
干旱胁迫下云南干热河谷区6个乡土草种生理生化特性的比较分析*
2024-01-05 11:34:54刘爱林伏春美罗中阳常恩福周云李品荣庞静
西部林业科学 2023年6期
刘爱林,伏春美,罗中阳,常恩福,周云,李品荣,庞静
(1.云南农业大学 动物科学技术学院,云南 昆明 650201;2.云南省林业和草原科学院,云南 昆明 650201;3.云南省高黎贡山生物多样性重点实验室,云南 昆明 650201;4.高黎贡山森林生态系统云南省野外科学观测研究站,云南 保山 678000)
干旱是全球面临的主要环境挑战之一,严重威胁着农业、生态系统和人类社会稳定。干旱是一种生理形式的水分亏缺,土壤可利用水分减少,对植物的代谢产生不利影响。叶片相对含水量是反映胁迫下植物水分有效性的指标[1]。植物一般进化出生理调节和生化反应等适应机制来应对干旱胁迫[2]。干旱对植物的影响表现在植物的生长、膜完整性、色素含量和渗透调节水分关系等[3-4]。有研究认为,丙二醛、脯氨酸、可溶性糖、叶绿素等可作为植物抗旱性生理评价的重要指标[5]。此外,干旱胁迫通常会导致活性氧(ROS)的增加,包括过氧化氢、超氧离子和羟基自由基[6]。高浓度的ROS会破坏植物细胞内的化合物,如蛋白质、细胞膜等[7]。因此,为维持干旱胁迫下细胞ROS稳态,植物进化出酶促抗氧化防御系统,从而缓解胁迫对植物造成的损害。主要的植物ROS清除酶包括超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)等[8]。因此,探究植物应对水分亏缺产生的生理生化反应对植物适应干旱环境尤为重要。
云南干热河谷是我国西南典型的生态脆弱地区,干旱作为当地重要的环境胁迫因素,严重制约着植物生长和发育[9]。乡土草种是干热河谷生态系统中的重要组成部分,具有很强的抗逆能力和适应性。李建查等[10]通过盆栽模拟自然干旱,分析扭黄茅(Heteropogoncontortus)、孔颖草(Bothriochloapertusa)、拟金茅(Eulaliopsisbinata)和百喜草(Paspalumnotatum)的土壤水分变化和植株苗期抗旱生理性状,探讨干热河谷优势草本植物的抗旱生理特性。赵琳等[11]对干热河谷的车桑子(Dodonaeaviscosa)、清香木(Pistaciaweinmannifolia)两个原生植物的抗旱性能进行系统综合评价。为筛选干热河谷地区优良耐旱草种,本试验以分布于金沙江干热河谷区的芸香草(Cymbopogondistans)、水蔗(Apludamutica)、茅叶荩草(Arthraxonprionodes)、黄花猪屎豆(Crotalariapallida)、红毛草(Rhynchelytrumrepens)、三芒草(Aristidaadscensionis)为试验材料,通过测定其生物学特性指标和生理生化特性指标,比较分析6个草种的抗旱能力,旨在为培育适合干热河谷种植的草种并以之用于生态修复提供理论依据。
1 材料与方法
