
寒武纪腕足动物起源及冠轮动物谱系演化
2024-01-02 13:05张志飞胡亚洲
西北大学学报(自然科学版) 2023年6期
张志飞,梁 悦,刘 璠,胡亚洲,付 饶
(1.西北大学 地质学系/大陆动力学国家重点实验室 陕西省早期生命与环境重点实验室,陕西 西安 710069;2.中国科学院南京地质与古生物研究所,江苏 南京 210008)
生命是能够自我繁殖、自我复制的有机体,是地球区别于其他星球的重要标志。对于生命如何分类而言,早期林奈[1]在《自然系统》中将生命分为两界:动物界和植物界。后期随着分子生物学的进步,尤其是基因组学科的发展,Woese等人[2]根据核糖体RNA碱基序列比对,提出了生命三域分类框架,即将生命分为细菌域(Bacteria)、古菌域(Archaea)和真核生物域(Eukarya)。其中真核生物又分为真核单胞的原生生物和真核多胞的植物、真菌和动物。基于此,地球生命形成了三大域、五大界的生物分类方案[3-4]。真核生物的起源和早期演化是揭示地球生命演化奥秘的核心科学问题之一,并且一直都是生命科学和古生物学领域研究热点。研究人员在中国华北燕山地区古元古代晚期常州沟组和串岭沟组(距今17亿年)地层中发现丰富多样的真核生物化石,主要以单细胞球形藻类为主,也包含少量具刺类型、单细胞集合体和丝状体[3-4]。此外,含有微生物的真核生物群落在全球多处分布的页岩(16~14亿年)中被报道[5-6],但最早的属于冠群真核生物的化石是产出自加拿大硅化碳酸盐中的简单红藻细胞多聚体,其年龄大约约束在距今12~11亿年[7]。
前寒武纪—寒武纪界线是地球历史上的重要转折期,在经历了长达35亿年由微生物席主导的原始海洋生态系统之后,开始向持续5亿年之久的以后生动物为主导的显生宙海洋生态系统转变,主要表现在至少18个与现代动物门类发育体制构型(body-plan)相似或者基本相同的动物门类“幕式”快速出现在寒武纪早期的地层中,这一快速辐射事件被形象地称为“寒武纪生命大爆发(Cambrian Explosion)”[8-18]。寒武纪大爆发因为地球动物门类的突发性快速出现而备受关注,事关地球海洋动物生态系统的起源和建立,因此是地球宜居性演化的重大地质和生命事件。根据最新的研究成果,寒武纪生命大爆发历时大约4千万年,使得地球动物树三大亚界(基础动物亚界、原口动物亚界和后口动物亚界)、四大超门(基础动物、后口动物、冠轮动物和蜕皮动物)依次成型[8-9,19-20]。关于寒武纪大爆发两侧动物起源的研究,亮点工作主要集中在与现代节肢动物相关的蜕皮类以及与我们人类有重要亲缘关系的后口动物类群[8-9,19-20]。但是,与现代海洋生物礁体和海洋环境紧密相关的底栖(草根)动物——冠轮动物(腕足动物、苔藓虫、软舌螺、环节动物和软体动物)在特异型化石库明显较少,在寒武纪地层中多以“碎渣子、烂壳子”碎片状的化石出现,化石生物学(palaeobiology)研究实属困难。
冠轮动物(Lophotrochozoa)包括动物界几乎一半的动物门类,涵盖了所有的非蜕皮类原口动物。目前主要分为三个支系,即由软体动物、环节动物、纽形动物、腕足动物、帚虫动物和星虫动物门组成的担轮动物(Trochozoa),由环口动物、内肛动物和苔藓动物门组成的群居虫类(Polyzoa),以及由许多小的动物门类组成的扁形动物(Platyzoa)(见图1)。对于其不同门类之间的相互关系,不同学者在不同研究阶段意见不一,尤其是腕足动物与哪一动物类群的系统亲缘关系最近[21-24]。有学者认为腕足动物和帚虫动物为姊妹群,而苔藓动物位于触手冠动物类群之外[25];而系统基因学研究表明触手冠动物(腕足动物、帚虫动物和苔藓动物)共同构成了单系群[24,26-30](见图1)。由此可见,触手冠动物以及冠轮动物超群的基因组成和演化比之前认为的要复杂的多,其类群之间的相互关系极其不稳定。此外,分子生物学不能揭示生物躯体构型进化的模式和方向。幸运的是,保存在岩石中的大量化石和生物地质记录无疑为地球生命的演化历史和起源提供了最直接的证据[31-33],为解决冠轮动物起源和演化的难题提供了来自古生物学角度的数据支撑[16,34-35]。因此,将现代的分子生物学理论与化石数据相结合,将无疑有助于最终解决这一困扰许久的科学难题。

注:最新的后生动物系统框架(修改自文献[36]),注意腕足动物门为唯一包含多种生物矿化形式的动物门类(红色虚线代表硅质,绿色代表磷酸钙,蓝色代表碳酸钙,黄色代表胶结质壳,黑色代表有机质或几丁质生物构成)。
近二十年来,笔者课题组一直聚焦寒武纪大爆发期间腕足动物的起源及其与其他冠轮动物的谱系演化研究[8,19,35,37-58],在寒武纪早期腕足动物形态解剖和生态研究方面取得了重要进展[59-61]。有研究发现,腕足动物是寒武纪大爆发和寒武纪演化动物群的主要组成部分[62-64],尤其在早古生代海洋底栖群落中发挥着主导作用,但研究表明显生宙海洋生物多样性与生态事件往往并不存在耦合关系。海洋生态系统的建立与海洋基质的演化紧密相关,尤其需要大量的生态工程建设者(ecosystem engineers)[65-67],包括大量的带壳动物(如三叶虫、软舌螺和腕足动物),也包含丰富的底栖固着生物,如海绵动物等[68-69]。带壳生物通过大量聚集的壳体富集层或分散的碎片作用于海洋底质,促进了海洋基质的演化。海洋底栖生物的相互作用同时也是海洋生态结构复杂度的反映和直接证据[70]。一定程度上,生态结构的建立和复杂性直接取决于关键物种的出现、繁盛和消失,而并非与群落生物多样性的增加和下降直接相关[71]。在寒武纪底栖固着的生物中,腕足动物非常关键,这不仅由于它们在寒武系化石出现早、丰度大[72],而且因为它们大量出现的壳体富集层对寒武纪早期海底硬化发挥着重要的作用,同时还充当底质生物(basibionts),为其他生物的发展提供关键的重要支持[48]。在生态分层方面(tiering complexity),腕足动物更是率先垂范,成为后生动物中最早建立生态分层的海洋生物,不仅出现大量的初级分层类型,还分化出了次级分层和中高次级生态分层的生态类型[43,45,48,50,59-60,73],极大地丰富了我们关于寒武纪大爆发期间后生动物的垂直群落结构的认识[8]。
