
Oxygen Availability Affects Behavioural Thermoregulation of Turtle Embryos
2024-01-02 06:49LiangLIANGandWeiguoDU
Asian Herpetological Research 2023年4期
Liang LIANG and Weiguo DU
Key Laboratory of Animal Ecology and Conservation Biology,Institute of Zoology,Chinese Academy of Sciences,Beijing 100101,China
Abstract Extending recent findings that reptile embryos seek optimal temperatures inside eggs for thermoregulation,our study demonstrates that this thermoregulatory behaviour can be affected by the amount of oxygen available to an embryo.We exposed embryos of a freshwater turtle (Mauremys reevesii) to two heat sources (an optimal temperature of 30°C and a high temperature of 33°C)under three different oxygen levels-hypoxia(12%O2),normoxia(21%O2)and hyperoxia(30%O2)-and quantified the interactive effects of temperature and oxygen availability on embryonic thermoregulatory behaviour.Our results demonstrated that,in both thermal treatments,embryos exposed to hypoxia did not move as close to the heat source and therefore selected lower temperatures than those exposed to normoxia or hyperoxia.Embryos may select low temperatures under hypoxic conditions to decrease oxygen consumption and therefore alleviate the negative impact of hypoxic stress.
Keywords behavioural thermoregulation,embryonic development,heat tolerance,hypoxia,reptile
1.Introduction
Unlike endotherms that maintain high body temperature via internal thermogenesis (Angillettaet al.,2010),most ectothermic animals behaviourally thermoregulate to utilize environmental heat sources,and thereby achieve optimal body temperatures that maximize their functional performance(Angillettaet al.,2002).Behavioural thermoregulation by ectotherms is very important for their daily activity,but may be affected by a number of biotic and abiotic factors(e.g.food,competition,predation and oxygen availability)(Herczeget al.,2008;Rusch and Angilletta,2017;Wood and Glass,1991).For example,many ectotherms select lower body temperatures as an adaptive response to protect vital organs under hypoxic stress (Wiggins and Frappell,2002;Wood and Glass,1991).Exploring the interaction of ecological factors on behavioural thermoregulation is critical for understanding how ectotherms cope with unpredictable environmental change.
Behavioural thermoregulation not only occurs at postembryonic stages,but also occurs in embryos of oviparous amniotes(Duet al.,2011;Liet al.,2014;Yeet al.,2021;Zhaoet al.,2013b).The adaptive significance of this behaviour was under debate(Corderoet al.,2018;Du and Shine,2022;Shine and Du,2018;Telemecoet al.,2016).Thermoregulatory behavior is expected to be conducive for successful embryonic development (Duet al.,2011).A recent study showed that behavioral thermoregulation by turtle embryos influenced offspring sex ratio,which expands the range of nest temperatures producing an equal sex ratio (Yeet al.,2019).Nonetheless,the adaptive significance of this behavior warrants further studies.
In the Chinese three-keeled pond turtle(Mauremys reevesii),when eggs were exposed to lateral heating,embryos moved towards a heat source at an optimal developmental temperature of 30°C,but moved away from a heat source at a stressful temperature of 33°C(Zhaoet al.,2013b).Given that the thermal preference of adult ecotherms can be lowered on exposure to hypoxia (Wood and Glass,1991),we hypothesize that reptilian embryonic thermoregulatory behaviour may also be affected by oxygen availability.Further,we predict that embryos will select lower body temperatures when exposed to hypoxia than when exposed to normoxia and hyperoxia.To test this hypothesis,we conducted a manipulation experiment with a two-factor design [three oxygen levels (hypoxia,normoxia,and hyperoxia) × two thermal environments (optimal and high temperatures)]to identify the effects of oxygen availability and temperature on the thermoregulatory behaviour of freshwater turtle embryos.
2.Material and Methods
2.1.Study species and egg collectionThe Chinese threekeeled pond turtle is a freshwater turtle distributed in southern and central China as well as southeastern Asia.Females lay eggs at the bank of rivers or ponds with vegetation cover from May to August.Mean nest temperatures of this species at semi-natural breeding sites range from 25.7°C to 30.1°C depending on the season(Yeet al.,2019).In 2016,we collected a total of 112 fertilizedM.reevesiieggs from a private turtle farm in Zhejiang,China.These eggs were weighed(mean±SD,egg mass=7.357±0.932 g),measured(mean±SD,egg length=32.88 ± 2.15 mm),and then incubated in boxes (220 mm × 100 mm × 80 mm) with moist vermiculite(-220 kPa,1 g water/1 g vermiculite,Zhaoet al.,2013a)at a constant room temperature of 28°C in normoxic conditions(21%O2).