1.1 供试材料
本试验所选用的6个乡土草种为黄花猪屎豆、红毛草、三芒草、芸香草、水蔗、茅叶荩草,均采自云南省楚雄彝族自治州永仁县莲池乡小尖山(101°37′43″E、 26°3′6″N,海拔1 743.4 m)。
1.2 试验设计
草种盆栽试验于2022年8月11日,在云南农业大学动物科学技术学院草学实验室进行。花盆内直径30 cm,高25 cm。采集干热河谷燥红土,风干,过2 mm筛,装盆,每盆装土6 kg。在每个盆里均匀播10粒饱满的种子,每个草种播种15盆。植株长出2~3片真叶时(播种出苗后第15 d)定株,每盆挑选5株长势一致的幼苗。对6个草种进行持续浇水,使花盆中的土壤水分处于饱和状态,保证其初始土壤含水量为田间持水量的75%~80%以上。待植株生长到4~5叶龄(播种出苗后第30 d)时,停止浇水,于9月10日开始进行自然干旱试验(干旱胁迫共持续37 d)。
参照植物水分梯度划分方法[12],设置4个水分梯度:正常水分(75%~80%以上)、轻度干旱(60%~65%)、中度干旱(40%~50%)、重度干旱(25%~35%),每个处理3个重复。在重度干旱后进行复水,复水时使土壤含水量恢复至对照水平(田间持水量的75%~80%以上),持续3 d后进行采样。土壤含水量于每天早上8:00—11:00使用土壤水分测定仪(MPM-160,ICT International Pey.Ltd.)测定。当土壤含水量降低至每个水分梯度时,每个草种从中取3盆植株进行采样。采样时每个处理都选取相同部位完全成熟的叶片,用锡纸迅速包裹并标记,液氮冷冻后放入-80 ℃低温冰箱保存以待进行各项指标的测定。
1.3 指标测定
用土壤水分测定仪测定土壤含水量;丙二醛(MDA)、可溶性糖(SS)、脯氨酸(PRO)、过氧化物酶(POD)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)采用苏州格锐思生物科技有限公司生产的试剂盒测定;叶片相对含水量(LRWC)采用称重法测定[13];叶绿素含量采用95%酒精浸泡法测定[14]。
1.4 抗旱性评价方法
抗旱性评价采用隶属函数综合评价法。以6种干热河谷优势草在不同程度干旱胁迫下的叶片相对含水量、脯氨酸、丙二醛、叶绿素、过氧化氢酶、过氧化物酶、超氧化物歧化酶、可溶性糖单项指标进行综合性评价。计算公式如下。
若所测指标与植物的耐旱性呈正相关。
F1=(Xi-Xmin)/(Xmax-Xmin)
若所测指标与植物的耐旱性呈负相关。
F2=〔1-(Xi-Xmin)/(Xmax-Xmin)〕
式中:Xi表示第i个指标值,Xmax表示所有品种第i个指标的最大值,Xmin表示所有品种第i个指标的最小值。
叶片相对含水量、脯氨酸、叶绿素、可溶性糖、过氧化物酶、超氧化物歧化酶、过氧化氢酶与抗旱性呈正相关,丙二醛与抗旱性呈负相关,根据以上公式得到6个草种各指标的具体隶属函数值并计算出其平均值后进行比较,平均值越大表示该草种的抗旱性越强[15]。
1.5 数据处理