特异保存的软躯体化石(布尔吉斯页岩型化石)已经成为我国寒武纪腕足动物研究的特色和优势。20世纪九十年代,由于特异型化石库的发现和系统研究,国际学者提出了腕足动物的蛤氏虫起源假说(Halkieria-like ancestor),但该假说并未得到特异型保存的腕足动物化石的直接支持[74-76]。随着澄江化石库的长期开采和化石数量的不断积累,澄江动物群腕足动物发现了特异保存的壳体内部解剖信息(soft-tissue preservation)[54-58]。国际学者[77]述评认为相关研究全面开启了寒武纪腕足动物化石生物学研究的新领域,并逐渐形成了我国寒武纪腕足动物研究的特色和优势研究领域,引领了国际相关领域的发展[59-60]。随着澄江动物群腕足动物软躯体化石研究的深入,特异型保存的腕足动物软躯体信息也引起了国际学者的极大兴趣[78],入选了国际古生物学教材、百科全书和腕足动物典籍等。以Lars Holmer为代表的国际腕足动物学者也相继加入了相关研究[77,79],并以布尔吉斯页岩化石库的化石为材料,与笔者团队全面合作,将寒武纪特异型保存的腕足动物研究推向高潮,成为国际相关领域多学科关注的热点[8,19,35,37-61,80-82]。此外,更重要的是我国华南已经成为国际上发现腕足动物壳体内部组织(如纤毛环取食系统、脉管循环系统、消化系统)的唯一地区[43,50],对理解武纪时期腕足动物的分类学、解剖学、生理学及生态学特征提供了重要信息(见图2)。截至目前,这些器官组织在国际其他地区的化石库包括布尔吉斯页岩化石库还鲜有报道[42]。
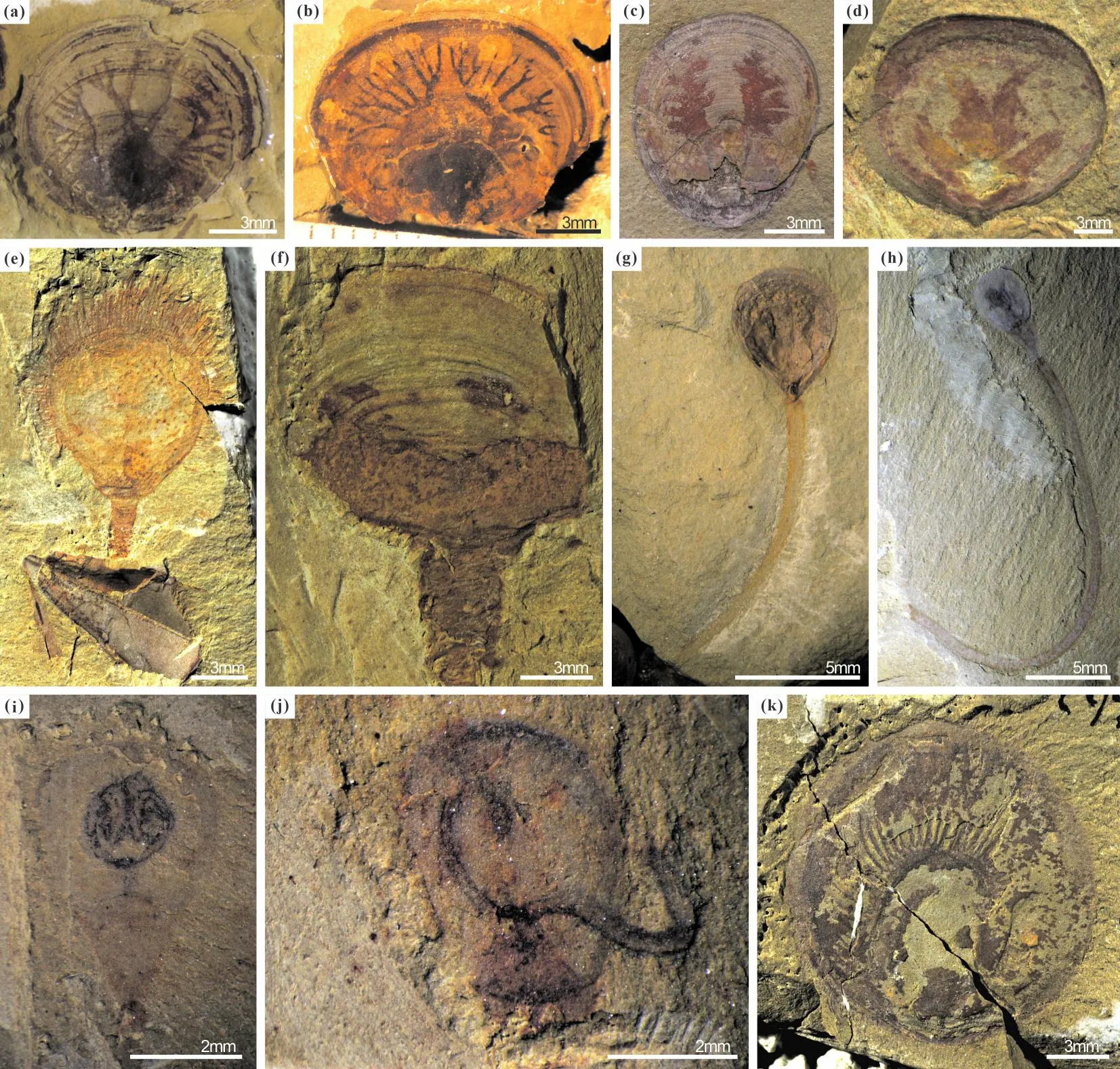
注:(a)~(d)澄江动物群和关山动物群腕足动物脉管系统,(a)、(b)纯真滇东贝,ELI-077和ELI-D-103A;(c)乌龙箐新圆货贝,ELI-GB-N-0008;(d)古柏石将军贝,ELI-SJJ-001A。(e)~(h) 澄江化石库和关山动物群发现的腕足动物肉茎,(e)海口西山贝,ELI-EJ 045A;(f)澄江顾脱贝,ELI-EJ 130;(g)马龙始圆货贝,ELI-WLP-BE0012;(h)玉案山舌孔贝,ELI-L-0085B。(i)~(k)澄江化石库腕足动物纤毛环,(i)、(j)玉案山舌孔贝,ELI-L-0050和ELI-L-0500;(k)东方日射水母贝,ELI-0662B。
寒武纪腕足动物研究长期以贝体形态和多样性为主。如上所述,腕足动物是地质历史上持续时间最长并繁盛至今的几个为数不多的后生动物门类之一,在寒武纪演化动物群和古生代演化动物群中均扮演着十分重要的角色[83],也是唯一由古生物学研究而非现代生物学研究主导的动物门类[21,84-85],其化石丰富、分布广泛,在古生态学、古地理学和生物宏演化(macro-evolutionary trends)研究方面发挥着其他门类无以伦比的重要作用[86]。同时,寒武纪腕足动物壳体类型已经出现有磷酸钙质壳(舌形贝型)和少量的碳酸钙质壳(小嘴贝型)的腕足动物[87]。由于早期学者认为寒武纪腕足动物在生物地层划分和对比方面的意义较小[88],因此其分类和多样性研究在国内外相关领域长期停滞不前。幸运的是,国际上磷酸钙质壳腕足动物的研究由于酸蚀处理方法的应用发展十分迅速,发表了大量的学术论文[89-90],极大地促进了人们对寒武纪生物辐射模式的理解[91]。然而,关于腕足动物起源的假说却主要停留在以现生动物为代表的研究材料上[92-93],长期存在单系起源、并系起源和多系起源之争[96],加之早期寒武纪腕足动物的形态和分类研究主要以贝体外形、铰合面的长度、宽度和肉茎孔的有无等硬体骨骼为鉴定特征[95],尤其在无铰类的分类谱系研究方面争议较多[96]。此外,腕足动物起源及内部各大分支的谱系关系通常仅以帚虫动物为外群[21,85,97-98],而分子生物学研究认为帚虫动物代表一类无壳的腕足动物[99-101]。因此,早期的腕足动物系统分析与外群选择存有很大异议,未能考虑腕足动物门之外的其他冠轮动物类群如软体动物、环节动物等其他动物化石记录和现代分子生物学研究的最新成果[100],使其演化极向长期很难确定并存有较大争议[102]。由此,根据多年的腕足动物壳体内部软躯体信息的研究,笔者在腕足动物起源方面提出了胶结质椎管状腕足动物起源新假说[35,37],得到了国际学者的普遍关注和肯定。基于笔者研究进展,不同的国际学者先后提出了U型肠道理论和管状世界等假说[103-104]。最近,该椎管状躯体构型模式和演化假说再次被国际学者在2017年的Nature论文中扩大,并包括了更多的寒武纪相关生物化石类群[105]。系列研究极大地促进了寒武纪冠动物分支(包括腕足动物、帚虫动物、软体动物、环节动物和软舌螺动物)的分异及其谱系演化研究。然而,国际上腕足动物的起源仍扑朔迷离,至少存在4种假说:帚虫状祖先起源、蛤氏虫起源、托莫特壳起源,以及笔者提出的胶结椎管状动物起源假说[37]。本文根据最新的化石发现及相关研究进展,对这些假说进行了广泛讨论,那么这些假说孰是孰非,相互关系如何,如何证实和证伪?
