
Redefinition of the Odorrana versabilis Group,with a New Species from China (Anura,Ranidae, Odorrana)
2024-01-02 06:49:24HanmingSONGSiyuZHANGShuoQIZhitongLYUZhaochiZENGYonghengZHUMinghaiHUANGFuchenLUANZufeiSHUYueningGONGZhifaLIUandYingyongWANG
Asian Herpetological Research 2023年4期
Hanming SONG ,Siyu ZHANG ,Shuo QI* ,Zhitong LYU,2 ,Zhaochi ZENG ,Yongheng ZHU ,Minghai HUANG,Fuchen LUAN,Zufei SHU,Yuening GONG,Zhifa LIU and Yingyong WANG*
1 State Key Laboratory of Biocontrol/The Museum of Biology,School of Life Sciences/School of Ecology,Sun Yat-sen University,Guangzhou 510275,Guangdong,China
2 CAS Key Laboratory of Mountain Ecological Restoration and Bioresource Utilization,Ecological Restoration and Biodiversity Conservation Key Laboratory of Sichuan Province,Chengdu Institute of Biology,Chinese Academy of Sciences,Chengdu 610040,Sichuan,China
3 Shenzhen Shuanghuamu Biological Technology Co.,LTD,Shenzhen 518000,Guangdong,China
4 Chebaling National Nature Reserve,Shaoguan 512000,Guangdong,China
5 Ruyang Forestry Station,Nanling National Nature Reserve,Shaoguan 512000,Guangdong,China
Abstract Taxonomic studies of the Odorrana versabilis group have been consistently disorganized,and they have often been incorrectly associated with an abandoned subgenus or genus Bamburana in existing literature.In this study,we conducted an integrative taxonomic analysis using molecular phylogenetic analysis of two mitochondrial gene fragments (16S rRNA and CO1) and morphological examination of 41 specimens collected during herpetological surveys in southern China.Our results confirmed the monophyly of O. versabilis group which exhibited distinct morphological traits.We also identified a cryptic lineage sister with O.exiliversabilis.Additionally,our study revealed that O.trankieni was nested within O. nasuta.Consequently,we redefined the morphological characteristics of the O. versabilis group,described a new species in this group, O. confusa sp.nov.,and proposed that O.trankieni be considered a junior synonym of O.nasuta.One of the most significant findings of this study was the observation of distinct morphological changes in adult males across all species within the O. versabilis group between the non-breeding and breeding seasons.This study contributes to research on various aspects of Asian amphibian fauna,including taxonomy,ecology,evolution,and physiology.
Keywords morphological changes, Odorrana confusa sp.nov., Odorrana nasuta, Odorrana trankieni,species group,taxonomic revision
1.Introduction
The Odorous frog genusOdorranaFei,Ye and Huang,1990,is a representative anuran fauna in the Oriental Realm.It currently comprises 64 recognized species and is widespread in the southern and eastern Himalayas,through southern China and Indochina,eastward to the Japanese Archipelago,and southward to Malaya and Sumatra to Borneo (Frost,2023).
Fei(1999)proposed that theOdorrana versabilisgroup can be identified by the “presence of well-developed dorsolateral folds.”Later,Feiet al.(2005)erected the subgenusBamburana,designatingO.versabilis(Liu and Hu,1962)as the type species,following a similar definition.Subsequently,Feiet al.(2010)elevated this to the status of a full genus.Chenet al.(2013)conducted a phylogenetic study onOdorranaand identified seven major clades (clade A to G).They found that theO.versabilisgroup,identified as monophyletic crown assemblage clade F,contained five species:O.exiliversabilisLi,Ye and Fei,2001,O.nasica(Boulenger,1903),O.nasutaLi,Ye and Fei,2001,O.tormota(Wu,1977),andO.versabilis.Subsequent studies (Liuet al.,2022;Raoet al.,2022) included two additional species,O.trankieni(Orlov,Le and Ho,2003)andO.yentuensisTran,Orlov and Nguyen,2008,in this clade.
However,most taxonomists have overlooked theOdorrana versabilisgroup(Table 1),despite the invalidation ofBamburanaby Cheet al.(2007).In addition,the morphological traits of these seven species pose challenges for species-level classification.This is because of the conservative interspecific differences during non-breeding seasons,and the significant individual changes in adult males during the breeding seasons.Consequently,some of these species have been incorrectly classified as different species groups or genera(Table 1).As a result,several taxonomic issues persist within this species group,implying the need for taxonomic research that incorporates both phylogenetic and morphological analyses.
Therefore,to resolve taxonomic concerns in theOdorrana versabilisgroup,we integrated the findings from molecular phylogenetic analyses and morphological examinations.We redefined theO.versabilisgroup and documented morphological changes in adult males between the nonbreeding and breeding seasons.Furthermore,based on our herpetological surveys conducted in Guangdong and Guangxi,China,our phylogenetic analysis clusteredO.trankieniwithO.nasuta,and also identified a cryptic lineage sister toO.exiliversabilis,which was confirmed by morphological evidence.Hence,we synonymizedO.trankieniwithO.nasuta,and subsequently described a new species in this taxonomic group.Our study suggests that theO.versabilisgroup comprises seven species.The genusOdorranaconsists of 64 species,42 of which are found in China.
2.Materials and Methods
2.1.Taxon samplingTotal of 22 frog specimens were obtained during fieldwork.After capture,these frogs were euthanized using tricaine methanesulfonate(MS-222),fixed in 4% buffered methanal,and stored in 75% ethanol.Tissue samples,specifically thigh muscles,were excised and preserved in absolute ethanol at-40°C.The specimens were stored at either the Museum of Biology,Sun Yat-sen University(SYS)or Chengdu Institute of Biology,Chinese Academy of Sciences(CIB).This study used specimens for molecular phylogenetic analysis (Table 2) and for morphological comparison and measurement,which were provided in the Taxonomic Accounts.