采用Excel 2013进行数据处理;运用SPSS 19.0对6个草种在不同胁迫程度下的各指标进行单因素方差分析,Duncan法对其多重比较,独立样本T检验对同一胁迫程度下各指标进行差异性分析。
2 结果与分析
2.1 干旱胁迫对各草种叶片相对含水量的影响
由表1可看出,胁迫前,黄花猪屎豆叶片相对含水量显著高于其他5个草种(P<0.05),高出幅度为4.18%~13.86%,三芒草的叶片相对含水量最低。随干旱程度加剧,各草种叶片相对含水量均呈降低的趋势,各水分梯度间6个草种叶片相对含水量降幅分别为2.51%~7.56%、9.53%~23.52%和16.81%~31.94%。重度干旱时,各草种叶片相对含水量下降至最低值,此时茅叶荩草叶片相对含水量显著高于其他草种(P<0.05),三芒草叶片相对含水量最小。干旱条件下,三芒草叶片相对含水量降幅最大,为46.05%,茅叶荩草降幅最小,为31.29%。复水后,各草种叶片相对含水量有所上升,但未恢复至对照水平。

表1 干旱胁迫各时期草种叶片相对含水量变化Tab.1 Changes of leaf relative water contents of drought stress in different drought stress period %
2.2 干旱胁迫对各草种叶片脯氨酸含量的影响
由表2可见,正常水分及轻度干旱条件下,三芒草脯氨酸含量显著高于其他草种(P<0.05);中度干旱和重度干旱时,芸香草脯氨酸含量显著高于其他草种(P<0.05)。6个草种随干旱加剧其叶片脯氨酸含量呈升高趋势,芸香草、水蔗、茅叶荩草、红毛草4个草种在各水分梯度间显著升高;黄花猪屎豆与三芒草脯氨酸含量在轻度干旱时没有显著变化,之后显著升高。各草种脯氨酸含量在重度干旱时达到峰值,此时,芸香草脯氨酸含量升高了3.62倍,增幅最大,红毛草增幅最小,为0.99倍。复水后,各草种脯氨酸含量显著降低(P<0.05),其中,黄花猪屎豆与三芒草脯氨酸含量恢复至对照水平。

表2 干旱胁迫各时期草种叶片脯氨酸含量Tab.2 Proine content of grass varieties’leaves in different drought stress period μg/g
2.3 干旱胁迫对各草种叶片丙二醛含量的影响
由表3可看出,胁迫前,红毛草丙二醛含量与三芒草相近,平均为15.30 nmol/g,显著高于其他草种(P<0.05)。当草种遭受干旱胁迫时,随土壤水分下降,除茅叶荩草外其余5个草种丙二醛含量呈升高趋势。茅叶荩草丙二醛含量在轻度至中度干旱时,茅叶荩草丙二醛含量没有显著变化。芸香草丙二醛含量呈升高趋势,中度干旱后趋于稳定。各草种丙二醛含量在重度干旱时达到最大值,且显著高于对照(P<0.05)。此时,黄花猪屎豆丙二醛含量最大,为43.74 nmol/g,芸香草丙二醛含量最小,为29.55 nmol/g。就增幅而言,黄花猪屎豆丙二醛增幅最大,比对照升高了2.81倍,三芒草丙二醛增幅最小,比对照升高了1.20倍。复水后,各草种丙二醛含量均有所降低,但未恢复至对照水平。