1 腕足动物起源与冠轮动物谱系演化
腕足动物因其独特的形态学特征和壳体矿化特点(兼具磷酸钙质和碳酸钙质),在后生动物演化和系统学研究中一直成为分类系统学和演化谱系研究的焦点和关键材料。目前有关腕足动物起源和早期演化的假说仍众说纷纭,争议较大[35,74,78,99,104,106-108]。西北大学腕足动物课题组前期的总结认为[37],腕足动物的起源与演化必须坚持以腕足动物的典型特征演化为基础,主要讨论和相关假说集中在以下3大问题:① 腕足动物和帚虫的系统学及演化关系;② 腕足动物和寒武纪托莫特壳(tommotiids)之间的关联及演化关系;③ 腕足动物和其他寒武纪冠轮类化石(如Wiwaxia和Halkieria)的谱系演化及关系3个方面。澄江化石库(寒武系第三阶)发现的化石新种——精美玉玕囊形贝(Yuganothecaelegans)因具有砂质胶结的椎管状身体、腕足动物状纤毛环取食器官和边缘刚毛、U形的消化系统和蠕虫状的肉茎附着结构,以及成对壳体内发现的纤毛环等结构,成为联系腕足动物门、帚虫动物门和椎管状托莫特壳的独特化石类群[35,37]。在此基础之上,随着更多类型化石的不断发现和技术手段的进一步提升,近年来的一系列进展为解决腕足动物的起源与冠轮动物谱系演化提供了更多数据支撑[21-23]。在此,笔者将对腕足动物起源的这些假说进行回顾,并根据最新的化石发现对这些假说进行梳理与讨论。
1.1 蛤氏虫式腕足动物起源假说
现代分子系统学为腕足动物和其他动物门类的相互关系提供了有力的分类框架[29,109], 但分子动物谱系研究结果和假说难以解释腕足动物祖先的形态、 出现时间和演化过程等问题。 为了解决此问题, 腕足动物分子系统学专家Cohen博士将自己的研究成果与发育生物学和古生物学结合, 在早期学者的研究成果基础上[85,108,110-111], 提出了“腕足动物躯体折叠假说”(Brachiopoda fold hypothesis, BF)[107](见图3), 简称腕足动物折叠假说。 该假说认为腕足动物与帚虫的发育躯体构型相同, 建议将腕足动物的背腹壳修订为前后壳, 并结合早期的化石研究[112], 认为腕足动物起源于寒武纪化石蛤氏虫(Halkieria)样的祖先[108](见图3)。
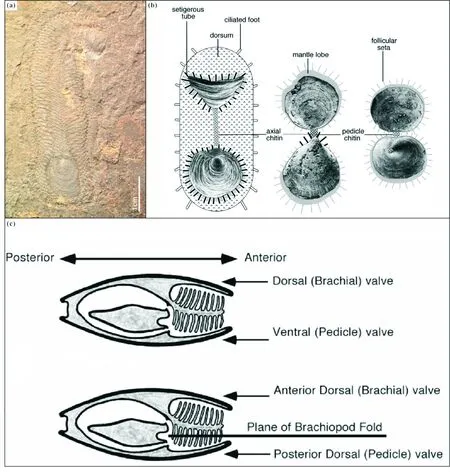
注:(a)Halkieria;(b)Micrina;(c)腕足动物折叠假说。
腕足动物躯体折叠起源假说或蛤氏虫假说受到了腕足动物学家的热烈欢迎,并长期作为腕足动物起源的演化模型[84,108,113-114]。随后,许多古生物学者试图通过壳体显微结构的研究解决缺失的中间环节[108,114]。一个很重要的潜在过渡类群即寒武纪的托莫特壳类化石(Tommotiids),其中的Tannuolina和Micrina壳体表面具有洞穿壳体的管状结构,且管状结构内壁保存有纵向排列的线状延伸,被解释为刚毛,据此Williams和Holmer[114]推测Micrina骨片与Halkieria的壳体同源,并提出蛤氏虫(Halkieria)状的生物如何演化成腕足动物的地质演化过程,在这个过程中涉及的重要过渡类型包括:Halkieria-Micrina-Mickwitzia-Prototreta[108](见图3)。与其他学说一样,该假说的提出受到了不同学者的质疑,Li和Xiao[115]认为Tannuolina的骨片由左右旋排列的帽状和叠瓦状的鞍形骨片构成,并据此推断Micrina的骨片与Halkieria的壳体纹饰的相似性并非同源,而可能为一种趋同现象,但Williams和Carlson[114]仍坚持Micrina和Halkieria壳体的相似性及其同源性。遗憾的是格陵兰寒武纪早期地层发现的蛤氏虫(Halkieria)最终被学术界普遍认为是一种软体动物[104,116-117],这项研究对古生物学家提出的腕足动物蛤氏虫模式来说无异于釜底抽薪。更重要的是,对托莫特壳类Micrina形态和生态的进一步研究发现早期根据蛤氏虫模型恢复的形态存在严重的错误[74,118],应修订为背腹铰合的表栖固着生物类型。与此同时,现生腕足动物髑髅贝的发育再研究取得了新的进展[115],新的证据表明髑髅贝Neocraniaanomala在发育过程中并未发生Nielsen[111]描述的折叠和反转(fold and inversion),新髑髅贝(Neocrania)幼虫在开始固着后,背外套膜分泌形成背壳,但此时腹壳并没有发生,延迟发育。因此,腕足动物躯体折叠起源假说也就是蛤氏虫假说在发育生物学和古生物学研究不断深入的条件下最终宣告失败。
除了蛤氏虫外,威瓦西虫(Wiwaxia)的相关研究表明其对冠轮动物的演化具有重要意义[42]。威瓦西虫是一类出现在寒武纪早期的奇特冠轮类生物,因为其背部覆盖叶片状的骨片、外形诡异,一直被认为是寒武纪大爆发的明星动物代表(见图4)并作为蛤氏虫类的关键类型。其化石最早发现于加拿大的著名布尔吉斯页岩化石库,近两年在我国华南多地发现[42]。但因缺乏背部骨片与身体组织如何相互连接的关键化石证据,其生物属性一直存有争议。笔者在澄江化石库发现了5.2亿年前的威瓦西虫化石标本,在形态上看似处于未成年幼虫阶段,因其骨片簇状聚集呈蝴蝶形,从而被命名为凤蝶威瓦西虫(Wiwaxiapapilio)[42]。化石分析表明凤蝶威瓦西虫身体背部的骨片主要可分为9排、2列,每排呈蝴蝶形簇状对称排列在身体两侧,其骨片显示了清楚的分节性特征以及明显的同律性的特点(即环节动物门的关键特征)。这种分节排列的骨片与早期恢复的外缘型的骨片排列明显不同(见图4)。此外,这些幼体标本同时还特异保存了软体动物齿舌状的口器和腹面的外套膜和铰合组织〔见图4(a)、(b)〕。综合系统分析认为5.2亿年前的威瓦西虫属于软体动物门和环节动物门的基干类群,支持了软体动物门和环节动物门的单系起源假说,但谱系演化上与腕足动物系统演化关系较远。

注:(a)、(b)ELI-W001;(c)ELI-W004;(d)ELI-W003;(e)ELI-W005;(f)威瓦西虫复原图。
1.2 帚虫状腕足动物起源假说
基于腕足动物壳体成分及形态研究,现生的三个亚门(髑髅贝型亚门,舌形贝型亚门,小嘴贝型亚门)都是由外套膜分泌形成双壳(bivalved shell),发育独立的过滤摄食系统。三个亚门的主要区别在于成体消化道的形态和排列不同:舌形贝型亚门的消化道为U形,口和肛门均开口于外套腔;髑髅贝型亚门消化道沿体轴向后延伸,两端洞开;小嘴贝型亚门消化道呈近C形,有口无肛门,口开于外套腔。再者,腕足动物肉茎的发育也有差异,舌形贝型亚门的肉茎中空,是体腔向后的延伸,这种肉茎与现生小嘴贝型亚门(碳酸钙质壳类或铰合类)表现出的纤维化实质充填且无体腔的肉茎明显不同。而现生髑髅贝型亚门腕足动物与前两者都不同,不发育肉茎,消化道直线型,开口向后。在生物与化石的形态解剖发育和壳体形态特征的基础上,国内外研究专家普遍支持腕足动物的单系起源[85,97-98,120-121],并认为与帚虫动物、外肛动物为姊妹类群,称为触手冠动物门(lophophorate group)[27,122]。