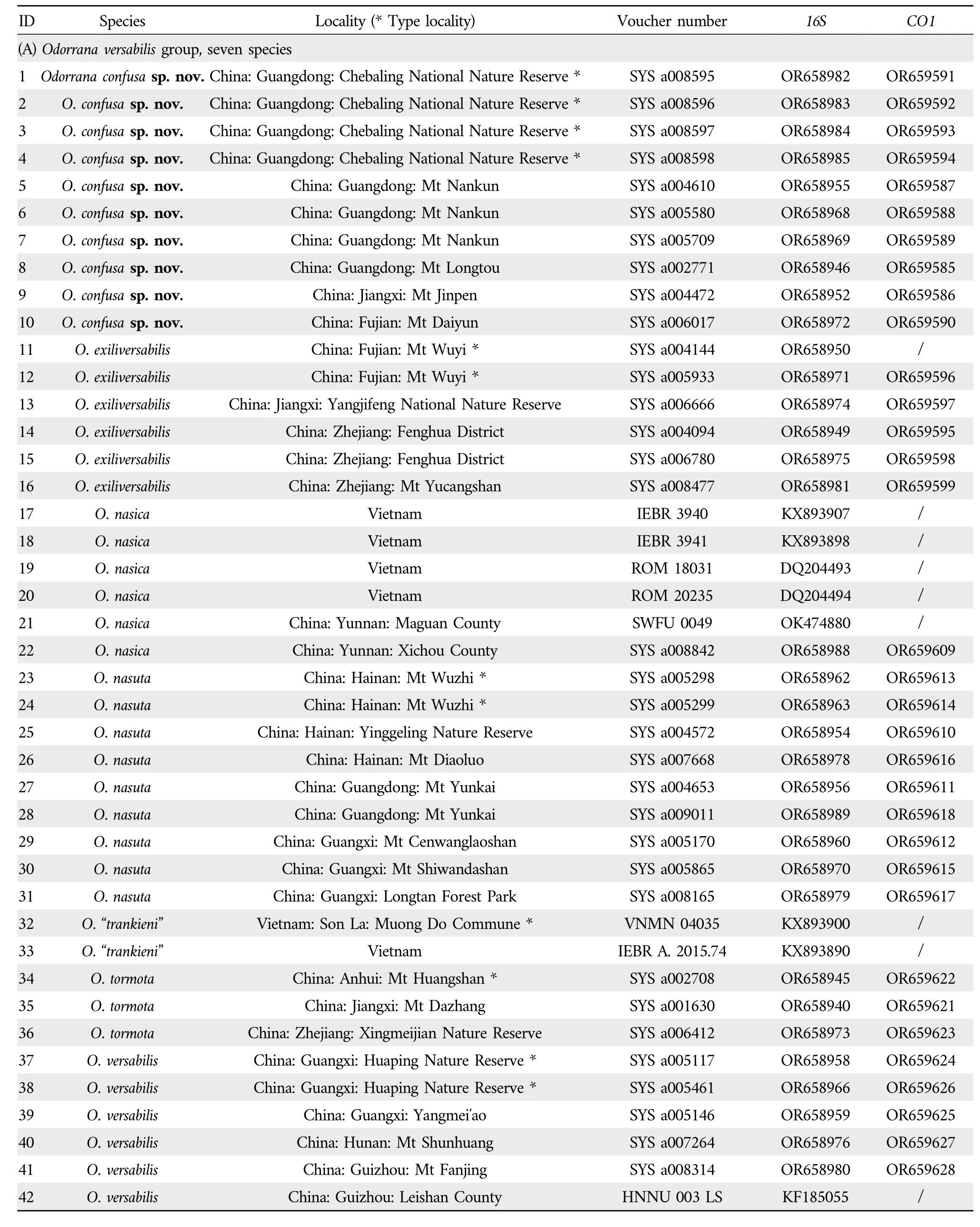
Table 2 Localities,voucher numbers,and GenBank numbers of all samples used in this study for molecular phylogenetic analysis.

Table 4 Comparisons of body size between different populations of Odorrana nasuta.

2.2.Phylogenetic analysisGenomic DNA was extracted from 51Odorranaand twoNidiranasamples (as outgroups)using the DNA extraction kit produced by Tiangen Biotech(Beijing) Co.,Ltd.Two mitochondrial genomic fragments,specifically,the large subunit ribosomal RNA gene(16SrRNA)and the cytochrome C oxidase 1 gene(CO1),were amplified using Ex-Taq DNA polymerase (TaKaRa) for analysis.The primer information is listed in Table 3.The PCR amplification conditions consisted of an initial denaturation step at 95°C for 5 min,followed by 35 cycles of denaturation at 95°C for 40 s,annealing at 49/51°C for 40 s,extension at 72°C for 1 min,and finally an extension step at 72°C for 10 min.After purification with spin columns,PCR products were sequenced using forward primers on an ABI 3730 automated genetic analyzer.In addition,36 sequences ofOdorranaspecies were retrieved from GenBank(Table 2).
The DNA sequences were aligned and trimmed using the algorithm of ClustalW with default parameters in MEGA 6(Thompsonet al.,1997;Tamuraet al.,2013).The sequences used for phylogenetic analysis were concatenated into a matrix with a total length of 1 684 bp,comprising 1 062 bp of the16SrRNA gene and 622 bp of theCO1gene.The nucleotide substitution models GTR+I+G were determined to be the best fit for both of these two genes through testing in jmodeltest v.2.1.2.
We used Bayesian inference (BI) using the software MrBayes 3.2.4 (Ronquistet al.,2012) and maximum likelihood (ML) using the software RaxmlGUI 1.3 (Silvestro and Michalak,2012) to analyze the sequenced data.There were two independent runs being performed for the BI analysis.Each run comprised 10 000 000 generations,with samples taken per 1 000 generations.The first 25% of the samples were excluded as burn-in,resulting in a potential scale reduction factor(PSRF)of <0.005.The evolutionary history of the analyzed taxa was represented using a 50%majority rule consensus tree derived from 1 000 replicates in the ML analysis.
2.3.Morphological charactersThe morphological descriptions and morphometric work adhered to the protocols outlined by Fei and Ye (2016),with slight modifications to meet the current research standards.The formula of sexual size dimorphism index(SSDi)was based on the approach described by Naliet al.(2014).The criteria for the webbing formula were based on the studies by Savage and Heyer(1997)and Cisneros-Heredia and Mcdiarmid(2007).
Measurements were obtained using a Deli DL91200 digital Vernier caliper,with a precision of 0.1 mm.The morphometric characters studied were: (1) Snout-vent length (SVL),the length from the tip of snout to vent;(2)head length(HDL),the length from the terminal of the jaw to tip of snout;(3)head width(HDW),the distance between two sides of jaw;(4)snout length(SNT),the length from anterior end of eye to tip of snout;(5) internasal distance (IND),the distance between inner edges of two nostrils;(6) interorbital distance(IOD),the shortest distance between the inner edges of two upper eyelids;(7) eye diameter (ED),the horizontal diameter of eye;(8)tympanum diameter(TD),the horizontal diameter of tympanum;(9)tympanum-eye distance(TED),the distance from posterior end of eye to anterior of tympanum;(10) hand length (HND),the length from tip of finger Ⅲto terminal of outer metacarpal tubercle;(11) radioulna length(RAD),the length from the terminal of outer metacarpal tubercle to elbow articulation;(12) foot and tarsus length(FTL),the length from tip of toe Ⅳto tibiotarsal articulation;(13) tibia length (TIB),the length from knee to tibiotarsal articulation;and (14) finger Ⅲ disk width (F3W),the horizontal width of the finger Ⅲdigital disk.
Data of all known congeners for morphological comparisons was obtained from specimens stored in the SYS and cited in literature(AmphibiaChina,2023;Bainet al.,2003;Bain and Stuart,2005;Dubois,1992;Feiet al.,2009;Haaset al.,2018;Honget al.,2021;Luonget al.,2021;Mahony,2008;Matsui,1994;Matsui and Ibrahim,2006;Nguyenet al.,2018;Phamet al.,2020;Poyarkovet al.,2021;Raoet al.,2022;Stuartet al.,2005;Stuart and Bain,2005;Stuart and Chan-ard,2005;Stuartet al.,2006;Sumarliet al.,2015).