2.2.Experimental design and thermoregulation behaviorWe commenced the following experiments when embryos had reached developmental stages 16-17(Greenbaum,2002).First,we located the initial position of an embryo in each egg by candling and then marked the position on the eggshells (Zhaoet al.,2013b).The eggs were then incubated individually in small plastic boxes filled with moist vermiculite (-220 kPa).We randomly assigned these boxes to the six treatments with three oxygen levels[hypoxia(12%O2),normoxia(21%O2)or hyperoxia(30%O2)]and two lateral heating treatments(30°C and 33°C,optimal and high temperature temperatures for embryonic development,respectively).We sealed plastic boxes with eggs into a 30 L polybag,which was supplied with gas containing different levels of oxygen.The polybags were then exposed to lateral heating of 30°C or 33°C.To achieve egg-surface temperatures of 30°C or 33°C,we adjusted the distance between the electronic heating mats (45 watt,300 mm ×700 mm)and the polybags accordingly(Zhaoet al.,2013b).After seven days during which the distance of embryo movements is substantial and readily measured (Zhaoet al.,2013b),the final position of each embryo was detected by candling.We recorded the distances between the initial and final positions of an embryo along the long axis of the egg with a vernier caliper (± 0.01 mm).The distance embryos moved towards or away from a heat source was used as a measure of the behavioural thermoregulatory ability of embryos.
During incubation,oxygen concentrations in the gas polybags were achieved by mixing oxygen with nitrogen.The gas in each polybag was renewed every day to maintain the desired oxygen concentration.During gas renewal,freshwater was added to the vermiculite to maintain the water potential of moist vermiculite.The process of gas renewal and freshwater addition was completed in under 2 minutes.We sampled gas from polybags periodically before and after each renewal.The sampled gas was checked by O2and CO2analyzers(FC-10A and CA-10A;Model TR3,Sable Systems,Henderson,NV,USA)with O2and CO2sensors(UI-2;Model TR3,Sable Systems,Henderson,NV,USA).During the experiment,the O2level of the gas in each polybag was kept within 2% of the three required treatment values (12%,21%or 30%O2),and the CO2level was kept below 0.2%.
2.3.Data analysisWe used two-way ANOVAs to analyze the effect of lateral heating treatment and oxygen concentration on the distance of embryonic movements towards or away from the heat source.Given the significant interaction between heating treatment and oxygen concentration,ANOVAs were used to analyze the effect of oxygen concentration on embryonic movement in each thermal treatment,and post-hoc tests (Tukey's test) were used to identify differences among oxygen treatments.
3.Results
The distances moved by embryonic Chinese three-keeled pond turtles were significantly affected by temperature(F1,106=28.996,P<0.0001),oxygen concentration (F2,106=25.008,P<0.0001),and the interaction between temperature and oxygen availability (F2,106=3.084,P<0.05).In general,embryos from the 30°C treatment moved greater distances towards the heat source than those from the 33°C treatment,which did not move as far or as close to the heat source(Figure 1).The distance that embryos moved towards the heat source decreased as oxygen concentration declined in both lateral heating treatments of 30°C(F1,56=10.061,P<0.001)and 33°C(F1,50=21.814,P<0.0001).Specifically,in the 30°C treatment embryos exposed to normoxia and hyperoxia moved towards the heat source,but those exposed to hypoxia did not;in the 33°C treatment embryos exposed to hyperoxia moved towards the heat source,but those exposed to hypoxia and normoxia moved away from the heat source (Figure 1).Therefore,the distance embryos moved towards a heat source decreased as oxygen concentration declined in both lateral heating treatments of 30°C and 33°C.