表3 干旱胁迫各时期草种叶片丙二醛含量Tab.3 MDA content of grass varieties’leaves in different drought stress period nmol/g
2.4 干旱胁迫对各草种叶片叶绿素含量的影响
正常水分条件下,水蔗叶绿素含量最高,显著高于其他草种(P<0.05),茅叶荩草叶绿素含量最低(表4)。在干旱胁迫过程中,水蔗、茅叶荩草、三芒草叶绿素含量呈降低趋势,但在重度干旱时,三芒草叶绿素含量有所回升,比中度干旱高出幅度为17.76%。芸香草、黄花猪屎豆、红毛草叶绿素含量在轻度干旱时略有升高,其升高幅度为0.33%~0.45%,之后持续降低。除三芒草外,其他5个草种在重度干旱时叶绿素含量降至最小值,其中,红毛草叶绿素含量最高,为6.02 mg/g,茅叶荩草叶绿素含量最低,为1.67 mg/g。与对照相比,茅叶荩草叶绿素含量降幅最大,为60.98%,红毛草降幅最小,为17.53%。复水后,6个草种叶绿素含量均显著升高(P<0.05),红毛草与三芒草叶绿素含量恢复至对照水平。

表4 干旱胁迫各时期草种叶片叶绿素含量Tab.4 Chlorophyll content of grass varieties’leaves in different drought stress period mg/g
2.5 干旱胁迫对各草种叶片可溶性糖含量的影响
胁迫前,三芒草可溶性糖含量显著高于其他草种(表5)。随干旱程度加剧,芸香草、水蔗可溶性糖含量呈升高趋势。茅叶荩草、红毛草、三芒草在轻度干旱时,可溶性糖含量显著降低,其降低幅度分别为33.77%、20.43%和19.62%,之后持续升高。黄花猪屎豆可溶性糖含量整体呈升高趋势,在中度干旱时出现降低,其可溶性糖含量降低了0.95 mg/g,之后大幅升高,提高了1.83倍。至重度干旱时,6个草种可溶性糖含量达到最大值,三芒草可溶性糖含量最高,为23.84 mg/g,茅叶荩草可溶性糖含量最低,为11.37 mg/g。与对照相比,黄花猪屎豆增幅最大,升高了1.82倍,红毛草增幅最小,升高了0.30倍。复水后,可溶性糖含量均有所降低,其中,红毛草与三芒草外可溶性糖含量恢复至对照水平。
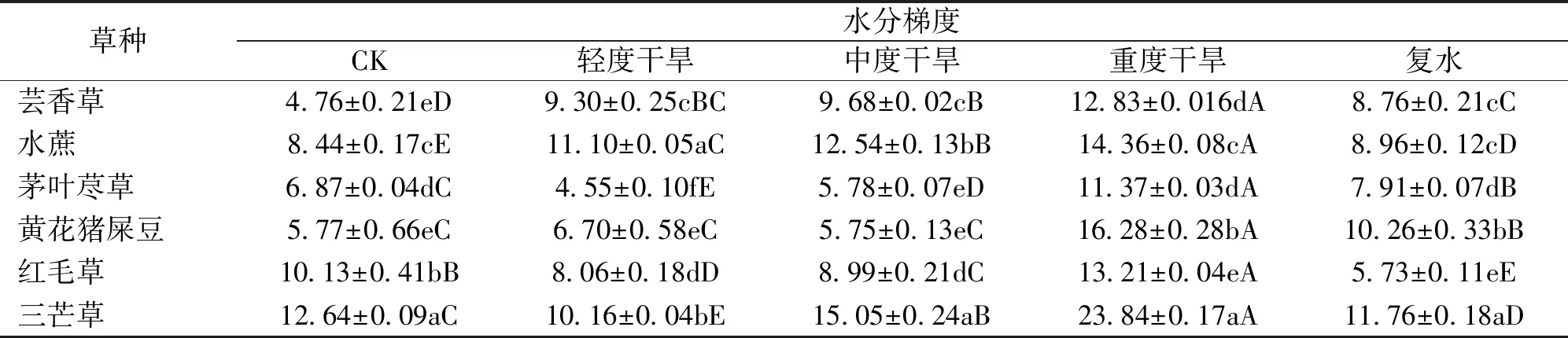
表5 干旱胁迫各时期草种叶片可溶性糖含量Tab.5 Soluble sugar content of grass varieties’leaves in different drought stress period mg/g
2.6 干旱胁迫对各草种叶片过氧化物酶活性的影响
