胞的耗氧率产生负面影响[12]。在人类研究中,发现该基因突变与婴儿癫痫发作、生长发育迟缓、智力发育迟缓、肌张力低下和听力障碍、肾脏疾病或肝脏异常等病例相关[6,12-14]。在动物中,有关NARS2基因的研究较少,本研究探讨了猪NARS2基因对性状表型的影响。发现猪NARS2基因在猪的各内脏器官、肌肉、脂肪组织中广泛表达,与其广泛参与蛋白质合成过程有关。在硒都黑猪群体中,猪NARS2基因外显子区域并未找到关键突变位点,这也可能是因为外显子突变对蛋白质表达影响较大,会导致个体发育不正常,因而在群体选育中逐渐被自然淘汰。
内含子序列占哺乳动物基因组的24%,虽然其不编码蛋白质,但与编码序列一起转录,共剪接形成成熟mRNA后,内含子才与mRNA脱离[15]。内含子作为pre-mRNA被剪接后的产物,具有复杂多样的生物学功能。例如,内含子参与可变剪接的调控,可变剪接增加了蛋白质的多样性[16,17];可通过提高转录速率、介导增强效应、介导mRNA降解、提高mRNA翻译速率等途径促进基因表达[15]。有研究证实内含子变异会导致表型性状发生变化,例如,人类研究报道发现内含子变异与苯丙酮尿症、癌症等疾病密切相关[18-20];湖羊STAT5a基因第10内含子变异影响泌乳性状[21];松辽黑猪KPNA7基因内含子变异影响繁殖性状[22]。
本研究在猪NASR2基因第9内含子中鉴定到1个单碱基突变位点g.101284 C>T,其显著影响皮厚、猪肉眼肌面积、滴水损失48 h和肌内脂肪含量4个胴体和肉质性状。而这4个性状中,除皮厚与眼肌面积性状不存在显著相关外,其他各性状之间存在一定程度的相关性,有的是正相关,而有的是负相关。其中滴水损失48 h与肌内脂肪含量性状相关性最高(r2=0.882,P<0.01)。g.101284 C>T位点对不同性状影响趋势存在一定差异。例如,CC基因型个体皮厚极显著低于TT基因型个体(P<0.01);CC基因型个体滴水损失48 h和肌内脂肪含量显著高于其他基因型个体,而TT基因型个体胴体眼肌面积显著小于其他基因型个体(P<0.05)。因此,该标记在育种应用中应结合该育种群体的既定育种目标及主选性状合理利用。例如,若要选择眼肌面积大且肌内脂肪含量高的群体,应予以保留携带CC基因型个体;若要选择滴水损失48 h小的群体,应淘汰携带CC基因型的个体。
综上,本研究鉴定了猪NASR2基因第9内含子中一个显著影响猪胴体和肉质性状的新育种标记,今后将对其具体作用机制开展更深入的研究。
参考文献:
[1] 陈 辉.猪重要经济性状选择的遗传进展[J].猪业科学, 2019, 36(1):116-118.
[2] 刘 航,侯黎明,王彬彬,等.显性效应对苏淮猪肉色性状遗传评估和基因组选择的影响[J].畜牧与兽医, 2021, 53(6):1-6.
[3] 廖印长, 张跃博, 何 俊.猪肉质性状全基因组关联分析的研究进展[J].畜牧与兽医, 2022, 54(8):125-131.
[4] 陈艳珍.猪肉品质的评定及影響因素[J].中国畜牧兽医, 2012, 39(7):155-158.
[5] VAFAEE-SHAHI M,FARHADI M,RAZMARA E,et al. Novel phenotype and genotype spectrum of NARS2 and literature review of previous mutations[J]. Irish journal of medical science,2022,191(4):1877-1890.
[6] ZHANG Y, ZHAO X, XU Y, et al. Study of novel NARS2 variants in patient of combined oxidative phosphorylation deficiency 24[J]. Translational pediatrics, 2022, 11: 448-457.