3.Results
3.1.Monophyly of the Odorrana and O.versabilisgroupBoth BI and ML analyses produced identical topologies,indicating strong support for most nodes with Bayesian posterior probabilities (BPP) ≥0.95 and bootstrap support(BS)≥70(Figure 1).This topology suggested that the species within the genusOdorranaformed a well-supported monophyletic group.Additionally,the genusBamburanawas found to be paraphyletic,indicating the need for its abolition,consistent with previous studies by Cheet al.(2007)and Chenet al.(2013).
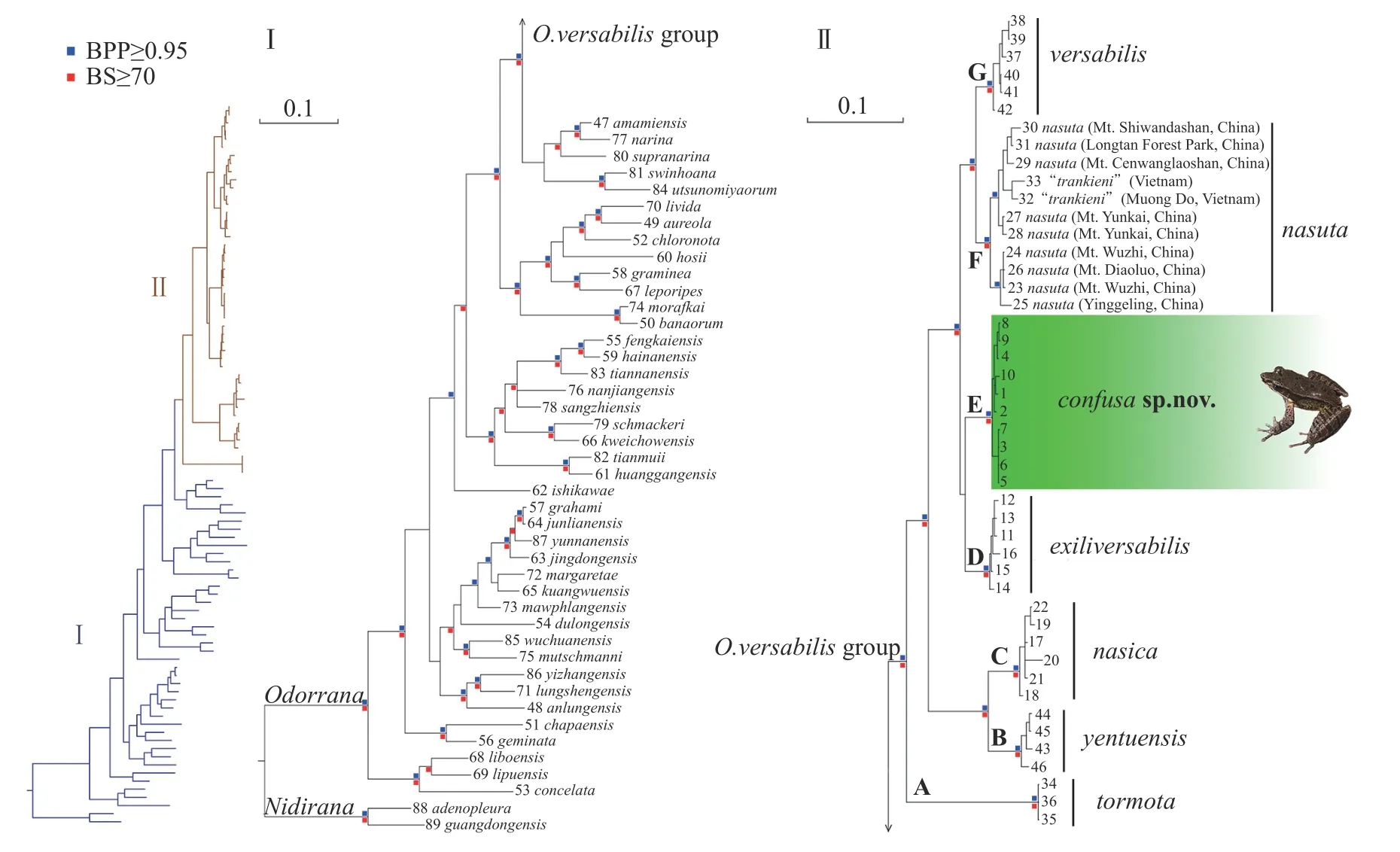
Figure 1 The BI phylogenetic tree of genus Odorrana,inferred by mitochondrial gene fragments 16S rRNA and CO1 of sample in Table 2.Bayesian posterior probabilities(BPP)less than 0.95 and bootstrap supports(BS)less than 70 were left out.Numbers at the tips of all branches are corresponding to the sample IDs in Table 2.
Phylogenetic analysis(Figure 1)indicated that all species of theOdorrana versabilisgroup formed a monophyletic clade within the genus(BPP 0.99,BS 93).This confirmed that clade F (sensu Chenet al.,2013),corresponded specifically to this species group.Morphologically,all members of theO.versabilisgroup shared some common,distinguishable,and exclusive morphological characteristics,including well-developed dorsolateral folds and significant changes in adult male morphology between the non-breeding and breeding seasons(Figures 2-3).Therefore,we redefined theO.versabilisgroup by integrating both the phylogenetic and morphological evidence,adhering to the principle of monophyly.
The clade of theOdorrana versabilisgroup could be further divided into seven highly supported lineages,marked A to G(Figure 1).Among these,lineages D,E,and F warrant in-depth taxonomic analysis.
3.2.A cryptic species previously identified as Odorrana exiliversabilisLineage D of theOdorrana versabilisgroup included samples from western Fujian,Zhejiang,and northern Jiangxi.This lineage represented the species ofO.exiliversabilis,primarily found in Huangkeng,a site located on Mt Wuyi in western Fujian.Lineage E,the sister group to lineage D,comprised samples from northern and eastern Guangdong,southern Jiangxi,and eastern Fujian (Figure 1,Table 2).Samples from Mt Daiyun,Fujian,previously identified asO.exiliversabilis(Feiet al.,2001),were nested in lineage E.However,this lineage showed significant divergence fromO.exiliversabilisin terms of phylogenetics and biogeography(Figures 1 and 4).The genetic distance between lineages D and E was 1.29%.Within each lineage,genetic distances were 0.00%and 0.19%,respectively(Table S1).
Both lineage D and E specimens of theOdorrana versabilisgroup shared common diagnostic characteristics.However,they could be differentiated based on differences in their body size and web shape(as outlined in the Taxonomic Accounts).Based on phylogenetic and morphological evidence,lineage E was identified and classified as a new species.
3.3. Odorrana trankieni,a junior synonym of O.nasutaLineage F of theOdorrana versabilisgroup comprised samples from China (previously identified asO.nasuta) and Vietnam (previously identified asO.trankieni) (Figure 1,Table 2).This topology indicated a close relationship betweenO.trankieniandO.nasutafrom Chinese mainland.Additionally,they were sister taxa ofO.nasutafrom Hainan Island,China (Figure 1).The genetic distance betweenO.trankieniandO.nasutawas 0.81%.InO.trankieni,the genetic distance was 0.76%.Both distances were lower than the genetic distance withinO.nasuta(0.93%;Table S1).
