
A New Species of the Genus Nanorana Günther,1896 (Anura:Dicroglossidae) from Hengduan Mountains of China
2024-01-02 06:49:26FeirongJIShengchaoSHIShunMAChengSHENLimingCHANGandJianpingJIANG
Asian Herpetological Research 2023年4期
Feirong JI ,Shengchao SHI ,Shun MA ,Cheng SHEN ,Liming CHANG and Jianping JIANG,3*
1 Chengdu Institute of Biology,Chinese Academy of Sciences,Chengdu 610041,Sichuan,China
2 University of Chinese Academy of Sciences,Beijing 100049,China
3 Mangkang Biodiversity and Ecological Station,Tibet Ecological Safety Monitor Network,Changdu 854500,Xizang,China
Abstract An integrative study based on morphological data and mitochondrial molecular data of all known Nanorana(sensu stricto)species have revealed a new species from the Hengduan Mountains, N. huangi sp.nov.The new species can be distinguished from its congers by its: (1)medium body size,SVL 31.3-40.6 mm in adult males (n=12),39.2-48.1 mm in adult females(n=8);(2) absence of subarticular tubercles on fingers,with supernumerary tubercles below the base of the fingers small but distinct;(3)presence of dark dorsal patches without pale margins,generally creamy yellow ventral body basically,bright yellow ventral limbs;(4) small but prominent tympanum;(5) tibio-tarsal joint that reaches to the tympanum or the shoulder when hindlimbs adpressed forward towards body;(6)head length more than 30% of SVL.In addition,the new species can be separated from all other congeners by uncorrected genetic distances:ranging from 2.8%-7.5% of the mitochondrial 12S rRNA,1.1%-2.1%of the 16S rRNA,and 7.5%-11.5% of the COI.
Keywords Hengduan Mountains,Nanorana huangi sp.nov.,new species,taxonomy
1.Introduction
The genusNanoranawas established by Gunther(1896)withNanorana pleskeidesigned as the type species.Boulenger(1920)arranged it as a subgenus of the genusRana,i.e.,Rana(Nanorana) according to its external morphological and skeletal characteristics.Following a systematic study of the tailless amphibians in China,Liu and Hu (1961) revived the subgenus as a genusNanorana,which was monotypic at that time.Feiet al.(1985)reported a new species of this genus,N.ventripunctata,which is found in Zhongdian(now Shangri-La),Yunnan of China,adding one more species to the genus.
Stejneger(1927)named a new speciesAltirana parkeri,based on a specimen collected from Dingri in Xizang,China,and established the genusAltiranawith the new species as its type species.Dubois(1974)suggested that it should be a subgenus of the genusRana,i.e.,Rana(Altirana).Dubois(1992)redefined the classification system of the family Ranidae,reviving the genusNanoranaand classifying the genusAltiranaas a subgenus ofNanorana,i.e.,Nanorana(Altirana),However,Yeet al.(1993) and Zhao and Adler (1993) listed them as two distinct genera,i.e.,Nanoranacontained two known species andAltiranawith the one known species mentioned above.
After having analyzed the allele frequency of 12 isoenzymes inNanorana pleskei,N.ventripunctataandA.parkeri,Lȕ and Yang(1995)found a closer relationship between the latter two than any to the former.They suggested that the alpine frogA.parkeribelongs in the genusNanoranaand therefore treated it asN.parkeri.The generic level and its known species members ofNanoranawere then widely adopted by herpetologists(Cheet al.,2007;Dubois,1992;Duellma,1993;Fei,1999;Feiet al.,2005,2009,2020;Frost,2007;Jiang and Zhou,2001;Jianget al.,2005a,b,2020,2021;Matsui M.1996;Roleantal,2004;Zhao,1997).
The systematics status of the genusNanoranaunderwent continuous research and refinement until a consensus was finally reached.Dubois (1992) placedNanoranain the tribe Ranini of the subfamily Raninae in family Ranidae.On the basis of morphological and molecular systematics evidence,Jiang (1999) and Jiang and Zhou (2005) concluded thatNanoranashould be placed in the tribe Paini of the subfamily Dicroglossinae in family Ranidae,a view consistent with that of Roelantset al.(2004)and Feiet al.(2005).Frostet al.(2006)constructed the tree of life for Amphibians worldwide,adopting the family Dicroglossidae Anderson,1871,which includes two subfamilies Dicroglossinae Dubois,1992 and Occydozyginae Fei,Ye,and Huang,1990.The genusNanoranabelong to the subfamily Dicroglossinae.
Based on molecular phylogenetic research(Cheet al.,2010),a broad definition of the genusNanorana(sensu lato) was proposed,encompassing almost all species belonging to these groups (genera or subgenera):ChaparanaBourret,1939;PaaDubois,1975;UnculuanaFei,Ye,and Huang,1990;QuadranaFei,Ye,and Huang,1990(replaced byFeiranaDubois,1992);PaaDubois,1992;GynandropaaDubois,1992;MaculopaaFei,Ye,and Jiang,2010;OmbropaaDubois,Ohler,and Pyron,2021andDiplopaaDubois,Ohler,and Pyron,2021.Feiet al.(2010)still insisted on aNanorana(sensu stricto)based on its monophyly inferred from 4 nuclear genes (Rhodopsin,Tyrosinase,Rag-1,Rag-2) (Cheet al.,2010) and morphological characteristics.We adopt the sensu strictoNanoranain this study,despite Frost (2023) using sensu latoNanorana.To date,theNanorana(sensu stricto) contains 4 species:N.parkeri,N.pleskei,N.ventripunctata,andN.bangdaensis(Rao,2022 “2020”),all of which are endemic to the Qinghai-Tibet Plateau.Nanorana parkeriis distributed in eastern and southern Xizang,China,and the northern high mountains of Nepal at elevations of 2 850-5 000 m a.s.l.;Nanorana ventripunctatais distributed in northwestern Yunnan at elevation of 3 120-4 100 m a.s.l.;Nanorana pleskeiis distributed in southern Qinghai and Gansu,western Sichuan,and eastern Xizang at elevations of 3 300-4 500 m a.s.l.;andN.bangdaensisis found from eastern Xizang at elevations of 4 000-4 500 m a.s.l.(Feiet al.,2009;Cheet al.,2020;Rao,2022“2020”).