那么,腕足动物的祖先形态如何,是否与现生腕足动物相似?Williams和Hurst[123]最早提出假说,认为腕足动物的原始类型(proto-typic)可能是一种弱发育或可萎缩的具三节体腔的原口动物,如现存的有铰类腕足动物幼体,发育有后肾管和一边弯曲的直肠构成消化系统,肛门在触手冠之外,由一排环形的丝状触手环绕口部组成摄食系统。基于壳体内部解剖特征的相似性[85,92,93,124-126],早期传统的形态发育研究认为腕足动物的起源与帚虫动物紧密相关。之后,有学者提出腕足动物可能起源于身体三分的、管栖的、有体腔的帚虫状祖先[94,127],因此两者被认为是姊妹类群[111]。
在后生动物的系统框架内,腕足动物门和帚虫动物门之间的系统亲缘关系得到了大量分子系统学和基因组系统学研究的有力支持,但这两个门之间的相互演化关系问题仍未能得到解决,分子演化系统学研究反而增加了更多的争议,提出了多种不同的分支假说[128]。Helmkampf等人[27,29]将来自于39个后生动物类群中包含的11445个氨基酸位置和79个核糖体蛋白通过贝叶斯分析证实了腕足动物门和帚虫动物门的单系起源,并建议两个门应合并组成一个新的分支或新门——帚腕动物(Brachiozoa)。但是Cohen[100]对腕足动物进行了分子生物学研究,认为帚虫动物应属于一类无壳的腕足动物,建议将传统的帚虫动物门降级为帚虫动物亚门或纲,然后置于扩大的腕足动物门中[99-100]。同时,他认为非铰合类腕足动物(包括舌形贝型亚门和髑髅贝型亚门腕足动物)与钙质壳有铰类腕足动物关系较远,而与帚虫动物门谱系演化关系更为紧密,因此建议舌形贝型腕足动物与帚虫动物形成单系群,缺乏引起了化石研究的极大关注[101]。若该假说正确的话,那么帚虫动物则无疑隶属于腕足动物门内群。但是,U形消化道在寒武纪舌形贝类腕足动物得到了很好的印证[53,55-56],其扁平的贝壳也很难直接与现生的帚虫动物形态相类比,加拿大发现的所谓“软壳”腕足动物实质上是泥页岩扁平保存的舌孔贝类壳体印痕[101],因此很难想象成现生帚虫动物的舌形贝型祖先。
但事实上,前期的研究与分析都存在一定的局限性,大部分研究都是基于腕足动物和帚虫动物分支的内群特征,缺乏帚虫-腕足动物分支之外的动物类型作为分析外群,致使演化极向的确定缺乏分析依据。同时,对于腕足动物起源于帚虫动物的假说对腕足动物祖先出现的时间、壳体起源和演化趋势等问题未能给出任何解释和答案。特别是Nielsen[111]通过研究发现髑髅贝腕足动物的胚胎发育过程与帚虫的体制构型截然不同:帚虫发育过程中躯体向背面弯曲形成U形,而腕足动物的U形消化道则是向腹面弯曲形成,因此推测腕足动物和帚虫之间不可能存在任何相互演化的关系,但不可否认的是它们可能具有共同的蠕虫状的祖先[129]。该推测也得到了部分分子生物系统学研究的证实[28,116],相关研究认为腕足动物和帚虫动物共同组成一个分支,并与纽形动物(Nemertea)一起形成一个更大的分支类群,并将其命名为隐担轮动物(Kryptrochozoa)[28],然而该谱系分类目前无法在化石上求证。
考虑到对腕足动物与帚虫动物演化关系的争议,笔者将该研究追溯至寒武纪澄江生物群,其中保存有早期被认为可能是帚虫动物的化石标本——澄江拟管虫(Iotubachengjiangensis)。澄江拟管虫于1997年见于我国云南澄江化石库[130],该动物身体前部发育有成圈的刺状结构因而被认为是最早的帚虫动物[131],但一直未有详细的系统描述和研究。基于西北大学早期生命研究所长期积累的15枚标本,笔者对澄江拟管虫化石进行了详细的观察(见图5)。

注:(a)腕足动物起源于帚虫状动物假说[93];(b)澄江拟管虫的古生态复原[132];(c)~(g)澄江拟管虫的化石形态及成分分析,(c)、(d)ELI-S-001;(e)、(f)ELI-S-002A;(g)ELI-S-005A。
研究发现,该动物保存成蠕虫型,形态上可分为圆柱形的躯干和可伸缩的吻部。吻的基部周围长有束状长刺,绕躯干前部周围环状分布。吻部可收缩和膨胀,膨胀时表面具有明显的瘤突,收缩后瘤突不可见。躯干表面瘤突比吻部表面的瘤刺或者钩状结构强烈,凸起较大。身体内中空,体腔内发育消化道,消化道前边形成一个漏斗形或鳞球形的前肠区(咽腔),咽部之下为肠道,肠道向下,部分膨大,向后变细,肛门位于蠕虫身体的后末端,两侧发育线状的腺体。新研究发现这些刺实质与帚虫的触手形态完全不同,呈束状和栉状分布,可以伸缩或内卷到身体前端,可伸缩的吻部上发育成对的瓣鳃,功能上猜测为取食器官。该生物化石与现代的扇毛虫类具有极大的形态相似性,通过肠道形态的对比,认为该生物也是在海底表面进行沉积取食[132]。同时,研究进一步整合了现代环节动物的形态和分子数据进行支序分析,结合贝叶斯分析、最大似然法和简约法等多种手段,所有分析都高度支持澄江拟管虫+扇毛虫分支,表明该化石代表地球已知最早的隐居型丝鳃虫中的扇毛虫(俗称笼头虫)(见图6)。因此,澄江拟管虫的研究将环节动物的辐射从分子生物学研究推测的奥陶纪前推到地球显生宙的起始——寒武纪大爆发时期,表明环节动物冠轮类原口动物在寒武纪第三阶之前已经大量出现,首次将环节动物的祖先前推至寒武纪早期;另一方面,澄江拟管虫属于高度演化的冠群环节动物,说明其他谱系类群在它们出现前应该已经发生辐射,推测环节动物的诸多类群在澄江动物时期已经高度多样化,同时表明现生浅海底栖的多数动物类型,如环节动物可能生态上并不适宜在寒武纪特异埋藏动物群环境中繁盛。
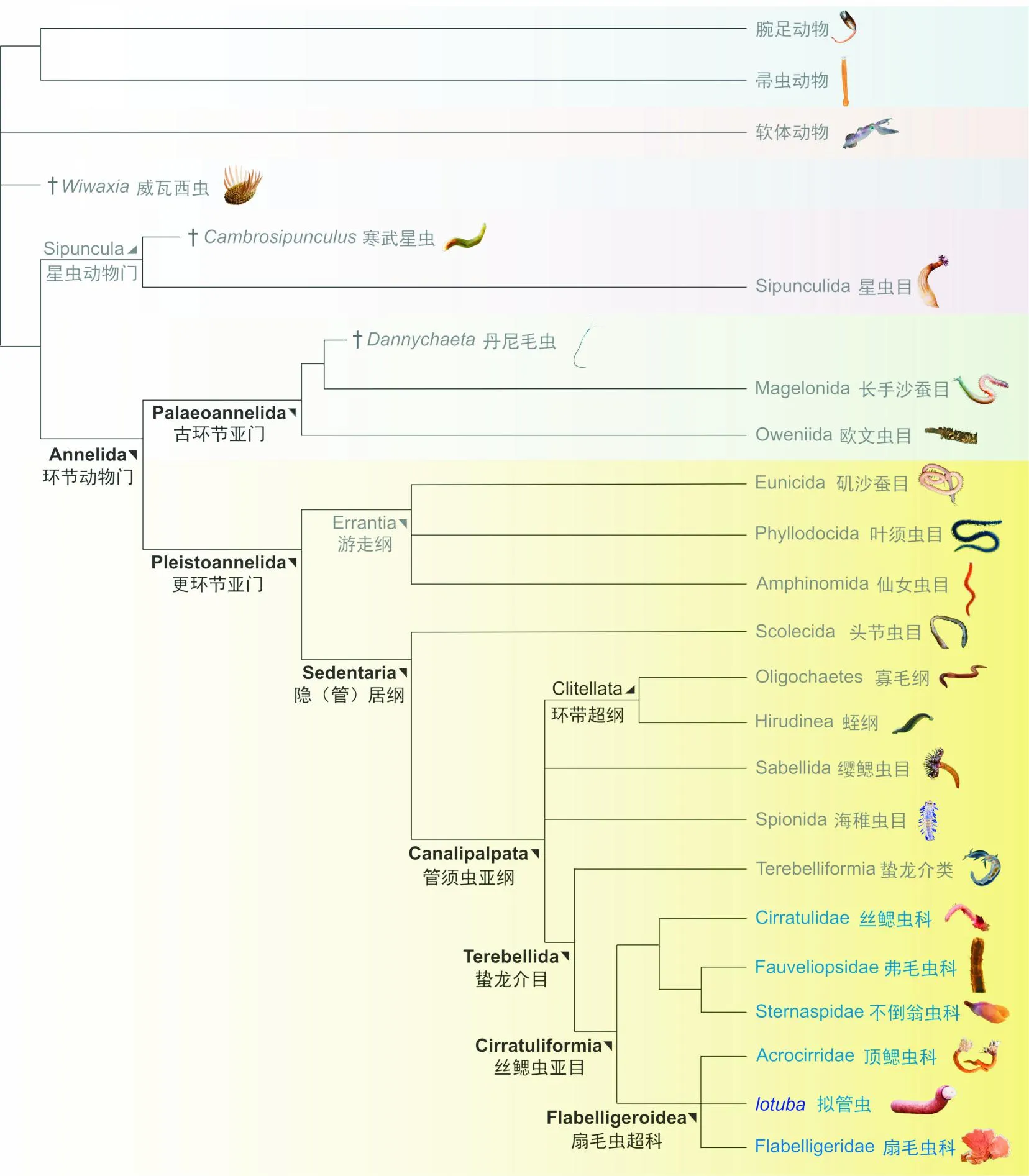
图6 澄江拟管虫的系统分类位置[132]Fig.6 Phylogenetic analyses of Iotuba chengjiangensis adapted from [132]
1.3 托莫特壳腕足动物起源假说
前面针对腕足动物起源的假说,主要从腕足动物的宏观形态上开展分析。随着近几十年寒武纪小壳化石研究的进步和深入,大量来自不同产地的寒武纪早期多门类动物的壳体化石被报道[78,106,108,114,118,132-134]。其中一些形态奇异且不完整的骨片系壳体化石——托莫特壳类,引起了学者的关注[84,108,113-114]。基于托莫特壳Tannuolina,Micrina表面穿孔,壳体外遍布管状的穿孔刚毛结构,Williams和Holmer[114]推测Micrina骨片与Halkieria的壳体同源,并指出这类托莫特壳类化石很可能是腕足动物由蛤氏虫起源到演化的潜在过渡类群[108,114]。然而,近期针对蛤氏虫Halkieria的相关研究表明其属于软体动物[104,116-117],因此表明蛤氏虫经由托莫特壳类演化为腕足动物的过程很难证实。
管状保存的托莫特壳Eccentrotheca和Paterimitra的发现重新引起了学者对托莫特壳和腕足演化关系的讨论[105,134]。Eccentrotheca和Paterimitra是呈多骨片铰合的椎管状保存的托莫特壳类,被论证是一类营表栖固着的底栖生物,与腕足动物亲缘关系较近[105,134]。研究认为腕足动物可能起源于托莫特壳类Eccentrotheca的椎管状生物[134]。Skovsted等人[134]发现托莫特壳Eccentrotheca状的管状壳体是通过一系列骨骼融合简化而成,并可能进而演化成两瓣壳封闭的腕足动物纤毛环滤食系统。随之,该化石的发现与生态恢复同时也深刻影响并改变了早期对于托莫特壳Micrina的解译,并使Holmer[74]很快修订了早期恢复的Micrina形态,彻底放弃了前后壳体排列的鼻涕虫状的蛤氏虫形态,认为Micrina为底栖固着的半封闭的铰合的腕足动物祖先过渡类型,代表着托莫特壳从完全开放的纤毛环滤食系统演化至双壳封闭的纤毛环滤食系统的过渡。