Odorrana trankieniwas originally described asRana trankienibased on the type series collected from Phu Yen,Vietnam.However,O.nasutawas not included in this comparison(Orlovet al.,2003).Phamet al.(2020) suggested that this species is distinct fromO.nasutadue to its larger body size and identified it as endemic to Vietnam.However,Poyarkovet al.(2021)proposed a need for additional verification to establish its taxonomic status.
ConspecificOdorranaspecimens were collected between 2015 and 2022 from Guangdong and Guangxi in China.Based on Feiet al.(2009) and Phamet al.(2020),these specimens were identified asO.nasutarather thanO.trankieni.This finding extended the distribution ofO.nasutato Chinese mainland.However,morphometric data (Tables 4 and S2)indicated that the SVL of someO.nasutaspecimens from China overlapped with that ofO.trankieni.This suggested that the body size difference previously used to distinguish between these two species,as proposed by Phamet al.(2020),was no longer reliable.Additionally,previous studies have also reported conflicting findings regarding the presence of supratympanic folds inO.trankieni(Orlovet al.,2003;Phamet al.,2020)but the absence inO.nasuta(Feiet al.,2009).Hence supratympanic folds were considered the only morphological distinction between the two species.Therefore,we analyzed photographs and specimens of both species and concluded that the supratympanic folds ofO.trankieniwere due to misidentification of the loosing skin on the swollen shoulder regions of adult males.This misinterpretation was irrelevant for species differentiation and highlighted the importance of taxonomists in avoiding describing amphibians based solely on desiccative specimens.
In conclusion,our integrative research confirmed the invalidity ofOdorrana trankieniand supported its synonymization withO.nasutabased on a combination of phylogenetic and morphological evidence.Based on the principle of priority,we consideredO.trankienias a junior synonym ofO.nasutain the following discussion.
4.Taxonomic Accounts
4.1.Redefinition of the Odorrana versabilis group
Order Anura Hogg,1839
Family Ranidae Batsch,1796
GenusOdorranaFei,Ye and Huang,1990
Odorrana versabilisgroup
Morphological definitionTheOdorrana versabilisgroup can be distinguished from otherOdorranaspecies based on specific morphological characteristics,including (1) well-developed dorsolateral folds;(2) significant morphological changes in adult males between the non-breeding and breeding seasons,during the breeding seasons,elongation of the snout beyond the lower jaw,lateral swelling of the shoulder regions,swelling of the dorsolateral folds,and increased robustness of the forelimbs;(3) shagreened skin,occasionally with conical or sharp tubercles behind the sacrum,around dorsolateral folds,or on rictal,temporal,and loreal regions,or the upper and lower lips;and(4) adult females larger than adult males,but absence of extreme sexual size dimorphism,SSDi from 0.51 to 0.97.
Suggested Chinese name竹叶蛙种组(zhú yè wā zhǒng zǔ).
RemarksFeiet al.(2001)and Liet al.(2001)were the first to document the morphological changes of adult males inOdorrana nasutaduring breeding seasons.These changes include the elongation of the snout and swelling of the head.However,previous researchers have been unaware of its potential as a diagnostic morphological characteristic for theO.versabilisgroup.Morphological examination revealed these changes occurring in each species of this group.Therefore,these changes were proposed as a term of morphological definition of theO.versabilisgroup;comparative images of some species are provided in Figure 3.

Figure 3 Morphological comparisons of adult males of (A) Odorrana confusa sp.nov.,(B) O. exiliversabilis,(C) O. nasuta and (D) O. versabilis between (1) the non-breeding season and (2) the breeding season in life.Photos by Yingyong WANG,Jian WANG,Zhaochi ZENG,Zhitong LYU,and Yongheng ZHU.
Key to species of Odorrana versabilis group
1a Tympanum dramatically concave,nearly forming an external auditory canal in adult males...........................O.tormota
1b Tympanum not concave or just slightly concave,not forming an external auditory canal in both sexes.......................2
2a Edge of web on toe Ⅳnot reaching disk...........................3
2b Edge of web on toe Ⅳreaching disk....................................5
3a Dorsal skin behind sacral vertebrae slightly smooth,no or less tubercles in both sexes..................................O.exiliversabilis
3b Dorsal skin behind sacral vertebrae rough,with numerous conical spinules,especially in adult males during the breeding seasons...................................................................................4
4a Web unfolded narrowly,the angle between toes ⅠandⅤless than or approximately equal to 90° when the web unfolded completely....................................................................O.nasica
4b Web unfolded widely,the angle between toes Ⅰand Ⅴgreater than 90°when the web unfolded completely..............O.yentuensis
5a Flanks with messy stains and numerous tubercles;rictal glands and lower jaws of adult males with numerous prominent white conical spinules,especially during the breeding seasons.....................................................O.confusasp.nov.
5b Flanks relatively clean,with few sparse tubercles;rictal glands and lower jaws of adult males with sparse or few white conical spinules..............................................................................................6
6a Web unfolded narrowly,the angle between toes ⅠandⅤless than or approximately equal to 90° when the web unfolded completely;adult male with a pair of external subgular vocal sacs......................................................................O.nasuta
6b Web unfolded widely,the angle between toes Ⅰand Ⅴgreater than 90° when the web unfolded completely;adult male with a pair of internal subgular vocal sacs...........O.versabilis
4.2.Revision of Odorrana exiliversabilis and description of the new species
Odorrana exiliversabilis Li,Ye and Fei,2001
Rana versabilis—Guoet al.,1966;Dinget al.,1980;Cai,1981.
Odorrana versabilis—Feiet al.,1990;Yeet al.,1993;Feiet al.,1999.
Odorrana(Bamburana)exiliversabilis—Feiet al.,2005,2009.
Odorrana exiliversabilis—Chenet al.,2005;Wanget al.,2015.
Huia exiliversabilis—Frostet al.,2006.
Bamburana exiliversabilis—Feiet al.,2010.
HolotypeCIB 64I2013,adult male,collected from Huangkeng (27.60 °N,117.63 °E;ca 1 030 m a.s.l.),Jianyang County(=Jianyang District),Fujian Province,China.
ParatypesFifteen specimens collected from the same locality as holotype:five adult males CIB 64I2008,2009,2205,2208,2263,and ten adult females CIB 64I2010-2012,2134,2206,2207,2209,2298,2299,2309;forty-nine specimens collected from Sangang and Miaowan(ca 600-1 100 m a.s.l.),Chongan County (=Wuyishan City),Fujian Province,China: 18 adult males CIB 64I1553,2318-2320,2543-2545,2587,2760,2761,2764,2765,2775,3415-3417,4101,4155,and 30 adult females,CIB 64I2316,2317,2342-2346,2583-2586,2752-2759,2762,2763,2772-2774,3418,3433,3669,3670,4031,4032,and one juvenile CIB 64I4102;and two specimens collected from Daiyunshan (=Mt Daiyun;ca 1 525 m a.s.l.),Dehua County,Fujian Province,China:two adult females CIB 64II0334,0335.
Specimens examinedFour specimens collected from Mt Wuyi,Wuyishan City,Fujian Province,China:one adult male SYS a005931,and three adult females SYS a004144,5932,5933;nine specimens collected from Licun,Fenghua District,Zhejiang Province,China:six adult males SYS a006775-6780,and three adult females SYS a006150,6151,6155;and one specimen collected from Yangjifeng Nature Reserve,Guixi City,Jiangxi Province,China:one adult female SYS a006665.