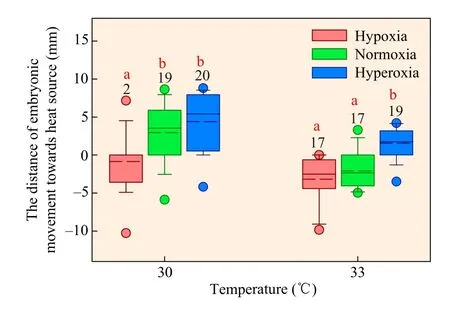
Figure 1 Effects of oxygen concentration and thermal treatment on the behavioural thermoregulation of embryos in the Chinese three-keeled pond turtle (Mauremys reevesii).Hypoxia,normoxia,and hyperoxia are 12%,21% and 30% oxygen concentrations,respectively.The numbers above the error bars are sample sizes.Means with different alphabets above the error bars are statistically different among different oxygen concentrations in each temperature treatment.The dots below and above the boxes are outliers.
4.Discussion
Our study concurs with the conclusion from a previous study(Zhaoet al.,2013b)that turtle embryos at normal oxygen levels move towards a heat source when exposed to an optimal temperature of 30°C,but move away from a heat source when exposed to a high temperature of 33°C.More importantly,consistent with our prediction,we demonstrated that oxygen availability during development significantly affects behavioural thermoregulation by turtle embryos;embryos exposed to hypoxia did not move as close to a heat source and,therefore,selected lower temperatures than embryos exposed to normoxia or hyperoxia in both thermal treatments.
This phenomenon of hypoxia-induced hypothermia has also been seen in post-embryonic stages of ectothermic animals (Petersenet al.,2003;Wiggins and Frappell,2002;Wood and Glass,1991).In adult reptiles,downregulation of their thermoregulatory setpoint is triggered by the release of adenosine due to a decrease in arterial oxygen(Petersenet al.,2003).The benefits of the regulated decrease in body temperature under hypoxia include improving oxygen loading in the lungs,reducing energetically costly cardiac hyperactivity,and decreasing oxygen consumption given the thermal sensitivity of metabolism(Sunet al.,2015;Wanget al.,1998).Therefore,embryonic selection of lower temperatures under hypoxia is a beneficial strategy that may alleviate the negative impact of hypoxic stress on development.
Reptile eggs in a nest likely experience hypoxia when unpredictable climate events such as excess rainfall and flooding occur,and when metabolism of microbes and eggs depletes the oxygen within the nest chamber(Ackerman and Lott,2004;Seymouret al.,1986).When facing hypoxia,reptile embryos may increase chorioallantoic membrane vascular density,shunt blood flow away from non-exchanging areas,increase cardiac output and carrying capacity of blood O2,or enlarge the opaque white patch (Corona and Warburton,2000;Tanget al.,2018).In addition to these physiological adjustments in response to hypoxia,our study demonstrates the adaptive response of embryonic thermoregulatory behaviour to hypoxia in turtles,expanding our knowledge of the capabilities that embryos possess to cope with unpredictable environments.
Hypoxia decreases the heat tolerance of ectotherms,because decreasing oxygen supply cannot meet the increasing demands of oxygen consumption when environmental temperatures increase,as suggested by the hypothesis of oxygen-and capacity-limited thermal tolerance(Pörtner and Knust,2007).Recent studies have demonstrated that oxygen supply significantly affects heat tolerance of reptile embryos,including turtles and lizards (Lianget al.,2015;Smithet al.,2015).Our results suggest that hypoxia not only decreases heat tolerance,but also the thermal preference of embryos.However,further studies are needed to fully understand the interactive effects of oxygen availability and temperature on the behaviour and physiology of ectothermic embryos.
AcknowledgementsWe thank Bo ZHAO,Teng LI,Liang MA and Baojun SUN for their help in the laboratory.This work was supported by grants from the National Natural Science Foundation of China(32030013,31821001).
 Asian Herpetological Research2023年4期
Asian Herpetological Research2023年4期
- Asian Herpetological Research的其它文章
- Next-generation Sequencing of MHC Class I Genes Reveals Trans-species Polymorphism in Eutropis multifasciata and Other Species of Scincidae
- Sexual Dimorphism,Female Reproductive Characteristics and Embryonic Thermosensitivity in the Tonkin Forest Skink (Sphenomorphus tonkinensis)from Hainan,South China
- Evidence for Expensive Tissue Hypothesis in the Asian Common Toad(Duttaphrynus melanostictus)
- Redefinition of the Odorrana versabilis Group,with a New Species from China (Anura,Ranidae, Odorrana)
- A New Species of the Genus Nanorana Günther,1896 (Anura:Dicroglossidae) from Hengduan Mountains of China
- Response of Distribution Range Against Climate Change and Habitat Preference of Four National Protected Diploderma Species in Tibetan Plateau