正常水分条件下,茅叶荩草POD活性显著高于其他草种(P<0.05),而水蔗POD活性最低(表6)。胁迫后,水蔗、茅叶荩草POD活性先降低后升高,在轻度干旱时分别下降了17.42%、23.95%。芸香草、黄花猪屎豆、红毛草、三芒草POD活性在干旱胁迫下呈升高趋势,至重度干旱时POD活性达到最大值。其中,茅叶荩草POD活性最大,黄花猪屎豆POD活性最小。重度干旱时,水蔗增幅最大,比对照升高了2.81倍,黄花猪屎豆增幅最小,比对照升高了1.54倍。复水后,各草种过氧化物酶活性显著降低(P<0.05),且显著高于对照水平(P<0.05)。
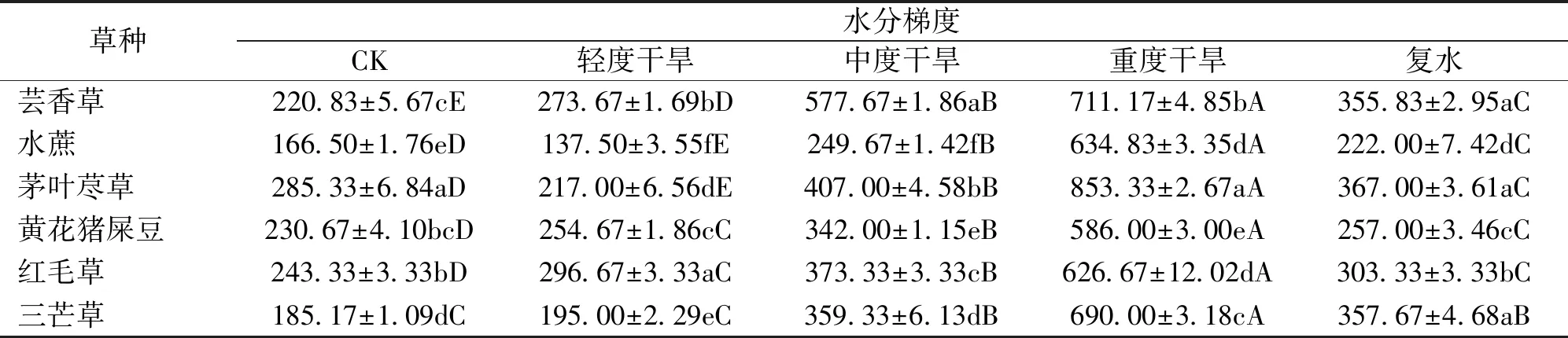
表6 干旱胁迫各时期草种叶片过氧化物酶活性Tab.6 Peroxidase activity of grass varieties’leaves in different drought stress period U/g
2.7 干旱胁迫对各草种叶片超氧化物歧化酶活性的影响
由表7可知,在干旱胁迫下,茅叶荩草SOD活性均显著高于其他草种(P<0.05)。随着干旱胁迫程度加剧,6个草种SOD活性显著升高(P<0.05)。重度干旱时SOD活性达到最大,茅叶荩草SOD活性值最大,为1 175.25 U/g,芸香草SOD活性值最小,为408.16 U/g。与对照相比,水蔗增幅最大,升高了3.91倍,黄花猪屎豆增幅最小,为1.67倍。复水后,6个草种SOD活性均有所降低,但未恢复至对照水平。
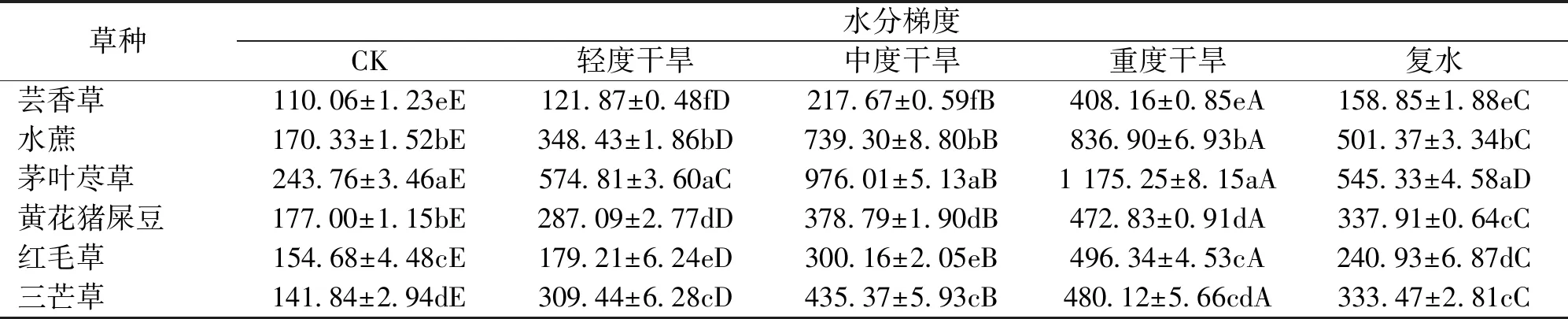
表7 干旱胁迫各时期草种叶片超氧化物歧化酶活性Tab.7 Superoxide dismutase activity of grass varieties’leaves in different drought stress period U/g
2.8 干旱胁迫对各草种叶片过氧化氢酶活性的影响
正常水分条件下,三芒草CAT活性显著高于其他草种(P<0.05),黄花猪屎豆CAT活性最小(表8)。受干旱胁迫影响,茅叶荩草、黄花猪屎豆、红毛草CAT活性呈升高趋势,黄花猪屎豆在中度至至重度干旱时CAT活性趋于稳定。水蔗与三芒草在轻度干旱时CAT活性显著降低(P<0.05),之后CAT活性持续升高。而芸香草在轻度、中度干旱时CAT活性显著升高(P<0.05),在重度干旱时CAT活性降低了63.63 U/g。重度干旱时,三芒草CAT活性最大,黄花猪屎豆CAT活性最小。此时三芒草增幅最小,仅升高了0.11倍,茅叶荩草CAT活性增幅最大,升高了0.91倍。复水后,6个草种CAT活性均显著降低(P<0.05),黄花猪屎豆CAT活性接近对照水平。

表8 干旱胁迫各时期草种叶片过氧化氢酶活性Tab.8 Catalase activity of grass varieties’leaves in different drought stress period U/g

表9 供试材料的抗旱性综合评价Tab.9 Comprehensive comparison of drought resistance of experimental materials
2.9 抗旱性综合评价