Eccentrotheca状椎管的形态〔见图7(d)〕及其表面的瘤状装饰引起了笔者课题组的注意(见图7),并发现这些管状的壳体与澄江化石库发现的瘤状杯形虫在杯体形态上具有极大的相似相。同时,杯体上都具有相似的瘤状凸起和环状纹饰〔见图7(d)、(e)〕。因此,笔者在欧洲交流学习期间,联合瑞典学者对澄江化石库的瘤状杯形虫(Cotylediontylodes)与托莫特壳开展了对比研究[44]。研究发现我国澄江化石库中发现的瘤状杯形虫的研究不仅对理解Eccentrotheca状的托莫特壳类的生物属性提供重要软躯体化石信息,而且对理解腕足动物的起源和托莫特壳腕足动物起源假说有重要意义。
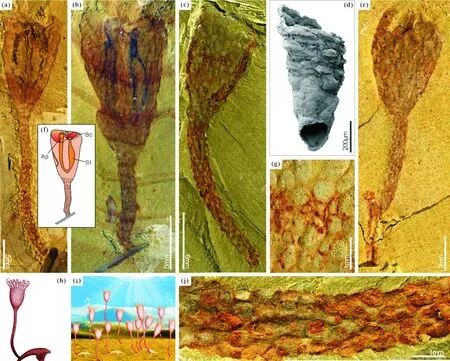
注:(a)~(c)瘤状杯形虫,ELI-C-0369,ELI-C-0359A,EJ 232A;(d)托莫特壳Eccentrotheca;(e)~(j)瘤状杯形虫,(e)ELI-EJ 224A;(g)、(j)为(c)的局部放大可见圆形或椭圆形骨片;(f)、(h)、(i)为瘤状杯形虫个体和群体的解译图与复原图。
研究发现瘤状杯形虫身体三分,上部为杯状的萼部,萼部长有杆状的柄,柄的末端发育可以收缩的吸盘(attachment disc)(见图7)[44]。化石显微观察发现瘤状杯形虫萼状的杯体表面布满圆形或椭圆形骨片,大小介于0.2~2.8 mm,这种杯体形态及其表面的骨片发育形态与Skovsted等人所发现的多骨片铰合、椎管状保存的托莫特壳类极为相似,两者同为底栖固着型生活。澄江化石库保存的瘤状杯形虫为碳酸盐岩酸蚀处理发现的Eccentrotheca状椎管提供了更多的软躯体化石信息,使理解Eccentrotheca状生物的生物属性成为可能。与仅保存了骨骼的托莫特壳类不同,澄江化石库中的瘤状杯形虫保存了极好的内部软组织结构。化石研究发现瘤状杯形虫杯体内包含U形的消化系统〔(见图7(a)、(b)、(f)〕,杯体顶部发育有成圈的柔韧触手〔(见图7(e)、(f)、(h)〕,消化道两端洞开,均包括在萼部顶部的环状触手内。环状的触手环绕着口和肛门是现代内肛动物的典型特征,该化石发现表明澄江动物群瘤状杯形虫与现生的内肛动物亲缘关系较近,代表着地球已知最早的内肛动物[44]。该研究引起了国内外学者的极大关注,英国BBC Nature和ScienceShot以“寒武纪奇异的动物伸出触手为题”做了专题评论。然而,大家都知道现生的内肛动物躯体狭小,属于假体腔动物。与之不同的是,澄江化石库的瘤状杯形虫个体较大,杯体内具有明显的泥质充填的空腔,固着柄内保存有明显的黑线,内部中空,属于典型的真体腔动物。因此,现生内肛动物的假体腔可能是在罅隙群居生活中通过小型化作用(miniaturization)演化而来的[44,135]。之后,该假体腔动物小型化起源假说得到了国际学者的注意和支持并发表在CurrentBiology杂志上[136]。
1.4 胶结质椎管状腕足动物起源假说
澄江化石库腕足动物的研究为寒武纪腕足动物的内部解剖和性状特征提供了直接化石证据。其中的腕足动物不仅保存了壳体外缘的刚毛和肉茎[64,75],而且还保存了壳体内的纤毛环、U型消化道和外套膜血管组织(外套管)[55-58]。系列化石研究发现,寒武纪腕足动物具有发达的肉茎固着系统[51-55]、不同类型的消化道包括U型和直线型[51-53],壳体内具有不同大小的纤毛环滤食系统[55-57],外套膜内能发育多级分叉的血管组织。所有特征表明寒武纪腕足动物的壳体形态和性状特征已经高度发育并具有明显的多样性[43,50],因此,发育的肉茎支持系统、外套膜管状循环系统、U型消化系统、螺旋形的取食系统以及外套膜发育的刚毛都是寒武纪腕足动物的原始特征,也是寒武纪腕足动物系统分析的共同特征,形成了寒武纪腕足动物的五大形态组织鉴定依据。
在澄江动物群研究中,一种砂质包裹的棕黄色碎屑状化石因缺乏明确的化石外形而经常被认为非化石或者假化石,而未受到重视。但笔者在海口地区尖山剖面的化石收集过程中,在一定的层位上因其化石数量极其丰富,因此引起了笔者的注意。观察发现,这类化石具有明显舌形贝肉茎状的柄状结构,肉茎上部通常是无固定形态的砂质碎片。经过大量的化石收集发现这些化石身体四分,上部为砂质胶结的成对瓣壳(valves),壳体后端为环状的中领(collar),中领下长有相对坚硬的椎管(conical tube),椎管下端为蠕虫状的肉茎(pedicle)(见图8)。肉茎的末端经常可以发现具有鳞球状的末端结构。解剖砂质胶结的壳体发现,成对的瓣壳内表面饰有外套膜管状分支的血管系统(脉管),壳体内包裹成对的纤毛环结构取食器官(触手冠),壳体边缘长有刘海状的刚毛,中领和椎管内具有U形的肠道,椎管末端具有蠕虫状的肉茎固着结构(见图8),所有这些发现表明玉玕囊形贝具有寒武纪腕足动物的五大典型特征。然而,壳体岩石切片和成分分析表明玉玕囊形贝的壳体缺乏矿化结构,两瓣壳不属于现代腕足动物的磷酸钙质壳也不属于碳酸钙质壳,瓣壳表面覆盖有硅质碎屑胶结的碎屑颗粒,这种瓣壳属性与现代的帚虫动物栖管十分相似。经SEM、BSEM、化石切片、能谱和TNT聚类分析,认为精美玉玕囊形贝代表一类与帚虫动物紧密相关,兼具有不同腕足动物亚门和寒武纪托莫特壳(tommotiids)的触手冠动物。化石分类研究表明这些化石代表一类新的化石属种,并命名为精美玉玕囊形贝(Yuganothecaelegans),以纪念腕足动物专家金玉玕院士为推动澄江化石库腕足动物研究做出的开创性贡献[64,75-76]。玉玕囊形贝虽具有寒武纪腕足动物的五大典型形态解剖特征,但其壳体外形与寒武纪舌形贝类和小嘴贝型的壳体形态明显不同,更重要的是其贝体砂质胶结、缺乏明显的矿化特征,贝体末端长有椎管(见图8)。因此,澄江化石库玉玕囊形贝兼具有腕足动物、帚虫动物和托莫特壳的共同特征,显示了它们之间祖先的原始特征[35]。化石系统研究表明腕足动物可能起源于一类底栖固着生活的、躯体砂质胶结的缺乏生物矿化的椎管状触手冠动物,从而挑战了早期的学者提出的BF假说,即Halkieria状的祖先。国际学者评论认为玉玕囊形贝代表腕足动物的“始祖鸟”化石,瑞典科学院和乌普萨拉大学以“沙包中的腕足动物”为题做了报道,该成果“以追溯腕足动物起源”为题入选最新的欧美古生物学教材[137-138],为解决腕足动物的起源和后生动物的矿化发生以及冠轮类动物(Lophotrochozoa)之间的系统关系提供了重要的化石依据。瑞典乌普萨拉大学教授、英国剑桥大学博士Graham Budd撰文对相关成果做了专门的评论[139],对我们提出的椎管状腕足动物起源假说进行了评论,并总结为U型肠道理论。

注:(a)ELI-BLW-0091;(b)ELI-JS 0141A;(c)ELI-EJ 0473A;(d)精美玉玕囊型贝的化石重建;(e)ELI-EJ 0580B;(f)ELI-EJ 0483A;(g)ELI-JS 0475A。