The differentiation and evolution ofN.parkeripopulations have been researched in detail based on molecular evidence(Liuet al.,2015;Wanget al.,2018),and this species was recognized in two clades,western and eastern,the latter one was described as a new species,i.e.,N.bangdaensis,by Rao(2022 “2020”).Similar work onN.pleskeiwas conducted by Zhouet al.(2016) and Wanget al.(2017),three clades were discovered,southern,northern and western(Zhouet al.,2016;Wanget al.,2017).However,morphological studies between different clades were deficient (Liuet al.,2015;Zhouet al.,2016;Wanget al.,2017;Wanget al.,2018).
Recently,we conducted an extensive field survey and collected samples ofNanoranafrom Yunnan from 2019 to 2022.We found that the specimens newly collected from Lanping County and Dali City in northwestern Yunnan represent a distinct form compared toN.pleskei,N.parkeri,N.bangdaensis,andN.ventripunctata,as inferred from morphological and molecular data.Therefore,we describe it as a new species herein.
2.Material and Methods
2.1.SamplingA total of 86 specimens of the genusNanorana(sensu stricto) were newly collected in this study(Table 1),including 11 specimens ofN.ventripunctata,10 specimens ofN.parkeri,12 specimens ofN.pleskei,23 specimens ofN.bangdaensis,and 30 unnamedNanoranaspecimens from Lanping and Dali.
Following euthanasia (paralysed them with a low concentration of clove bud oil),all the specimens were fixed in 75%ethanol.Tissue samples were taken and preserved in 95%ethanol prior to fixation and stored at-20°C for later use.All specimens were deposited in the Chengdu Institute of Biology(CIB),and the Chinese Academy of Sciences(CAS).
2.2.Molecular phylogenetic analysesGenomic DNA was extracted from muscle tissue or liver tissue using a DNA extraction kit (Tiangen Biotech Beijing Co.,Ltd).Three mitochondrion genes namely the 12S ribosomal RNA gene(12S),16S ribosomal RNA gene (16S) and cytochrome C oxidase subunit I(COI),were amplified.For12S,the primers FPhe40L and 600H were used following(Simonet al.,1994),for16S,the primers 16SP7 and 16SP8-R were used following(Zhanget al.,2013),and forCOI,the primers Anurans-COIChmf 4 and Anurans-COI-Chmr4 were used following(Cheet al.,2012).Amplification of the12S,16SandCO1fragments was performed in a 25 μL volume reaction,with an initial denaturing step at 95°C for 4 min;35 cycles of denaturing at 95°C for 30 s,annealing at 55°C(for12S/16S)/52°C(forCOI)for 30 s and extending at 72°C for 30 s,and a final extending step of 72°C for 7 min.PCR products were sequenced with both forward and reverse primers,utilizing the same primers as those used in the PCR process.Sequencing was conducted using an ABI3730 automated DNA sequencer from Chengdu Youkang Jianxing Biotechnology Co.,Ltd.(Chengdu,China).New sequences were deposited in GenBank (for GenBank accession numbers see Table 1).
Other than the sequences of the three gene segments,including 410 bp of12S,537 bp of16S,and 550 bp ofCO1,from 86 samples in this study,the same segments of an additional 21 samples were downloaded from Genbank(Table 1) for phylogenetic analysis.Four species (Gynandropaasichuanensis,Maculopaa chayuensis,M.maculosa,andQuasipaa boulengeri)were used as outgroups following Jianget al.(2005),Cheet al.(2010),and Qiet al.(2019).
The12S,16SandCO1sequences were input to MEGA11(Tamuraet al.,2021)and aligned by MUSCLE(Edgar,2004).Uncorrected pairwise distances (P-distance) were then calculated.IQ-TREE 1.6.12 was used to conduct the maximum likelihood (ML) analysis (Nguyenet al.,2015).The best-fit model HKY+F+G4 was selected by Modelfinder according to BIC (Kalyaanamoorthyet al.,2017) for12SandCO1,while K2P+I was the best-fit model for16S.Two analyses were performed,one for all species,and one for species for which we had data for all three genes.The12S,16SandCO1sequences were concatenated seriatim into a 1497 bp single partition.The best partitioning scheme and evolutionary models for these 3 pre-defined partitions were selected using PartitionFinder2 (Lanfearet al.,2017),with greedy algorithm and AICc.GTR+I+G was found to be the best-fit model of evolution for both the12Sand16S,and HKY+G was found to be the best-fit model of evolution forCO1.Ultrafast Bootstrap Approximation(UFB)was used to assess node support,using 10000 ultrafast bootstrap replicates with values of UFB(%)≥95 considered as significantly supported(Hoanget al.,2018).Single branch tests were conducted using SH-like approximate likelihood ratio test(SH-aLRT)via 1000 replicates,and values of SH (%) ≥80 were deemed as significantly supported(Guindonet al.,2010).
Bayesian inference(BI)was conducted using MrBayes 3.2.6(Ronquistet al.,2012)under partition model(2 parallel runs,2000000 generations),sampled every 1000 generations with the first 25% of samples discarded as burn-in,resulting in a potential scale reduction factor (PSRF) of ≤0.005.Bayesian posterior probabilities (BI) ≥ 0.95 were considered significantly supported.
2.3.Morphological analysesThe newly collected specimens were morphologically compared with the other congeners on the basis of descriptions in literatures and specimens deposed in CIB.