根据隶属函数公式[15]求出6个草种在干旱胁迫下生物学特性指标(叶片相对含水量)和7个生理生化特性指标(脯氨酸、丙二醛、叶绿素、可溶性糖、过氧化物酶、超氧化物歧化酶、过氧化氢酶)的隶属函数值,并进一步计算得到抗旱性综合得分。结果表明,干旱胁迫下6种植物抗旱性由强到弱依次是:茅叶荩草、水蔗、三芒草、芸香草、红毛草、黄花猪屎豆。
3 讨论与结论
叶片相对含水量(RWC)能较好反应植物叶片的水分可用性。与蒸腾作用相同,植物在进行呼吸作用时CO2通过叶片气孔进入,水的有效性平衡着CO2的吸收和水分蒸腾[16]。植物受水分胁迫时,自身通过渗透调节作用降低渗透势,使得植物产生足够低的组织水势,以用于持续获取土壤水分[17]。面对干旱胁迫,植物叶片相对含水量变化幅度越小,表明其持水和保水能力越强,细胞受损程度越小,即抗旱能力越强[18]。在本试验中,随土壤水分含量的降低,6个草种RWC呈下降趋势。其中,黄花猪屎豆与三芒草的RWC降幅最大,茅叶荩草降幅最小,即茅叶荩草叶片保水能力较强,而黄花猪屎豆相对较弱。本试验中豆科草种RWC降幅大于禾本科草种,与其他研究结果不同[13]。一方面,可能土壤水分减少,植株根系吸收土壤水分减少,RWC自然降低;另一方面,黄花猪屎豆发达的根系虽具有典型的抗旱特征,但可能由于其叶面积远超出一般豆科牧草,地上部分水分的蒸腾速率远大于地下根系对土壤水分的吸收速率[19]。
除了三芒草,其余5个草种随干旱时间延长其叶片叶绿素含量均有不同程度的降低,这可能是因为干旱胁迫可导致叶绿素合成受阻或叶绿素解体[20-21]。三芒草叶绿素含量呈先降低后升高的趋势,重度干旱时叶绿素含量略有回升,这与其他研究显示干旱胁迫下叶片叶绿素含量略有升高的结果一致[22],说明在干旱后期三芒草仍具有一定的光合能力,能较强维持叶片细胞体内光合碳素同化的进行。复水后,各草种叶绿素含量均显著升高,且红毛草与三芒草叶绿素含量恢复至对照水平。表明干旱胁迫并未对6个草种叶绿素的合成机制造成不可逆的损伤,而红毛草与三芒草在复水情况下可较短时间通过自身调节机制恢复至正常水平。
干旱胁迫下,随水分亏缺植物持续产生活性氧(ROS),引起氧化损伤,胁迫对植物产生不利影响,导致细胞程序性死亡(PCD)[23]。脯氨酸具有较优的水合能力,通过调节细胞渗透压以及消除ROS,是质膜和酶的保护性物质[24]。丙二醛则是活性氧造成脂质过氧化的产物,其含量越多,反映出胁迫对植物造成的损伤越大[25]。本试验中,6个草种丙二醛和脯氨酸含量均随干旱胁迫时间的增加而升高,且在胁迫过程中芸香草的丙二醛积累量最高,说明6个草种受到干旱胁迫后,均受到不同程度的伤害,导致丙二醛含量的升高,胁迫过程中ROS的不断积累,促使植物产生更多脯氨酸消除ROS,而芸香草细胞膜是受干旱胁迫影响最大的一个草种。该结果与干旱胁迫下对不同品种早熟禾(Poaspp.)[26]、狗牙根(Cynodondactylon)[27]、以及扭黄茅[20]等的研究结果一致。同样,可溶性糖作为植物重要的渗透调节物质,通过降低水势,提高水分主动积累以此缓解水分亏缺对植物造成的不利影响[28-29]。