Revised diagnosisOdorrana exiliversabiliscan be differentiated from its congeners based on the following combination of characteristics:(1)well-developed dorsolateral folds;(2) prominent morphological changes in adult males between the non-breeding and breeding seasons,during the breeding seasons,elongation of the snout beyond the lower jaw,lateral swelling of the shoulder regions,dorsolateral fold enlargement,and increased robustness of the forelimbs;(3)shagreened skin with occasional conical or sharp tubercles behind the sacrum,around the dorsolateral folds,or on the rictal,temporal,and loreal regions,or the upper and lower lips;(4) tympanum distinct and round,with a diameter of approximately half of the eye diameter,not concave or just slightly concave,not forming an external auditory canal in both sexes;(5)tips of fingers expanding into disks;(6)edge of web on toe Ⅳ not reaching the disk;(7) dorsal skin behind sacral vertebrae slightly smooth,with either no or fewer tubercles in both sexes;and (8) supratympanic folds absent.
EtymologyThe specific name,exiliversabilis,is derived from the combination of“exili-”and“versabilis,”indicating that this species is smaller in body size thanOdorrana versabilis.
Suggested common name“Little Bamboo-leaf Frog” in English and“小竹叶蛙(xiǎo zhú yè wā)”in Chinese.
DistributionThe phylogenetic and morphological findings suggested thatOdorrana exiliversabilisis endemic to China and is currently distributed in Mt Wuyi in Fujian,Yangjifeng in Jiangxi,Mt Yucang and Fenghua District in Zhejiang,and Mt Huangshan in Anhui (Figure S1).Additional research should be dedicated to this species to better understand its precise distribution range.
RemarksThe speciesOdorrana exiliversabiliswas initially described by Feiet al.(2001)in Huangkeng,Mt Wuyi,Fujian,China.The type series of this species includes two paratypes,CIB 64Ⅱ0334 and CIB 64Ⅱ0335,which were collected from Mt Daiyun,Fujian,China.However,based on our taxonomic revision,the population in Mt Daiyun should be reclassified asO.confusasp.nov.Consequently,these two specimens should be excluded from the type series ofO.exiliversabilis,and attributed to the historical records ofO.confusasp.nov.
Odorrana confusa Song,Zhang,Qi,Lyu,Zeng and Wang sp.nov.
Rana versabilis—Dinget al.,1980;Zou,1983.
Odorrana exiliversabilis—Feiet al.,2001.
Odorrana(Bamburana)exiliversabilis—Feiet al.,2005,2009.Odorrana versabilis—Raoet al.,2013;Liet al.,2018;Gong and Wu,2020.
Bamburana versabilis—Yanget al.,2018.
HolotypeSYS a008595,adult male,collected by Jian WANG,Yingyong WANG and Xinwang ZHANG,on 28 March 2021,from Chebaling National Nature Reserve,Shixing County,Guangdong Province,China.
ParatypesThree specimens with the same collection information as the holotype: SYS a008596/ CIB 119029,adult male,and SYS a008597,8598,adult females;and three specimens collected from Mt Nankun,Longmen County,Guangdong Province,China: SYS a004610,adult male,collected by Jian WANG and Zhitong LYU,on 22 March 2016,SYS a005580,adult female,collected by Yingyong WANG,Zhitong LYU and Jian WANG,on 11 November 2016,and SYS a005709,adult female,collected by Jian WANG and Zhitong LYU,on 8 April 2017.
DiagnosisOdorrana confusasp.nov.is distinguishable from its congeners based on the following combination of characteristics: (1) well-developed dorsolateral folds;(2)prominent morphological changes in adult males between the non-breeding and breeding seasons,during the breeding seasons,elongation of the snout beyond the lower jaw,lateral swelling of shoulder regions,dorsolateral fold swelling,and increased robustness of the forelimbs;(3)shagreened skin with occasional conical or sharp tubercles behind the sacrum,around dorsolateral folds,or on the rictal,temporal,and loreal regions,or the upper and lower lips;(4) tympanum distinct and round,not concave or just slightly concave,not forming an external auditory canal in both sexes;(5) tips of fingers expanding into disks;(6)edge of web on toe Ⅳreaching the disk;(7) dorsal skin behind sacral vertebrae slightly smooth,with either no or few tubercles in both sexes;(8)flanks with messy stains and numerous tubercles;(9) rictal glands and lower jaws of adult male with numerous prominent white conical spinules,especially during breeding seasons;and(10)body size large,with the SVL of adult males and females averaging 68.5 mm (64.4-72.5 mm) and 70.9 mm(67.0-73.9 mm),respectively.
ComparisonsOdorrana confusasp.nov.can be distinguished from all other members of theO.versabilisgroup based on morphological characteristics.
Odorrana confusasp.nov.has been misidentified asO.exiliversabilis.However,it differs fromO.exiliversabilisby (1)larger body size,with the SVL of adult males and females averaging 68.5 mm (64.4-72.5 mm) and 70.9 mm(67.0-73.9 mm),respectively [vsadult males and females averaging 45.7 mm (41.9-49.5 mm) and 58.0 mm(51.0-66.5 mm),respectively] and (2) fully developed webbing,with the edge of the web on toe Ⅳreaching the disk(vsnot fully developed webbing and edge of web on toeⅣnot reaching the disk).
Odorrana confusasp.nov.differs fromO.nasicaby (1) a relatively smooth dorsal skin surface behind the sacral vertebrae in both sexes,lacking or having fewer tubercles(vsrough dorsal skin with numerous conical spinules,especially in adult males during the breeding seasons)and(2)edge of the web on toe Ⅳreaching the disk(vsedge of the web on toe Ⅳnot reaching the disk).
Odorrana confusasp.nov.differs fromO.nasutaby(1)flanks with messy stains and numerous tubercles(vsrelatively clean flanks,with few sparse tubercles) and (2) rictal glands and lower jaws of adult males with numerous prominent white conical spinules,especially during breeding seasons(vssparse or few white conical spinules in these areas in adult males).
Odorrana confusasp.nov.differs fromO.tormotaby (1) a larger body size,with the SVL of adult males and females averaging 68.5 mm (SVL 64.4-72.5 mm) and 70.9 mm(67.0-73.9 mm),respectively [vsadult males and females averaging 32.4 mm (28.9-36.6 mm) and 58.7 mm(54.9-62.1 mm),respectively] and (2) not concave or just slightly concave tympanum,not forming an external auditory canal in both sexes(vsdramatically concave tympanum,nearly forming an external auditory canal in males).
Odorrana confusasp.nov.differs fromO.versabilisby (1)flanks with messy stains and numerous tubercles(vsrelatively clean flanks,with few sparse tubercles) and (2) rictal glands and lower jaws of adult males with numerous prominent white conical spinules,especially during the breeding seasons(vssparse or few white conical spinules in these regions in adult males).