Measurements of adultsAll measurements were taken with digital calipers to the nearest 0.1 mm.Following Feiet al.(2009)and Qiet al.(2019),the following morphometrics were measured:
Body and head SVLsnout-vent length,from tip of snout to vent;AGaxilla to groin,distance from posterior base of forelimb at its emergence from body to anterior base of hindlimb at its emergence from body;HLhead length,from posterior corner of mandible to tip of snout;HWhead width,at the greatest cranial width;HHhead height,greatest height of head;SLsnout length,from tip of snout to the anterior corner of eye distance;SNDsnout to nostril distance,distance from tip of snout to nostril;ENDeye to nostril distance,distance from anterior corner of eye to nostril;IBEDinternal back of eyes (the shortest distance between the posterior borders of the orbits);IFEDinternal front of eyes (shortest distance between the anterior borders of orbits);INDinternasal distance,distance between nostrils;IODinterorbital distance,least distance between upper eyelids;UEWupper eyelid width,maximum width of upper eyelid;EHDeye horizontal diameter;HTDhorizontal diameter of tympanum;VTDvertical diameter of tympanum;TEDtympanum-eye distance (distance from the anterior border of the tympanum to the posterior orbital border).
Forelimbs LALlength of lower arm,from proximal end of outer metacarpal tubercle to elbow joint;LADdiameter of lower arm;HALhand length,from proximal end of outer metacarpal tubercle to tip of the finger III.
Hindlimbs HLLLength of hindlimb:from tip of disk of toe IV to vent;FMLfemur length;TILtibia length;TFLlength of tarsus and foot,from proximal end of tarsus to tip of the toe IV;FLfoot length,from proximal end of inner metatarsal tubercle to tip of toe IV;IMTLlength of the inner metatarsal tubercle.
20 individuals of the undescribed species,11 individuals ofN.pleskeiand 10 individuals ofN.ventripunctata,were measured.
Measurements of tadpolesThree newly collected tadpoles of the undescribed species were measured.The stages of tadpoles were 29,39,and 40,respectively,identified following Gosner (1960).All measurements were taken with digital calipers to the nearest 0.1 mm.Following Feiet al.(2009),seventeen tadpole morphometric characters were measured:TOLtotal length;SVLsnout-vent length(from the tip of the snout to the posterior edge of the anus);TLtail length(from the base of anal tube to the tip of the tail);BHbody height(the highest region between the dorsal and ventral surfaces of the body);BWbody width(largest width between the sides of the body);SNDsnout-narial distance(from the tip of the snout to the center of the naris);NEDnaris-eye distance(distance from center of naris to anterior corner of eye);SLsnout length,from tip of snout to the anterior corner of eye distance;SSDsnout-spiracle distance (distance from tip of snout to opening of spiracle);EDmaximum eye diameter;INDinternasal distance(distance between center of two naris);LTFHmaximum height of lower tail fin;ODWoral disc width (largest width of oral disc);SUFDsnout-upper fin distance (distance from snout to beginning of upper tailfin);TMWmaximum tail muscle width;TMHmaximum tail muscle height;UTFHmaximum height of upper tail fin.
All statistical analyses were performed using the Origin 2023 software.In order to reduce the impact of allometry,the correct value for each character's ratio to SVL was calculated and then was log-transformed for subsequent morphometric analyses.Mann-WhitneyUtests were conducted to test the significance of differences on morphometric characters between the undescribed species,N.pleskeiandN.ventripunctata.The significance level was set at 0.05,P≤0.05 indicates significant difference.Furthermore,principal component analyses (PCA)were conducted to highlight whether the different species were separated in morphometric space.
The new taxon was also compared with all other congeners ofNanorana(sensu stricto)based on morphological characters(Table 2),which were obtained from specimens (Appendix)and literature sources.
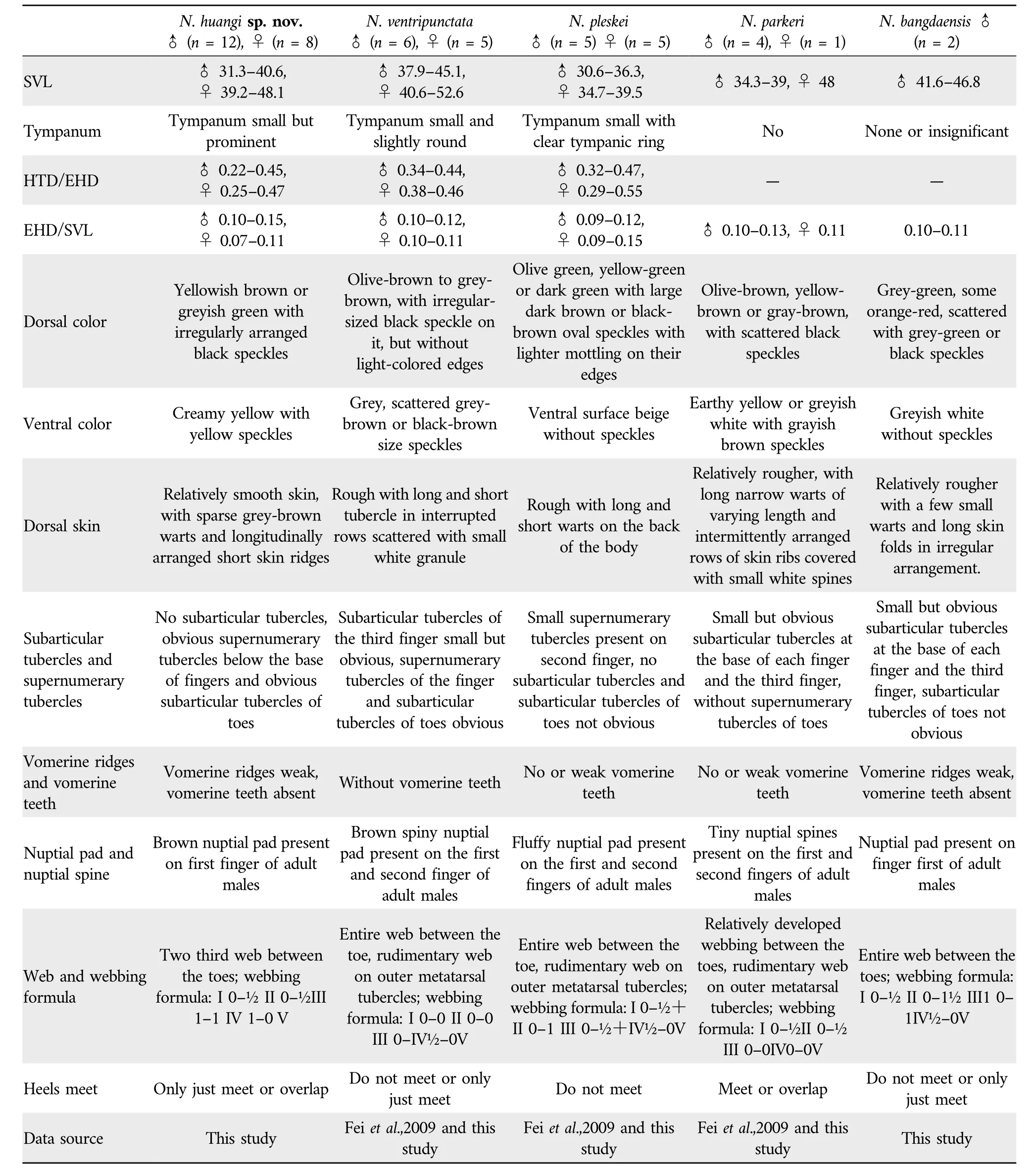
Table 2 Morphological comparisons among the N. huangi sp.nov., N. ventripunctata, N. pleskei, N. parkeri,and N. bangdaensis.