本研究中,随干旱时间增加,6个草种可溶性糖整体呈现增加的趋势。其中,茅叶荩草、红毛草、三芒草可溶性糖呈现先减少后增加的趋势,其原因可能是植物在胁迫初期,植物为适应该逆境,主动消耗部分可溶性糖完成体内代谢调节,或者随着胁迫时间的延长,植物本身的渗透调节机制受到抑制,即淀粉分解为葡萄糖的化学反应受到阻碍,但随着干旱胁迫加剧,植物在适宜环境后便不断积累更多的可溶性糖以此减缓胁迫对植物的危害[30]。
植物在受到干旱胁迫初期,低浓度的ROS可起到调节植物生长以及发出胁迫响应信号的作用,然而过多的ROS积累会破坏生物膜的结构及功能。抗氧化酶系统会对ROS进行清除,平衡植物因胁迫产生的过量ROS,通常植物持续受到胁迫后会造成ROS的过量积累[31-32]。本研究结果显示,在干旱条件下6个草种过氧化物酶(POD)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)含量均高于对照组,与前人研究结果一致[33-34]。植物为避免遭受干旱胁迫带来损害,持续产生抗氧化剂和次生代谢物清除积累过多的ROS,保护植物生物膜结构及功能,从而提高植物对干旱胁迫的耐受性,因此抗氧化酶活性不断升高[35-36]。复水后,6个草种POD和SOD活性有所降低,但均未能恢复至对照水平。而水蔗、三芒草、黄花猪屎豆、红毛草的的CAT活性低或接近对照组,芸香草、茅叶荩草CAT活性有所降低但未能恢复至对照组。复水后大部分种质抗氧化酶活性未能恢复至对照组,其原因可能是干旱胁迫对6个草种的损害超出了抗氧化酶清除ROS的能力范围,水蔗、三芒草、黄花猪屎豆和红毛草CAT活性低于或接近对照组也显示存在复水时间不足的可能性。
综上所述,随着干旱程度的加深,不同草种在同一土壤水分梯度下的胁迫响应也不尽相同,复水后,各指标得到了一定恢复。通过隶属函数法对干热河谷6个乡土草种在干旱胁迫下的生物学形态特征指标和生理生化特性指标进行综合比较,得出茅叶荩草抗旱能力最佳,其次是水蔗、三芒草、芸香草、红毛草,黄花猪屎豆抗旱能力最弱。由此可见,茅叶荩草是干热河谷地区生态修复与草地建植的较优乡土草种。
猜你喜欢
快乐语文(2020年31期)2021-01-18 04:04:42
作文与考试·初中版(2020年36期)2020-12-11 09:20:09
语文世界(小学版)(2020年11期)2020-01-08 02:19:30
孩子(2019年8期)2019-08-26 05:43:32
中国检察官·经典案例(2019年2期)2019-04-08 01:10:08
伴侣(2018年7期)2018-07-25 05:47:36
今日农业(2017年3期)2017-12-09 07:58:17
绿色科技(2017年13期)2017-07-31 00:27:30
语文世界(小学版)(2016年1期)2016-02-17 20:54:59
群文天地(2011年22期)2011-08-15 00:43:40