[7] AL-SHARIF F, ALSADEQ H, ROZAN A, et al. Bilateral nonsyndromic sensorineural hearing loss caused by a NARS2 mutation[J]. Cureus, 2022, 14(11): e31467.
[8] TANAKA R, TAKEGUCHI R, KURODA M, et al. Novel NARS2 variant causing leigh syndrome with normal lactate levels[J]. Human genome variation, 2022, 9(1):12.
[9] WEI N, ZHAGN Q, YANG X L. Neurodegenerative charcot-marie-tooth disease as a case study to decipher novel functions of aminoacyl-tRNA synthetases[J]. Journal of biological chemistry, 2019, 294(14):5321-5339.
[10] LANDRIEU I, VANDENBOL M, HARTLEIN M, et al. Mitochondrial asparaginyl-tRNA synthetase is encoded by the yeast nuclear gene YCR24c[J]. European journal of biochemistry, 1997, 243: 268-273.
[11] HU W, FANG H, PENG Y, et al. Clinical and genetic analyses of premature mitochondrial encephalopathy with epilepsia partialis continua caused by novel biallelic NARS2 mutations[J]. Frontiers in neuroscience, 2022, 16:1076183.
[12] SIMON M, RICHARD E M, WANG X, et al. Mutations of human NARS2, encoding the mitochondrial asparaginyl-tRNA synthetase, cause nonsyndromic deafness and leigh syndrome[J]. PLoS genetics, 2015, 11(3): e1005097.
[13] SOFOU K, KOLLBERG G, HOLMSTROM M, et al. Whole exome sequencing reveals mutations in NARS2 and PARS2, encoding the mitochondrial asparaginyl-tRNA synthetase and prolyl-tRNA synthetase, in patients with alpers syndrome[J]. Molecular genetics & genomic medicine, 2015, 3:59-68.
[14] MIZUGUCHI T,NAKASHIMA M,KATO M,et al. PARS2 and NARS2 mutations in infantile-onset neurodegenerative disorder[J]. Journal of human genetics, 2017, 62:525-529.
[15] 曹艷娟.不同表达水平基因内含子与mRNA序列的匹配特征[D].呼和浩特:内蒙古农业大学, 2023.
[16] OLSON S, BLANCHETTE M, PARK J, et al. A regulator of dscam mutually exclusive splicing fidelity [J]. Nature structural & molecular biology, 2007, 14(12): 1134-1140.
[17] WANG Z, BURGE C B. Splicing regulation: From a parts list of regulatory elements to an integrated splicing code[J]. RNA, 2008, 14(5): 802-813.
[18] 张 钏.甘肃地区遗传代谢病的遗传图谱及PAH基因深部内含子变异致病性研究[D].北京:北京协和医学院, 2022.
[19] SCIARRILLO R, WOJTUSZKIEWICZ A, ASSARAF Y G, et al. The role of alternative splicing in cancer: From oncogenesis to drug resistance [J]. Drug resistance updates, 2020, 53: 100728.
[20] PADGETT R A. New connections between splicing and human disease[J]. Trends in genetics, 2012, 28(4): 147-154.
[21] 刘 玉, 张林林, 房 义,等.湖羊STAT5a基因第10内含子多态性及其与泌乳性状的关联分析[J].中国畜牧兽医, 2023, 50(9):3680-3687.
[22] 张方玮, 张 琪, 张云鹏,等.松辽黑猪KPNA7基因多态性及其与繁殖性状的关联分析[J].中国畜牧杂志, 2023, 59(9): 235-239.
猜你喜欢
网络空间安全(2017年1期)2017-03-10
江苏农业科学(2017年1期)2017-02-27
档案管理(2017年2期)2017-02-25
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
计算机应用(2016年12期)2017-01-13
软件导刊(2016年11期)2016-12-22
科技视界(2016年15期)2016-06-30
珠江水运(2016年10期)2016-06-20
电脑知识与技术(2016年4期)2016-04-11