3.Results
3.1.Molecular phylogenetic analysisFour ML phylogenetic trees were constructed inferred from12S,16S,CO1and12S+16S,respectively,and they have approximately the same topology(Figure S1).BI and ML phylogenetic trees were constructed based on the concatenated mitochondrial12SrRNA,16SrRNA andCO1DNA sequences with a total length of 1 497 bp,and they had largely identical topologies(Figure 1),also similar to the four ML tree above.Here,just the tree of Figure 1B was used to illustrate their phylogenetic relationships.All samples ofNanoranawere clustered together as one monophyletic group,the newly collected unnamed specimens clustered together as one monophyletic group with well supporting values (SH 99/UFB 99/BI 1),and formed a sister group to the clade composed byN.ventripunctataandN.pleskei(SH 83/UFB 85/BI 0.99),indicating that these new specimens may belong to a different taxon to the latter two.In addition,the uncorrectedP-distance between the group composed by these new specimens and its congeners are 2.8%-7.5%inferred from12S,1.1%-2.3%from16S,and 7.5%-11.5% fromCO1,which are larger than some of the ones among the remaining species(Table 3).For12S,the minimum genetic distance between the new species and the other four species was 2.8 %,while the minimum genetic distance between the other four species were 1.5%(similarly16S:1.3%vs 1.1%;CO1:7.5%vs 3.3%).Therefore,molecular systematics indicate that these newly collected specimens (Lanping-Dali population)may represent an undescribed species.
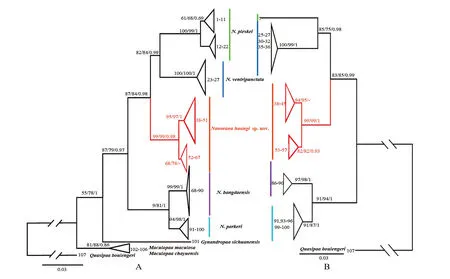
Figure 1 Maximum likelihood tree of the genus Nanorana based on the sequences of the mitochondrial 12S,16S,and CO1 gene.A:Tree of all studied specimens (some of them have not all the three genes’sequence),B: Tree of those specimens with the three genes’sequences.The supporting values of tree nodes on the branches:SH-like approximate likelihood ratio test(SH,%),Ultrafast Bootstrap Approximation(UFB,%),and Bayesian posterior probabilities (BI),the ones lower than 50 are displayed as “-”.
3.2.Morphological analysisThe Lanping-Dali population is distinctly different fromN.ventripunctata,N.pleskei,N.parkeri,andN.bangdaensisby having smooth skin,sparse graybrown warts and short vertical rows of skin ribs;being medium in size and having no subarticular tubercles with obvious supernumerary tubercles below the base of the fingers(Figure 2).Although it is most similar toN.ventripunctata,andN.pleskei,the Lanping-Dali population is also morphological distinct from them (Table S1).Ten characters were significantly different between Lanping-Dali population andN.ventripunctata: SVL,AG/SVL,UEW/SVL,IFED/SVL,IBED/SVL,TED/SVL,SND/SVL,LND/SVL,FML/SVL,FL/SVL;five characters were significantly different between Lanping-Dali population andN.pleskei:HH/SVL,UEW/SVL,IFED/SVL,SND/SVL,END/SVL;and eight characters were significantly different betweenN.ventripunctata.andN.pleskei:SVL,AG/SVL,SL/SVL,SND/SVL,LAD/SVL,HAL/SVL,FL/SVL (Table S1).These differences are more pronounced in male individuals than in females.In the PCA analysis(Figure 3),both male and female adults can be differentiated between the two species,but males show more distinct separations inN.huangisp.nov.andN.ventripunctata.Despite having a sympatric distribution,these two species can be readily distinguished based on their morphology.

Figure 2 Comparisons of morphological characters among Nanorana huangi sp.nov.(A),N.ventripunctata(B),N.pleskei(C),N.parkeri(D),and N. bangdaensis (E).1:dorsolateral view,2:ventral view,3:side view,4:hand ventral view,5: foot ventral view,6: lateral head.