Odorrana confusasp.nov.differs fromO.yentuensisby(1)a larger body size,with the SVL of adult males and females averaging 68.5 mm(SVL 64.4-72.5 mm)and 70.9 mm(67.0-73.9 mm),respectively [vsadult males and females averaging 42.5 mm(41.6-43.4 mm) and 58.8 mm(50.4-61.3 mm),respectively],(2) a relatively smooth dorsal skin surface behind sacral vertebrae in both sexes,lacking or with fewer tubercles (vsrough dorsal skin behind sacral vertebrae,with numerous conical spinules,especially in adult males during the breeding season),and(3)edge of web on toe Ⅳreaching the disk(vsedge of web on toe Ⅳnot reaching the disk).
Description of holotypeAdult male.Body slender and moderate in size,SVL 72.5 mm.Head longer than width(HDL/HDW=1.22);snout long,prominently projecting beyond lower jaw;triangular mastoid on tip of the lower jaw,just embedded in notch on upper jaw;snout length longer than eye diameter (SNT/ED=1.46);loreal region concave,with distinct canthus rostralis;nostril between snout and eye,closer to tip of snout than to eye;maxillary edge with some tiny white mastoids;eye large;tympanum distinct,round and not concave,not forming an external auditory canal,tympanum diameter approximately half of eye diameter(TD/ED=0.52);numerous distinct white conical spinules around tympanum,on loreal,temporal,and rictal regions;top of head flat,pineal body invisible;internasal distance longer than interorbital distance (IND/IOD=1.39);shoulder region swollen;vomerine ridge present with vomerine teeth;internal naris distinct;the free end of tongue notched.
Forelimbs stout;fingers long but thin,relative finger lengthsⅡ<Ⅳ<Ⅰ<Ⅲ;the finger tips moderately expanded into oval disks,each finger with lateroventral groove;finger web absent;all fingers with slight fringes;subarticular tubercles prominent,1,1,2,and 2,respectively;metacarpal tubercles elongated and oblate.
Hindlimbs long and moderately strong,tibiotarsal articulation exceeding the end of snout when leg stretched alongside of body,and heels overlapping when thighs flexed vertically to the axis of body;toes slender,relative toe lengthsⅠ<Ⅱ<Ⅲ<Ⅴ<Ⅳ;toe tips expanded into oval disks,each toe with lateroventral groove;toe fully webbed,webbing formulaⅠ0-0+Ⅱ0-0Ⅲ0-0+Ⅳ0+-0Ⅴ;fringes on toes Ⅰand Ⅴslightly visible;subarticular tubercles big and ellipsoidal,1,1,2,3,2 respectively;inner metatarsal tubercle large and oblong,outer metatarsal tubercle round but small;no tarsal fold.
Dorsal skin smooth,sparsely granular;dorsum of head bearing some granules;rictal glands prominent and beanshaped;dorsolateral fold well-developed,from end of eye to hip;flank with several granules;a few tubercles around and under vent;dorsal skin of limbs slightly granular.Ventral surface of head,body,and forelimbs smooth,thighs with many tubercles,and tibia and tarsus very smooth.
Coloration of holotypeIn life (Figure 4): dorsal surface of head and body light brown,with a bit of dark brown spots and irregular green stains;iris yellowish-brown on the upper one fourth and dark orange on the lower three fourth;rictal glands light yellow;flank brownish-black,with yellow tubercles;dorsal skin of limbs light brown,with clear dark brown transverse bars;nuptial pads vivid waxy yellow;ventral surface of head and chest milk-white,abdomen bright yellow,limbs yellow,and scattered irregular brown spots.

Figure 4 Holotype of Odorrana confusa sp.nov.in life on(A)dorsolateral view,(B)dorsal view,(C)ventral view,(D)head,(E)hand,(F)foot,(G) nuptial pad and (H) habitat.Photos by Jian WANG and Shuo QI.
In preservative(Figure 5):dorsal surface of head and body dark brown,spots and stains invisible or scarcely visible;tiny white spines on loreal and temporal regions become more distinct;rictal glands and tubercles on flank milk-white;dorsal skin of limbs darkened,transverse bars indistinct;nuptial pads greyish-white;ventral surface of head and chest dirty and tawny,abdomen maize-yellow,limbs beige,and mottled with dark brown patches.

Figure 5 Holotype of Odorrana confusa sp.nov.in preservative on(A)dorsal view and(B)ventral view.White scale bar shows the length of 10 mm.Photos by Shuo QI.
VariationsTable 5 lists the measurements for the type series.All paratypes are morphologically similar to the holotype,except for the following traits:(1)sparse and slightly distinct white conical spinules on the lower edge of the maxilla in SYS a 008596;(2) distinct,tiny,and white spinules exclusively around tympanum in SYS a 008598;(3)pineal body visible in SYS a004610,SYS a005580,SYS a005709,and SYS a008597-8598;(4) right hand abnormal,with finger Ⅲbeing shorter and approximately equal to finger Ⅱ,two subarticular tubercles of finger Ⅲoverlapping in SYS a 008597;(5)hindlimbs strong in SYS a 004610;(6) when leg stretched alongside of body,tibiotarsal articulation reaching the end of snout in SYS a004610 and SYS a008598,reaching nostril in SYS a 005580;(7)outer metatarsal tubercle relatively small in SYS a 008596;(8)dorsum of head smooth in SYS a004610 and SYS a 005580;(9) rictal glands moderately prominent and relatively small in SYS a 008597;(10)dorsal skin of tibia and tarsus with several tiny white spines in SYS a005580 and slightly distinct in SYS a005709.
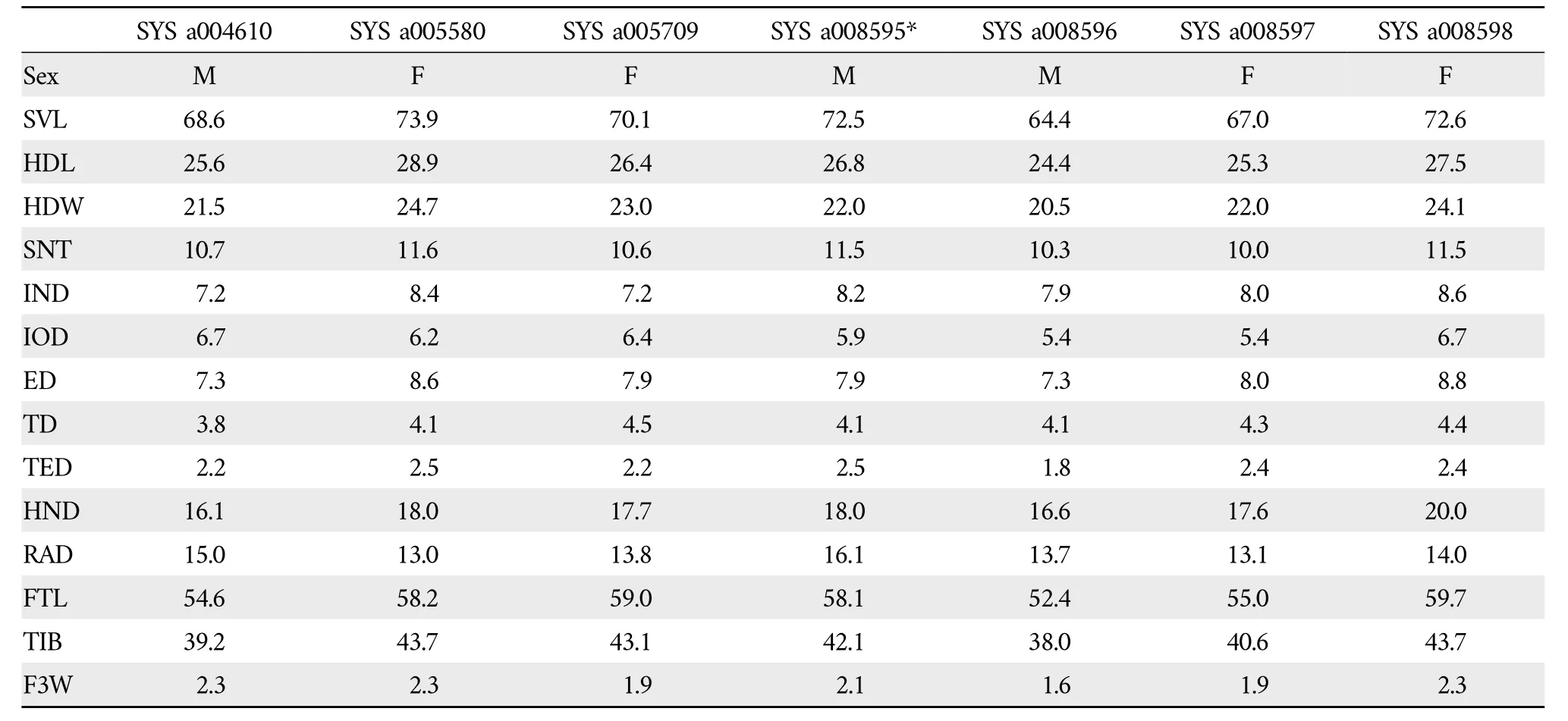
Table 5 Morphological measurements of type series of Odorrana confusa sp.nov.(in mm,*=holotype,M=Male,F=Female).