玉玕囊形贝的发现和胶结质椎管状腕足动物起源假说的提出得到了国内外学者的极大关注,并在国际学者的古生物前沿进展一文中原图引用[102]。该类谱系分析被国际学者进一步扩大包括寒武纪更多的椎管状化石[105]。部分学者在该假说的框架下,重新梳理了冠轮动物的系统发育树,并试图寻找类似形貌特征的化石。2017年Moysiuk等人[105]继Babcock和Robison[140]后再次报道了保存在布尔吉斯页岩中软舌螺目单臂螺Haplophrentiscarinatus,与前期的报道不同的是Moysiuk等人将软舌螺上保存的丝状触手重新解释为与腕足动物类似的触手冠(lophophore)。在胶结质椎管状腕足动物起源假说的基础上,软舌螺的椎管状形貌对比前期报道的托莫特壳类Micrina以及精美玉玕囊形贝,具有极高的相似性,加之软舌螺兼具底栖滤食的生活方式,为此Moysiuk等人进一步认为软舌螺是以腕足动物为代表的触手冠动物的一分支类群,代表一类由多骨片(multi-scleritome)愈合发育而成的触手冠动物。随后,Sun等人[141]2018年又重新修订了在澄江生物群中一类直管螺动物——偶线带螺。认为这类直管螺的末端保存类似腕足动物的肉茎结构,将其命名为云南肉茎螺,从而进一步论证了软舌螺化石属于腕足动物类触手冠动物,代表腕足动物的基干类群,后期相关研究甚至将软舌螺放置在腕足动物冠群中[141]。但是该观点并没有得到学术界的认同[143-148],反而引起了软舌螺生物亲缘性讨论的更多争议,后续关于软舌螺解剖学和壳体纤维结构学的证据彻底证伪了“软舌螺是触手冠动物”的猜想[146,147,149]。
在壳体的显微结构对比方面,Li等人[147]对寒武纪的软舌螺(包括软舌螺目与直管螺目)和软体动物的壳壁微细结构与矿物组成进行了精细研究,结果表明软舌螺的壳体与软体动物具有相似的生物矿化类型(壳体的微细结构都显示文石板片状),同源的矿化壳体特征指示软舌螺应该与软体动物具有更近的亲缘关系,而并非腕足动物。最重要的是,澄江化石库中三槽螺(Triplicatella)化石的发现进一步证实了软舌螺并非属于触手冠动物而是冠轮动物干群。三槽螺保存了精美的触手状软躯体结构,粗壮的扇形取食器官位置并不固定,化石显示这种触手可在口盖前侧腹扇面内外自由伸缩。对比该触手状结构与触手冠动物(腕足动物、帚虫、苔藓虫等)环绕口部的纤细滤食纤毛环结构(lophophore),形态上可以表明二者是两种摄食功能不同的触手结构。触手冠动物(帚虫、腕足动物等)的纤毛环是中体腔延伸的触手状摄食器官,呈环口状,纤毛环固定附着壳体上,触手上发育的成排纤毛,由纤毛触手控制通过扰动水流的方式,滤食水流中存在的食物颗粒,沿触手进入腕沟最后流经外套腔进行取食,而三槽螺的粗壮可伸缩触手据推测为可以在壳体内自由伸缩进行沉积摄食(见图9,10)。同时,Liu等人[146]对千余块澄江化石库中的软舌螺的椎管始端结构进行了详细观察与分析,所谓的“肉茎”结构被证实为椎壳末端残留的部分碎片或印痕而非软躯体组织。因此,软舌螺并非属于触手冠动物类群以及托莫特壳类化石与腕足动物演化过程中的某种椎管状干群动物,而其在系统发生树上的位置应该位于冠轮动物的基部。后续软舌螺相关的形态结构演化更是证实了软舌螺并非触手冠动物的观点[144-145,149,152-153]。软舌螺起源于直管螺目,经近小角螺科过渡,逐步演化出具附肢结构的典型软舌螺目[150],生态上逐步实现由食泥、食腐转变为被动滤食的摄食方式[152-153]。

注:(a)、(b)澄江生物群中Triplicatella opimus[146,150],ELI H-113,ELI H-129A;(c)、(d)关山生物群中‘Linevitus’ malongensis[151],WD-SJJ-584,WD-SJJ064A;(e)、(f)显微CT识别出石牌生物群中Doliutheca orientalis保存板片状的锁骨结构,ELI QJP-SP-H-CT001;(g)~(q)石牌生物群中Doliutheca orientalis及其次生保存的线状壳顶末端,(g)ELI-QJP-SP-H-SSF-81A2001; (h)ELI-QJP-SP-H-SSF-81A4001;(i)ELI-QJP-SP-H-SSF-81D7001; (j)ELI-QJP-SP-H-SSF-81A16001;(k)ELI-QJP-SP-H-SSF-0781039;(l)ELI-QJP-SP-H-SSF-61016;(m)ELI-QJP-SP-H-SSF-816027;(n)ELI-QJP-SP-H-SSF-816045;(o)ELI-QJP-SP-H-SSF-8110124;(p)ELI-QJP-SP-H-SSF-811067;(q)ELI-QJP-SP-H-SSF-61021。
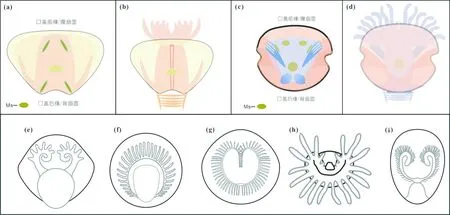
注:(a)、(b)直管螺丰满三槽螺触手[146,150];(c)、(d)关山生物群中软舌螺马龙‘线带螺’触手[151];(e)寒武纪澄江生物群中玉玕囊型贝触手;(f)寒武纪澄江生物群中东方日射水母贝触手;(g)现生腕足动物Pumilus antiquatus触手;(h)现生帚虫动物触手Phoronis ovalis;(i)寒武纪澄江生物群中玉案山舌孔贝。
除了软舌螺这种椎管状化石引起了广泛关注外,以胶结质椎管状腕足动物起源假说为依据,笔者及团队还对另一类出现在寒武纪纽芬兰世的发育固着器的更早椎管状化石进行了深入的研究与探索,试图探索椎管状腕足动物的深时更多证据。在湖北宜昌地区寒武系底部岩家河组灰岩夹层的泥质粉砂岩中发现和收集有大量的椎管状宏体化石——双腕小岩家河虫(Yanjiahellabiscarpa)[154-155](见图11)。该化石身体也可分3部分,上部具有圆形或者椎形的球囊,表面覆盖有钙质的骨板,前端长有一对触手状结构,球囊后端长有具环纹肉茎状的固着柄[154-155]。通过分支系统学和谱系研究发现小岩家河虫与触手冠动物并无亲缘关系,应该属于棘皮动物祖先。小岩家河虫身体两侧对称,兼具有棘皮动物骨板状结构和半索动物的分节特征,可能通过萼部顶端的成对触手营滤食生活。该研究对理解地球后口动物亚界棘皮动物门和半索动物门的起源提供了化石关键证据,该研究将棘皮动物的化石记录前推至少1千万年,表明现生的棘皮动物门起源上根植于两侧对称动物,其五辐射对称的躯体构型为获得性状,在演化上要晚于钙质骨板,为步带动物超门的起源提供了最早的化石证据[144,154]。
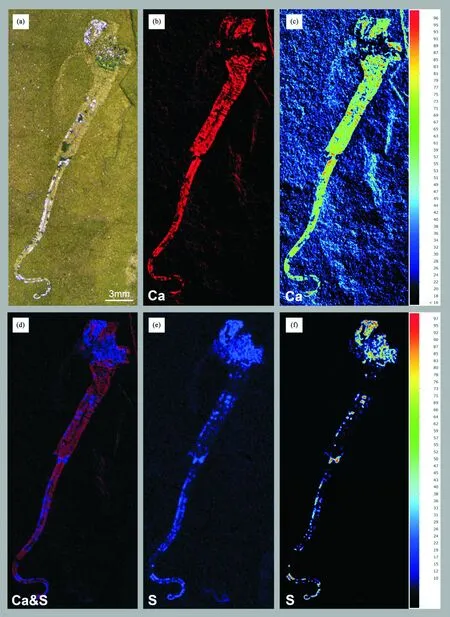
注:(a)双腕小岩家河虫,ELI-G21;(b)~(f)化石成分分析。
2 特异型化石库保存和赋存于碳酸盐岩的壳体化石