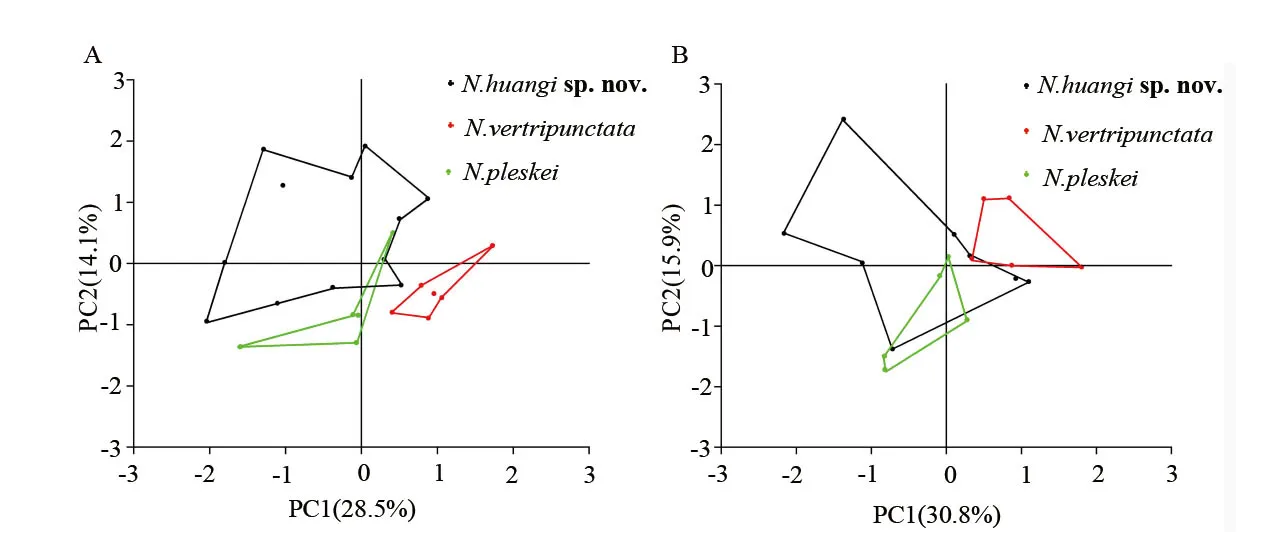
Figure 3 PCA plot for morphometrics of N.huangi sp.nov.,N. ventripunctata,and N.pleskei.A: male adults,B: female adults.

Figure 4 The holotype specimen CIB YN201909255 of Nanorana huangi sp.nov.A:dorsal view,B:ventral view,C:lateral view of head,D:ventral view of hand,E: ventral view of foot.
3.3.Taxonomic conclusion and taxonomic accountWith the combination of the molecular phylogenetic results and morphological differences above,the Lanping-Dali population ofNanoranafrom northwestern Yunnan should represent a new species and we describe it herein.
Taxonomic account
Nanorana huangisp.nov.(Figures 2,4,5,and 6)
https://zoobank.org/B7C0B475-687C-406F-9C60-31013476F028
ChresonymyNanorana pleskei: Zhaoet al.(1997).Nanorana ventripunctata:Population from Weixi,Yunnan Province in Feiet al.(2009,2012)and Fei,(2020).
HolotypeCIB YN201909255,adult male,collected by Shengchao Shi,Wenbo Zhu,and Lei Fu on 19 September,2019,from Jinghuadian wetland,Lanping County,Nujiang Prefecture,Yunnan Province,China (26.874593°N,99.544008°E;3 389 m a.s.l.),and preserved at Chengdu Institute of Biology,Chinese Academy of Sciences(CIB,CAS).ParatypesThree adult males (CIB YN201909256-CIB YN201909258),five adult females (CIB YN201909083,CIB YN201909084,CIB YN201909086,CIB YN201909226,CIB YN201909276),and two subadults (CIB YN201909085,CIB YN201909259) were collected along with the holotype,elevations range from 3 389-3 455 m.One adult female(CIB YN201909226)and one adult male(CIB YN201909227)and were collected on the same night from Dayangchang,Yulong County,Lijiang Prefecture,Yunnan Province,China.These two sites share the same wetland bordering Lanping County and Yulong County.Seven adult male specimens(CIB XZ2022268-CIB XZ2022270,CIB XZ2022272-CIB XZ2022273,CIB XZ2022278,CIB XZ2022282) and three adult females (CIB XZ2022271,CIB XZ2022275,CIB XZ2022280)were collected by Liming Chang,Cheng Shen,and Dequn Zhang on 15 July,2022,from Huadianba in Xizhou Town,Dali City,Yunnan Province,China(25.857178 °N,100.029178 °E;2900 m a.s.l),and preserved at Chengdu Institute of Biology,Chinese Academy of Sciences(CIB,CAS).
EtymologyThe new species is named after the Chinese herpetologist Yongzhao HUANG 黄永昭for his contribution to herpetological taxonomy research in the Qinghai-Tibet Plateau.The specific nomen “huang” also means yellow in Chinese,which also refers to the ventral coloration of the new species.For convenience in identification,we suggest the Chinese name “黄腹倭蛙” (huáng fù wō wā) for its yellow ventral coloration.The English common name is suggested as“Yellow-bellied Plateau Frog”.
DiagnosisNanorana huangisp.nov.is distinguished from its congeners by a combination of the following morphological characters: (1) medium body size,SVL 31.3-40.6 mm in adult males (n=12),39.2-48.1 mm in adult females(n=8);(2)absence of subarticular tubercles on fingers with supernumerary tubercles below the base of the fingers small but distinct;(3) presence of dark dorsal patches without pale margins,generally creamy yellow ventral body,bright yellow ventral limbs;(4)small but prominent tympanum;(5) tibio-tarsal joint that reaches to the tympanum or the shoulder when hindlimbs adpressed forward towards body;(6) head length more than 30% of SVL.
Description of holotypeAdult male,medium body size,SVL 31.3 mm(Table S2).
Head:Head wider than length(HL11.5 mm,HW 12.4 mm),snout bluntly rounded and protruding from lower lip,canthus rostralis obvious,nostril closer to eye than snout (SND 2.5 mm,END 1.8 mm),distance between nostrils slightly larger than that between upper eyelids (ID 2.4 mm,IUE 1.8 mm);tympanum rounded,small but prominent,diameter smaller than half of eye diameter(EL 1.2 mm,TYD 3.7 mm),and slightly larger than eye-tympanum distance (TED 1.1 mm);tympanum rounded,slightly raised above surrounding areas,upper edge concealed with supratympanic ridge;interorbital space flat,wider than upper eyelid (UEW 3.4 mm);vomerine ridges weak,vomerine teeth absent.No vocal sac,tongue weakly notched,medial lingual process absent.
Forelimbs:Forearm moderately long and shorter than hand,forearm and hand length less than half of SVL;fingers strong,without webbing and lateral fringes,supernumerary tubercles present on finger III and IV,absent on other fingers,base of fourth finger distinctly larger than the third one,finger length formula IV <III <II <I;inner metacarpal tubercles slightly raised above hand;tips of fingers slightly enlarged;nuptial pad present.