Sexual dimorphismThe average body sizes of adult females(SVL 67.0-73.9 mm,70.9 mm on average,n=4)are slightly larger than males(SVL 64.4-72.5 mm,68.5 mm on average,n=3),with an SSDi of 0.97;males have a pair of subgular vocal sacs and stronger forelimbs than females.
Adult males exhibited significant morphological transformations during the transition from the nonbreeding to breeding seasons.In non-breeding seasons(Figure 3 A1),the snout appeared round in dorsal view,slightly extending beyond the lower jaw;rictal glands weak or medium-sized;spines on loreal and temporal regions,lips,and rictal glands weak and indistinct;dorsolateral folds distinct but thin,with individual granules;shoulder regions not swollen;and forelimbs moderate.During breeding seasons (Figure 3 A2),the snout appeared elongated and protruded in the dorsal view,extending beyond the lower jaw;rictal glands developed;spines on loreal and temporal regions,lips,and rictal glands prominent;dorsolateral folds swollen,with indistinct tubercles;lateral swelling of shoulder regions,the widest on dorsal view;forelimbs sturdy,with a developed velvety nuptial pad occurred on the inner side of finger Ⅰ.
EtymologyThe specific name,confusa,is derived from the feminine form of the Latin adjective “confusus,” meaning“confused”or“mixed.”This refers to the chronical taxonomic confusion of this newly discovered species with theOdorrana exiliversabilis.
Suggested common name“Chebaling bamboo-leaf frog”in English and “车八岭竹叶蛙(chē bā lǐng zhú yè wā)” in Chinese.
DistributionThe phylogenetic and morphological findings suggested thatOdorrana confusasp.nov.is endemic to China,specifically in Chebaling,Mt Nankun and Mt Longtou in Guangdong,Mt Jinpen in Jiangxi,and Mt Daiyun in Fujian(Figure S1).Further research is required to better understand the precise distribution of this species.
Natural historyOdorrana confusasp.nov.inhabits moist subtropical montane regions,typically surrounded by a combination of evergreen and deciduous broadleaved forests.This species is commonly found in close proximity to or within torrential streams.Male specimens collected in March and November displayed developed velvety nuptial pads,whereas female specimens showed mature eggs.This suggests that the breeding season for this species occurs in November and March.However,the reproductive calls and tadpoles of this species remain unknown.
4.3.Supplementary description of Odorrana nasuta
Odorrana nasuta Li,Ye and Fei,2001
Rana versabilis—Liuet al.,1973;Shiet al.,2011.
Rana trankieniOrlov,Le and Ho,2003.
Odorrana(Bamburana)nasuta—Feiet al.,2005,2009.
Odorrana(Bamburana)versabilis—Feiet al.,2005,2009.
Odorrana nasuta—Chenet al.,2005;Cheet al.,2007.
Huia nasuta—Frostet al.,2006.
Huia trankieni—Frostet al.,2006.
Odorrana trankieni—Cheet al.,2007;Phamet al.,2020.
Rana nasuta—Stuartet al.,2008.
Bamburana nasuta—Feiet al.,2010.
Bamburana trankieni—Feiet al.,2010.
HolotypeCIB 64III0462,adult male,collected from Wuzhi Shan (=Mt Wuzhi,18.90°N,109.67°E;ca 720 m a.s.l.),Qiongzhong County,Hainan Province,China.
ParatypesFour specimens collected from the same locality as the holotype:two adult males CIB 64III0795,0796,and two adult females CIB 64III0794,0797;three specimens collected from Dali(ca 350 m a.s.l.),Lingshui County,Hainan Province,China:three adult males CIB 64III2305-2307;four specimens collected from Yinggeling (ca 520-850 m a.s.l.),Baisha County,Hainan Province,China: four adult males CIB 64III3858,3938-3940.
Specimens examinedFour specimens collected from Mt Wuzhi,Wuzhishan City,Hainan Province,China: four adult females SYS a005296-5299;one specimen collected from Yinggeling,Baisha County,Hainan Province,China:one adult male SYS a 004572;seven specimens collected from Mt Shiwandashan,Shangsi County,Guangxi Zhuang Autonomous Region,China: one adult male SYS a005864,and six adult females SYS a003517,3540-3542,5859,5865;four specimens collected from Mt Yunkai,Xinyi City,Guangdong Province,China: three adult males SYS a004653,6035,6863,and one adult female SYS a 004656;three specimens collected from Mt Cenwanglaoshan,Tianlin County,Guangxi Zhuang Autonomous Region,China: two adult males SYS a005167,5169,and one adult female SYS a 005168;one specimen collected from Longtan Forest Park,Guiping City,Guangxi Zhuang Autonomous Region,China:one adult male SYS a008165.
Revised diagnosisOdorrana nasutadiffered from all other congeners based on the following combination of characteristics: (1) well-developed dorsolateral folds;(2)prominent morphological changes in adult males between the non-breeding and breeding seasons,during the breeding seasons,elongation of the snout beyond the lower jaw,lateral swelling of the shoulder regions,dorsolateral fold swelling,and increased robustness of the forelimbs;(3)shagreened skin with occasional conical or sharp tubercles behind the sacrum,around dorsolateral folds,or on the rictal,temporal,and loreal regions,or the upper and lower lips;(4) tympanum distinct and round,not concave or just slightly concave,not forming an external auditory canal in both sexes;(5) tips of fingers expanding into disks;(6)edge of web on toe Ⅳreaching the disk;(7) dorsal skin behind sacral vertebrae slightly smooth,with few or no tubercles in both sexes;(8)supratympanic folds absent;(9) relatively clean flanks,with few sparse tubercles;(10)rictal glands and lower jaws of adult males with sparse or few white conical spinules;(11) web unfolded narrowly,the angle between toe Ⅰand Ⅴless than or approximately equal to 90° when web unfolded completely;and (12) adult males with a pair of external subgular vocal sacs.