有关于腕足动物起源的相关理论化石依据多来自特异埋藏型化石库,其中以我国澄江动物群为主,化石不仅类型分异度高,软躯体组织的细节特征也较为丰富。以舌形贝型腕足动物为例,可以观察到包括肉茎、脉管、纤毛环等组织在内的多种软躯体,为验证腕足动物起源的多种假说提供了丰富的材料[8,19, 35, 37-42,43-58,156-159]。从截止目前世界范围内报道的特异化石库生物群的生物群组成类型来看,均以泛节肢动物为主,而草根型的腕足动物与其他类型的冠轮动物往往仅占据较少的比例[160-161]。近期笔者对于关山生物群的相关研究中,发现了以腕足动物为主导的海洋底栖生态系统,但从整体的化石属种多样性产出来看,其中仍以泛节肢动物占据主导[160]。这与源自碳酸盐岩或磷块岩中的壳体化石组成相比有着巨大的差异。在寒武系底部,我国梅树村剖面产出的大量壳体化石中含有大量不同类型的管状化石与软舌螺类,从数量来看以冠轮动物占主导。寒武系岩家河组与宽川铺组经过多年酸蚀处理,其中所包含的化石类型也以管状化石、软体动物与锥石类为主,明显也以冠轮动物占主导[162-163]。而近期有关于寒武纪冠轮动物的相关研究进展,如寒武纪发现最早的苔藓虫类,便来源自壳体化石富集的灯影组西蒿坪段[34],并且在同一层位发现了寒武纪最早的舌形贝类腕足动物与最早的三叶虫伴生[34](见图12)。在寒武纪大爆发极盛时期的寒武系第二统水井沱组上部的碳酸盐岩中仍以多种类型的小壳化石占主导,在其中同样以冠轮动物占主导,共发现腕足动物化石8属12种及多种未定属种,可归于2个目5个科;除腕足动物外还包括软舌螺类、软体动物等[38,40,41]。对于华北板块而言,特异埋藏化石库较少,仅有近期报道的临沂生物群[164]。然而,其中小壳化石的相关研究取得了比较大的进展。自20世纪七八十年代开始,对于华北寒武系底部碳酸盐岩赋含化石的研究,其中仅发现有Estingia(=Hsuaspis)这一种三叶虫存在,整体看来其中产出的化石以软体动物、软舌螺类、开腔骨类为主,另有部分托莫特壳、微网虫、棘皮动物骨板和海绵骨针,但仍显示出以冠轮动物占主导的特征。同时,对华北寒武系第四阶-古丈阶碳酸盐岩中壳体化石相关研究发现[165],虽然一定程度上壳体化石的出现、保存与分布明显受控于碳酸盐岩沉积微相,但腕足动物、软舌螺类在较晚时代的壳体化石中仍然占据主导地位。
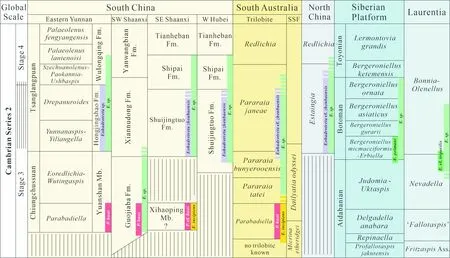
图12 华南、华北、冈瓦纳、西伯利亚和劳伦板块寒武纪第三阶至第四阶以三叶虫化石带为基础的生物地层对比,图示澄江动物群产出层位Eoredlichia-Wutingaspis三叶虫化石带,以及最早的舌形贝类腕足动物与最早的三叶虫伴生和苔藓虫产出层位Parabadiella三叶虫化石带[166]Fig.12 Tentative correlation chart for Cambrian stages 3-4 and trilobite zones of South China, East Gondwana, North China, Siberia and Laurentia, indicating stratigraphic ranges of Parabadiella huoi (marked in red) and Eoobolus incipiens (marked in yellow), and the first occurrences of other lingulide Eoobolus (marked in green) and acrotretide Eohadrotreta (marked in purple) in different palaeocontinents[166]
通过以上对比不难发现,特异埋藏型化石库与普通碳酸盐岩中(非奥斯坦型)产出的生物群从门类构成上存在着显著的差异。诚然,这可能与获取化石过程中使用的实验方法有一定的关联,但更重要的原因可能是沉积环境之间的差异,即不同相区、不同沉积环境中的生物类型本身便存在着差异。此外,从特异埋藏化石库所代表的环境来看,无论是寒武纪早期的清江[167]、澄江[168]、Sirius Passet[169],还是寒武纪中期的凯里[168]、布尔吉斯和临沂生物群[164],均代表了较深水的沉积环境[168]。其中清江生物群所处环境与同时期其他化石库相比,水体更深,导致其生物群组成与其他较深水的生物群落构成之间也存在着较大的差异[167]。特异型化石库受埋藏差异的影响,即使是同一化石库中相同类型的生物,在不同地区的保存特征也存在着显著的差异[150,152]。而特异埋藏化石库与碳酸盐岩中壳体化石的保存特征更存在着显著区别,其中特异埋藏化石库以保存生物不同部位的软组织特征而最为著名。通过这些软组织结构,能够恢复生物的解剖学特征,为了解生物的取食、消化、循环系统的组成与形态提供了独一无二的证据。同时为生物的亲缘关系分析也提供了较为独特的信息[34]。与之相反,碳酸盐岩中的壳体化石对软躯体的保存“爱莫能助”,反而却能保存较为完好的壳体纹饰[40,41,170]。此外,在壳体结构的保存上更有独到优势。特异埋藏化石库中的生物多为碳质压膜(有时为黄铁矿化)保存[171],因个体较大,对于生物生长、发育的研究难以提供有效信息。但是,碳酸盐岩中的壳体化石,不仅能够产出微小的幼体化石,还能通过对处于不同生长发育阶段生物的壳体显微解析,恢复其生长发育过程[40,172]。另一方面,碳酸盐岩赋含的壳体化石在研究壳体超微结构上有着显著优势,反观布尔吉斯页岩型化石,仅有极少数标本能够保存壳体的超微结构[158]。
从壳体成分方面来看,目前对于最早腕足动物的相关研究仍未给出舌形贝型亚门与小嘴贝型亚门出现的先后顺序,即最早腕足动物壳体化学成分的起源目前仍未解决[21]。从腕足动物起源的时间线来看,目前已知最早的腕足动物为始现于寒武纪第二期的神父贝类[173],虽然目前普遍认为后期的神父贝类具有磷酸钙质壳体[174],然而对于其原始壳体成分组成的证据却极为缺乏,仍需进一步研究。而腕足动物的矿化过程、方式与机制,对于理解腕足动物起源这一重大科学问题,有着重要的参考价值。这也是胶结质椎管状腕足动物起源假说目前越来越受到重视的原因[35,102]。亦即早期的腕足动物很可能不能自身主动分泌矿化骨骼或只有较弱矿化能力,只能通过吸附周围的碎屑颗粒,形成胶结质壳[35]。因此,对于腕足动物起源与演化的相关研究,不能仅依靠特异埋藏型化石库,无论是在地球46亿年的演化历史上,还是在长约4 000万年的寒武纪大爆发期间,虽然保存化石极为连续,但特异埋藏化石库极为稀少,也只能为我们揭开腕足动物甚至是其他类型生物起源与演化的冰山一角[175],难以了解生物群组成的全貌,故而更多关于生物演化的信息深藏在常见的沉积记录中。因此,为了更全面理解地球动物起源,仍需对寒武纪大爆发前后碳酸盐岩中所保存的壳体化石进行细致探索,以期获取足够多有关于腕足动物和冠轮动物起源与演化的壳体信息,进而恢复多门类生物起源与演化奥秘。