Hindlimbs: Relatively long and stout,tibio-tarsal articulation reaches tympanum when hindlimb adpressed forward to body;thighs slightly shorter than shanks (FML 12.0 mm,TIL 13.2 mm);toes long,with narrow lateral fringes,tips rounded and not expanded;toes relative lengths I <II <V<III <IV,webbing between toes well developed,webbing formula: I 0-½ II 0-½III1-1IV1-0V;subarticular tubercles absent;outer metatarsal tubercle absent,inner metatarsal tubercle oval,small,situated at the base of first toes (IMTL 1.7 mm),tarsal fold absent.
Skin: Dorsal surface generally smooth,body with several sparsely distributed longitudinal short skin ridges from shoulder to rear,few small warts present on flanks and dorsal hindlimbs.Dorsolateral fold absent;supratympanic fold flat.Dorsal head,upper eyelids,lateral head and forelimbs smooth.Ventral body generally smooth,multiple tiny granules present on belly and skins around cloaca,size larger around cloaca.
Coloration in life(Figure 2) Dorsal surface yellowish brown,scattered with distinct dark patches on body and limbs,patches similar size to eye diameter with clear boundaries and without bright colored edges.One dark stripe present from tip of snout along canthus rostralis and supratympanic fold to corner of mouth on both lateral sides of head.Ventral body creamy yellow,ventral limbs bright yellow,ventral areas of hands and feet grey.Granules on ventral surface yellow,but white around cloaca.
Coloration of holotype in preservedThe background coloration of the dorsum fades to light grey,dark brown patches and stripes become more distinct.Ventral body and limbs white,ventral hand and feet grey.
Measurement of holotype(mm) SVL 31.3;AG 13.7;HL 10.0;HW 11.9;HH 5.8;SL 4.8;ID 2.4;IOD 1.9;UEW 1.8;IFED 4.2;IBED 7.1;EHD 3.7;HTD 1.2;VTD 1.2;TED 1.1;SND 2.5;END 3.7;LAL 5.4;LAD 3.5;HAL 7.9;HLL 44.0;FML 12.0;TIL 13.2;TFL 22.0;FL 15.7;IMTL 1.7.
Sexual dimorphismAdult females (SVL 39.2-48.1 mm,n=8)are larger than adult males(SVL 31.3-40.6 mm,n=12).Morphometric differences between adult males and females are summarized in Table 4.Lower arms of adult males are enlarged compared to upper arms;brown nuptial pad present on first finger of adult males;a pair of long elliptical “/ ”shaped chest gland present on the chest of adult males but without spines at the time of collection.
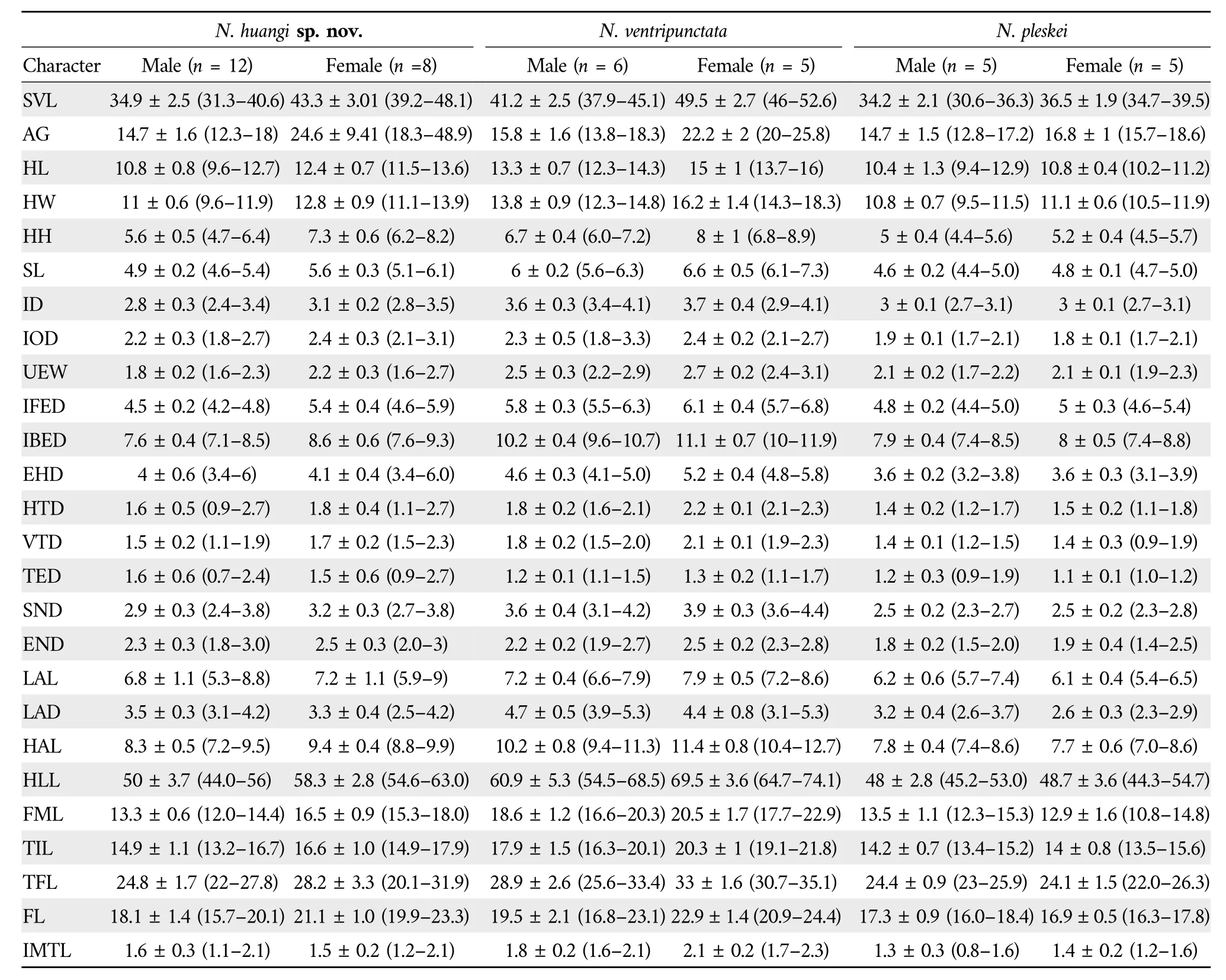
Table 4 Measurements [Mean ± SD (Range)] of Nanorana huangi sp.nov., N. ventripunctata and N. pleskei.