Supplementary descriptionMorphologically,Odorrana nasutapopulations from various regions of Chinese mainland,including Mt Yunkai,Mt Shiwandashan,Mt Cenwanglaoshan,and Longtan Forest Park,closely resemble conspecific individuals from Hainan Island in China and northern Vietnam,except for variations in body size.Specimens from Hainan Island are typically slightly smaller or comparable in size to those from Mt Yunkai.In contrast,the specimens from northern Vietnam and Mt Shiwandashan were collectively the largest.The specimens from Mt Cenwanglaoshan and Longtan Forest Park were of intermediate sizes(Table S2).
Adult males ofOdorrana nasutaexhibit significant morphological transformations during the transition from non-breeding to breeding seasons.In non-breeding seasons,snout is not elongated,and appears round in dorsal view,protruding slightly beyond the lower jaw;rictal glands weak or medium-sized;temporal regions and lips lack or have indistinct spines;dorsolateral folds distinct but thin,with distinct granules;shoulder regions not swollen;forelimbs moderate (Figure 3 C1).In breeding seasons,snout becomes elongated and prominently extends beyond the lower jaw;rictal glands developed;prominent spines on temporal regions and lips;dorsolateral folds swollen,with indistinct granules;lateral swelling of shoulder regions;forelimbs sturdy,with a developed velvety nuptial pad on the inner side of finger Ⅰ(Figure 3 C2).
Based on our morphological analysis,individuals in the northern Vietnam populations ofOdorrana nasuta,previously reported asO.trankieni,lack the morphological character of“well developed supratympanic folds”(Orlovet al.,2003).This incorrect description was a result of misidentification of the loosing skin on the swollen shoulder regions of adult males.EtymologyThe specific name,nasuta,means“big-nosed”in Latin and refers to the prominently projecting snout of this species.
Suggested common name“Duck-beak Bamboo-leaf Frog”in English and“鸭嘴竹叶蛙(yā zuǐ zhú yè wā)”in Chinese.
DistributionPhylogenetic and morphological findings indicated thatOdorrana nasutais distributed around the Beibu Gulf (=Tonkin Bay),including Chinese mainland,Hainan Island of China,and northern Vietnam(Figure S1).
5.Discussion
5.1.The dramatic morphological changes of adult males of the Odorrana versabilis group between the non-breeding and breeding seasonsAdult males of theOdorrana versabilisgroup undergo significant morphological changes between non-breeding and breeding seasons.These changes include elongated snouts,swollen shoulder regions,stronger arms,and development of nuptial pads and dorsolateral folds.These distinctive changes serve as significant morphological characteristics that define this species group.Feiet al.(2001) initially observed these characteristics onO.nasuta,and our morphological analysis confirmed that all species within this species group collectively share this trait.We described these morphological changes in Taxonomic Accounts and updated previous definitions and diagnoses.However,more extensive taxonomic research is required to provide a comprehensive analysis and evaluation of each species to deepen our understanding of this species group.Future research should prioritize providing detailed descriptions,illustrations,and physiological mechanisms underlying these morphological changes to reveal the ecological significance of this morphological trait.
5.2.The need of further research on the new species
Odorrana confusa sp.nov.Species in theOdorrana versabilisgroup exhibit conservative morphological differences due to their shared habitat and adaptive responses.O.confusasp.nov.has been chronically mistaken forO.exiliversabilisbecause of their morphological similarities (Feiet al.,2001,2005,2009).This study used phylogenetic and morphological analyses to identify and compare the morphology of the two species.However,crucial biological information regarding this new species,such as bioacoustic data and life-cycle details,remains unknown.This information is crucial for understanding the physiological and ecological characteristics of the species,as well as its species-level limitations.Future research should prioritize these aspects to facilitate a comprehensive understanding of this species.
Nomenclatural acts registrationThe electronic version of this article in portable document format represents a published work according to the International Commission on Zoological Nomenclature (ICZN),and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone(see Articles 8.5-8.6 of the Code).This published work and the nomenclatural acts it contains have been registered in ZooBank,the online registration system for the ICZN.The ZooBank LSIDs(Life Science Identifiers)can be resolved and the associated information can be viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/.
Publication LSID: urn:lsid:zoobank.org:pub:A402A6DF-2E42-49F4-9A1E-1C95068EE95E
Odorrana confusaLSID: urn:lsid:zoobank.org:act:A0622C35-2877-48FB-BD1E-C1A10DB383BD
AcknowledgementsWe thank Jian WANG,Honghu CHEN,Xinwang ZHANG,Mengchan YANG,Jiahe LI,and Zhenhong KONG for their help on fieldworks,Robert W.MURPHY for offering required references,Shun MA for facilitating access to specimens,and Jiajun ZHOU and Weiliang XIE for offering required photos.We also thank Bryan STUART and another anonymous reviewer for their valuable suggestions,and an anonymous native English speaker at ZYEdit for polishing the linguistic expression.This work was supported by DFGP Project of Fauna of Guangdong-202115,Nanling National Park Construction Project (No.ZD22-ZC016AG),2022 Wildlife Monitoring Projects of the Guangdong Forestry Bureau,and the National Animal Collection Resource Center,China.
猜你喜欢
中国野生植物资源(2023年9期)2023-10-20 09:11:32
河北农机(2021年11期)2021-11-29 13:16:20
少儿美术(2020年9期)2020-11-05 09:11:54
孩子·小学版(2019年11期)2019-09-10 09:33:39
陶山(2019年1期)2019-03-29 01:03:02
农业机械学报(2018年11期)2018-12-04 09:03:34
东坡赤壁诗词(2018年1期)2018-03-31 09:10:10
湖南农业大学学报(自然科学版)(2017年6期)2017-12-21 23:29:20
农业机械学报(2017年9期)2017-10-11 01:33:48
女士(2015年6期)2015-05-30 20:22:32
 Asian Herpetological Research2023年4期
Asian Herpetological Research2023年4期
- Asian Herpetological Research的其它文章
- Response of Distribution Range Against Climate Change and Habitat Preference of Four National Protected Diploderma Species in Tibetan Plateau
- A New Species of the Genus Nanorana Günther,1896 (Anura:Dicroglossidae) from Hengduan Mountains of China
- Evidence for Expensive Tissue Hypothesis in the Asian Common Toad(Duttaphrynus melanostictus)
- Sexual Dimorphism,Female Reproductive Characteristics and Embryonic Thermosensitivity in the Tonkin Forest Skink (Sphenomorphus tonkinensis)from Hainan,South China
- Next-generation Sequencing of MHC Class I Genes Reveals Trans-species Polymorphism in Eutropis multifasciata and Other Species of Scincidae
- Oxygen Availability Affects Behavioural Thermoregulation of Turtle Embryos