矿化壳体在对腕足动物起源与演化的研究中扮演着极为关键的角色,实际上不仅是腕足动物或冠轮动物,目前各个门类的研究者均已意识到生物矿化在研究早期动物生活、发育过程中的重要作用[176],使得生物矿化研究一直处于国际热点[177-178]。广义的生物矿化包括生物诱导矿化与生物控制矿化,前者多见于细菌、真菌等以微生物细胞外的胞外聚合物(EPS)为诱导基础,形成不同类型的矿物,如方解石[179]与磷灰石[170,180]。而生物控制矿化的形成又可分为两种类型,即外骨骼和内骨骼。外骨骼从生物组织器官的角度来看,属于表皮组织,而内骨骼则属于结缔组织。两者具有完全不同的形成过程与成因模式。从已有证据来看,最早的动物骨骼出现于埃迪卡拉纪末期[181-182]。通常,环境的剧烈变化被认为是矿化骨骼出现的重要诱因[181,183]。目前广受认可的另一观点则从生物间的生态关系出发,认为生物间捕食关系引发了军备竞赛,从而导致了生物骨骼的诞生[184-185]。随着生物的演化和生态系统的复杂化,生物能进行更加复杂的生物矿化结构。从前寒武纪仅有简单矿化外骨骼的克劳德管类[186],到寒武纪初期具有较强矿化能力的软体动物[177],再到寒武纪第三期之后迅速爆发、增长的其他类型骨骼生物(如腕足动物、三叶虫类、叶足动物等),不难看出自前寒武纪末期至寒武纪,组成生物矿化骨骼的矿物类型发生了改变,即从前寒武纪克劳德管的高镁方解石壳体[182],到寒武纪早期生物骨骼,多以文石为主导,进而在寒武纪中晚期到奥陶纪生物的矿化骨骼则以低镁方解石占据主导[182],其中还有大量原生有机磷酸钙质腕足动物出现。组成生物矿化骨骼矿物颗粒的成分变化,仅能反应生物在不同环境中对不同离子的利用能力出现了改变,很可能并没有反映生物自身矿化能力的演化。事实上,经历了寒武纪大爆发的后生动物,不仅在生物的体制构型上出现了显著变化,在生物控制矿化的能力、矿化结构多样性上也有了极大的提升。除了一些矿化能力较弱的后生动物,可能通过黏附外部沉积颗粒形成外部骨骼,如Yuganothecaelegans[35],而有些生物不仅可以通过生物膜发育矿化结构,也可通过身体表面黏附外部沉积物颗粒形成不同的矿化结构,如Phoronopsisharmeri[187]。
实际上, 绝大部分外骨骼的形成均由生物膜所控制, 在动物生长的过程中, 首先建立生物膜/有机质框架,而后通过生物膜向外部分泌形成矿化骨骼所必需的阴、阳离子,使得不同种类矿物在生物膜所形成的框架进行堆积进行矿化,同时矿物的堆积范围也受到生物膜的控制(见图13)。而动物的表皮组织衍生物,在壳体表面形成的瘤凸、刺等也受到生物膜的控制。腕足动物中的乳孔贝类具有特征鲜明的表皮层、初级层和次级柱状层[189]。乳孔贝在同心方向上的外缘型生长方式,便受到生物膜的控制,即由乳孔贝类首先形成在次级柱状层及初级层中的生物膜,这些膜状结构决定了乳孔贝次级柱状层在壳体不同位置的生长高度、柱状结构的宽度与分布方式(见图14)[158],不仅形成了锥形腹壳,还在背壳形成了多种形态的中隔板(刀片状、爪状等)。相较于乳孔贝类较为复杂的柱状结构,早期环节动物所具有的加积型矿化骨骼分泌方式,虽然较为简单,但也完全受到生物膜的控制,即向外分泌不同成分的矿化物质,形成外骨骼,但随着生物膜的逐渐生长或环境条件的变化,由生物膜分泌形成矿物的晶体结构与晶体取向可能会发生变化[190],形成多级层状结构。
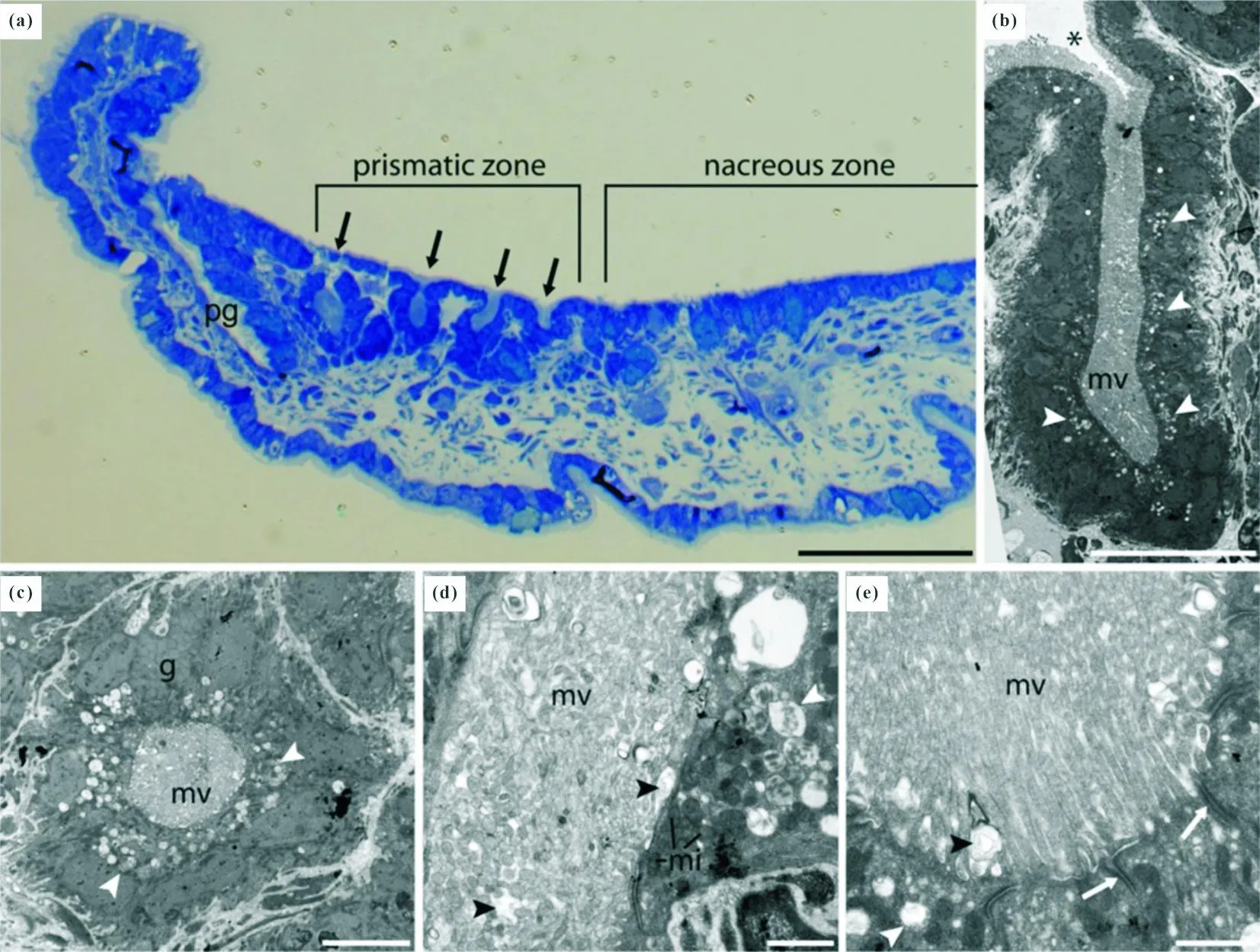
注:(a)不同壳体分泌区域的外套膜形态,珍珠层区域(nacreous zone)外套膜较平缓,棱柱层区域(prismatic zone)外套膜多呈褶状;(b)透射电镜下外套膜中腺体细胞(灰黑色)的纵切面形态,腺体细胞中充填有微绒毛(mv=microvilli);(c)透射电镜下外套膜中腺体细胞(灰黑色)的横切面形态,呈多边形不规则状;(d)、(e)微绒毛区域细节展示,白色箭头指示囊泡,黑色箭头指示分泌物。

注:(a)沿肉茎孔附近切开后的壳体形态;(b)、(c)由不同高度柱状结构导致壳层厚度出现变化;(d)黄色箭头指示单独柱体特征,壳层厚度完全受柱体控制。
从本质上,生物的多样性变化受到矿化的控制,由生物特化细胞组织作为有机质框架为生物的整体形态进行塑形,通过不同类型的生物膜调用环境中不同的阴、阳离子,分泌形成不同类型矿物,形成了不同的矿化结构。因此对于生物矿化结构的研究最近受到了持续性关注,而对于五亿多年前、经历复杂演化历史、多期成岩作用生物矿化结构研究的首要任务是对不同期次成岩作用进行区分,恢复真正的生物矿化壳体与内模结构,更重要的则是通过矿化结构恢复分泌矿化结构的“膜”状系统。这在后生动物矿化的研究中值得留意,而目前对于生物内模、内核、多期次交代化石结构的研究可能与生物膜结构难以保存有着较大关联。如果仅依靠内核、内模等非原生生物壳体对于早期动物的矿化机制进行分析,而忽略生物骨骼的属性(内骨骼或外骨骼)、生物膜结构的分布,很可能导致复原出的生物矿化模式不准确或不全面。
3 结论
腕足动物是古生代海洋底栖生物群落中最重要的冠轮类无脊椎动物,它们经历了地史中最残酷的5次生命大灭绝事件而延续至今。腕足动物身披双壳,两侧对称,是后生动物中唯一不分节且包含两类矿化壳体(包括磷酸盐质壳和碳酸钙质壳)的动物门类,发育上兼具有原口动物和后口动物的共同特点。长期以来,关于腕足动物的起源问题,存在着多种假说,包括帚虫状祖先起源、蛤氏虫假说、托莫特壳起源假说和椎管状触手动物起源假说等。本文在对以上几种腕足动物假说进行了系统回顾,并结合相关的最新化石发现和研究进展,以腕足动物起源为主线,探讨了冠轮动物的起源、辐射、演化。在此过程中,作者发现了以瘤状杯型虫为代表的地球最早的内肛动物,填补了寒武纪大爆发缺乏内肛动物门化石的空白,提出了假体腔动物小型化起源新假说。同时研究发现威瓦西虫既具有环节动物同律分节特点,又具有软体动物齿舌状口器,因此在分类上属于软体动物门和环节动物门的基干类群,支持了软体动物门和环节动物门的单系起源假说,揭示了这两个无脊椎动物门的共祖特征(Plesiomorphy)。该化石的发现和深入研究进一步否定了腕足动物哈氏虫假说,对理解后生动物门类,尤其是冠轮超门(Lophotrochozoa)中的担轮动物分支的系统演化有重要科学意义。
腕足动物在寒武纪第三阶已高度分异,出现了不同的目和属种类型。胶结质腕足动物干群可能延续了一段时期直至玉玕囊形贝的发现,该化石成为联系帚虫动物门、腕足动物门和椎管状托莫特壳的重要化石类群,因而被誉为腕足动物的“始祖鸟”化石,为解决腕足动物的起源和后生动物的矿化发生以及冠轮类动物之间的系统关系提供了重要的化石依据。但是如何理解玉玕囊形贝壳体的矿化的过程,以及对寒武纪早期椎管状化石的追溯,能否发现两端开口与玉玕囊形贝椎管相对应的壳体化石将是下一步腕足动物起源研究的思路和重点。同时结合特异型化石库中保存软躯体的化石以及碳酸盐岩中产出的三维立体小壳化石,并将宏观形态和微观结构相结合、化石成分与保存相结合,恢复壳体矿化的模式与过程,有助于对腕足动物起源及冠轮动物的演化提供更多新的见解和认识。
致谢:感谢西北大学地质学系舒德干教授的有益建议和支持,以及团队外专Lars Holmer、Glenn Brock、Timothy Topper和Luke Strotz的长期合作与交流。
猜你喜欢
小学科学(学生版)(2021年12期)2021-12-31
云南农业科技(2021年6期)2021-12-30
科学(2020年4期)2020-11-26
小学科学(学生版)(2020年7期)2020-07-28
创造(2020年12期)2020-03-17
小哥白尼(趣味科学)(2019年10期)2020-01-18
疯狂英语·初中天地(2019年4期)2019-10-17
中国漫画(2017年4期)2017-06-30
微型小说选刊(2015年27期)2015-11-17
故事会(2015年12期)2015-05-14