TadpolesThree tadpoles ofNanorana huangisp.nov.were collected at stages 29-40 (Gosner,1960) (Figure 5).The measurement data is summarized in Table 5.Description based on sequenced tadpole CIB YN201909089-1 at stage 40(BL=17.6):the body is elliptical in dorsal and ventral view;snout rounded in dorsal and lateral view;eyes dorsolaterally directed,pupils round;nares rounded,anterodorsally directed,snout-nostril distance longer than nostril-orbitals distance(SND/NE 1.4),without projections but with prominent marginal rim;spiracle short (SSD/BL 0.6);tail long and strong(TL/BL 1.9,TWH/BH 0.8,TMW/BW 0.3);upper fin,slowly rise from proximal eighth of tail,extending to tip of tail,highest at distal third and smooth;lower fin attached to anal tube,smooth,highest at distal third of tail,reaches tip of tail(LTFH/UF 1.2);Oral disc (Figure 5) relatively small,ellipse,positioned ventrally,not emarginated laterally (ODW/BL0.2,ODW/ BW 0.4);small papillae on margin around oral disc present above level of mouth corner,absent on lower region;KRF I: 2+2/Ⅲ.Dorsal surface of body and tail olive green,ventral surface lighter,and semitransparent,a light black patch present near middle dorsal tail,tail fin semitransparent scattered with grey pigments on upper fin.

Figure 5 The tadpole CIB YN201909089-1 in life of Nanorana huangi sp.nov.A:dorsal view,B:lateral view,C:ventral view,D:dorsal view of head,E: oral disc.
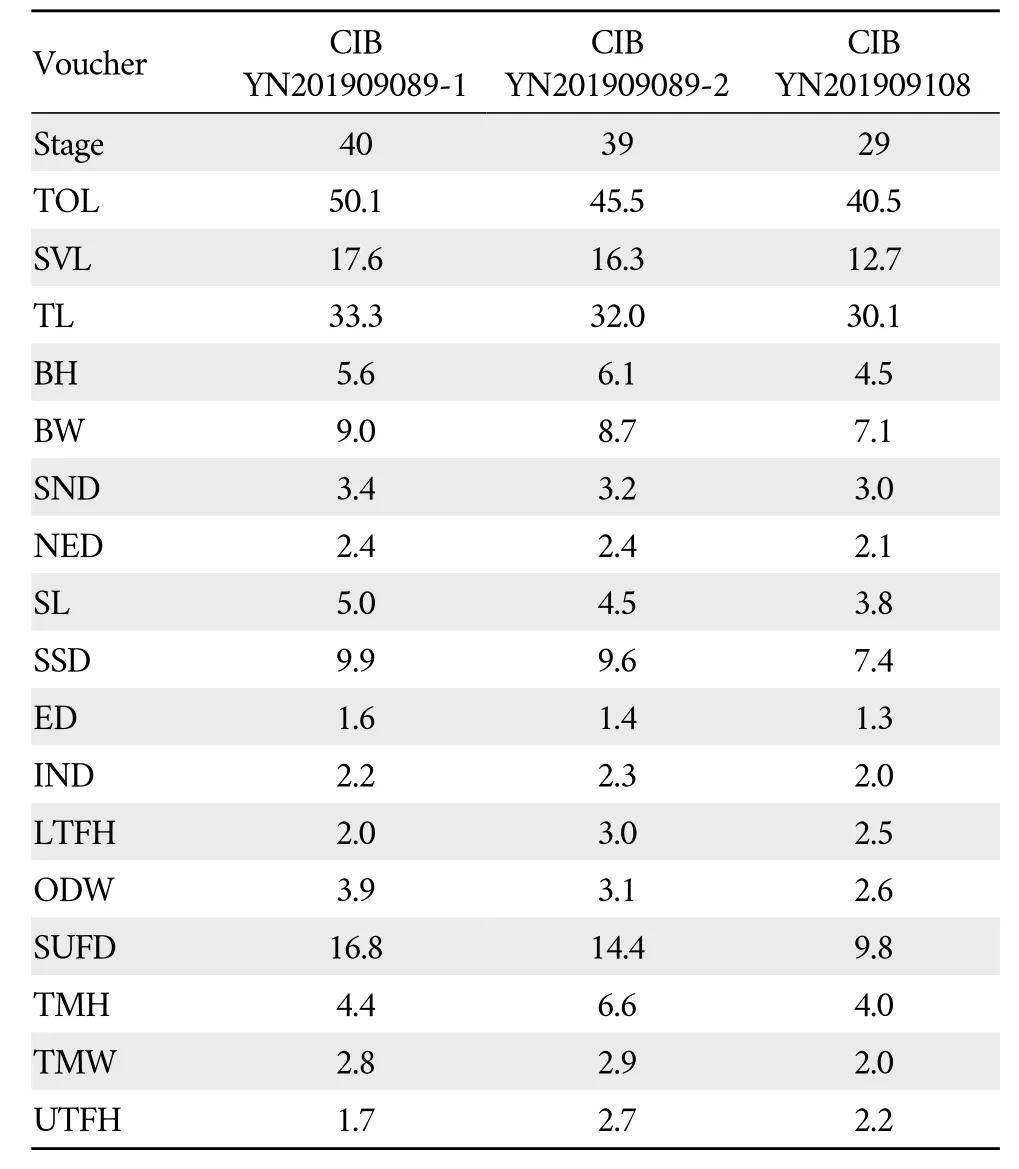
Table 5 Measurements of tadpoles of Nanorana huangi sp.nov.
Coloration of tadpole in preserveDorsal body brown,tail muscle pale and ventral parts light brown and fins translucent.
VariationParatypes generally similar to holotype in external morphology.Only few dark patches present on dorsum of adult female CIB YN201909276,and its belly cream white with few small dark speckles (Figure 6A).Dorsal surface of adult male CIB YN201909227 olive brown(Figure 6B).

Figure 6 Variation of Nanorana huangi sp.nov.in life.A:paratype(CIB YN201909226),adult female;B: paratype (CIB YN201909227),adult male.1: dorsolateral view,2:ventral view.
Distribution and ecologyAt present,Nanorana huangisp.nov.is found in an alpine grass wetland bordering Jinghuadian,Lanping County,Nujiang Prefecture and Dayangchang,Yulong County,Lijiang Prefecture,and Xizhou County,Dali Prefecture of Yunnan Province (Figure S2,Table 1).It was found in temporary puddles,ponds,or moist meadows of alpine wetland (Figure 7) at elevations of 2 900-3 885 m.The highest elevation record for the new species is from Houzhuping,Lanxiang Village,Yulong County,Lijiang Prefecture,Yunnan Province,China (99.717700 °E.26.630314°N,3885 m)by Song-Lin Shi.It is sympatric withRana chaochiaoensis,Rana shuchinae,Bufo andrewsi,Bombina maximaandOreolalax xiangchengensis.

Figure 7 Habitat of Nanorana huangi sp.nov.A-B:Dali,Yunnan,altitude 2 900 m;C-D:Lanping,Yunnan,altitude 3 389 m.
4.Discussion
Nanorana parkeri,N.pleskei,N.bangdaensis,andN.ventripunctataare endemic to the Tibetan Plateau,and have evolved a range of specialized adaptations for high-altitude habitats (Chenet al.,2011;Zhouet al.,2016;Wanget al.,2017;Wanget al.,2018;Cheet al.,2020).The description ofN.huangisp.nov.suggest that the species diversity of the genusNanorana(sensu stricto)may still be underestimated.
The distinct terrain of deep valleys has played a central role in shaping the genetic divergence ofNanorana parkeri(Wanget al.,2018).The identification ofN.huangisp.nov.further underscores the significance of valleys as geographical barriers in the speciation ofNanorana,implying that the Hengduan Mountains could potentially be the origin ofNanorana(sensu stricto).Notably,Zhao and Yang (1997) recorded the distribution ofN.pleskeiin Weixi,Yunnan,while Feiet al.(2009)suggested that those specimens may actually belong toN.ventripunctatabased on the geographical distribution.The distribution sites ofN.huangisp.nov.in Lanping County andN.pleskeiin Weixi County are situated on the same mountains and both lie on the eastern side of the Lancang River.The Jinsha River separates them from the main distribution areas ofN.ventripunctataandN.pleskei.Therefore,we speculate that theNanoranafrog in Weixi may actually beN.huangisp.nov.,though further work in the future would be needed to confirm this.
The new species is distributed in the southernmost parts of the current distribution range for the genusNanorana(sensu stricto).It expands the known distribution range by at least one latitude to the south in comparison with previously documented (Zhao and Yang,1997;Feiet al.,2009,2012),which holds significant importance for zoogeographical research.
Nomenclatural acts registrationThe electronic
version of this article in portable document format represents a published work according to the International Commission on Zoological Nomenclature (ICZN);hence,the new names contained in the electronic version are effectively published under that Code from the electronic edition alone (see Articles 8.5-8.6 of the Code).This published work and the nomenclatural acts it contains have been registered in ZooBank,the online registration system for the ICZN.The ZooBank Life Science Identifiers(LSIDs)can be resolved and the associated information can be viewed through any standard web browser by appending the LSID to the prefixhttp://zoobank.org/.Publication LSID: urn:lsid:zoobank.org:pub:d7936d56-cd21-48f5-ba3b-09ad79945132.Species LSID: see in Taxonomic accounts.
AcknowledgementsThis work is supported by the Second Tibetan Plateau Scientific Expedition and Research Program(STEP,Grant No.2019QZKK05010503),and China Biodiversity Observation Networks (Sino BON-Amphibian and Reptile).Field work was conducted under permission of the Forestry and Grassland Administration of Xizang Autonomous Region (No.2021[71]) and Forestry and Grassland Administration of Yunnan Province (No.2019[825]).We thank many people who provided us help during field trips,especially Zhen XIAO and Duozha from Forestry and Grassland Administration of Luozha County,Zhongchuan BAI from Lakang Town Government and Dazeng Gusang from Lajiao Town.We thank Xianguang GUO,Mingqiang WANG,Wenbo ZHU,Lei FU,Peng YAN,Jianyi FENG,Ningning LU,Zhongyi YAO,Shiyao DU from CIB,and Zhipang HUANG from Dali University for their help in field and lab work.We also thank Forestry and Grassland Administration of Yunlong County of Yunnan Province,and Qamdo Forestry and Grass Bureau of Xizang Autonomous Region,Mangkang Biodiversity and Ecological Station,Tibet Ecological Safety Monitor Network for help in field work.We thank Songlin SHI from Chengdu University of Technology for providing a location record of the new species.We thank Yin QI and Xiudong SHI for providing partial material and photographs ofN.ventripunctata.
 Asian Herpetological Research2023年4期
Asian Herpetological Research2023年4期
- Asian Herpetological Research的其它文章
- Response of Distribution Range Against Climate Change and Habitat Preference of Four National Protected Diploderma Species in Tibetan Plateau
- Redefinition of the Odorrana versabilis Group,with a New Species from China (Anura,Ranidae, Odorrana)
- Evidence for Expensive Tissue Hypothesis in the Asian Common Toad(Duttaphrynus melanostictus)
- Sexual Dimorphism,Female Reproductive Characteristics and Embryonic Thermosensitivity in the Tonkin Forest Skink (Sphenomorphus tonkinensis)from Hainan,South China
- Next-generation Sequencing of MHC Class I Genes Reveals Trans-species Polymorphism in Eutropis multifasciata and Other Species of Scincidae
- Oxygen Availability Affects Behavioural Thermoregulation of Turtle Embryos