
低温胁迫对辣椒果实形态和抗氧化酶系统的影响
2023-12-30 14:12邵登魁邓昌蓉王亚玲文军琴张彦君
青海农林科技 2023年4期
邵登魁,邓昌蓉,王亚玲,文军琴,3,张彦君
(1.青海大学农林科学院,青海 西宁 810016;2.青海省蔬菜遗传与生理重点实验室,青海 西宁 810016;3.青藏高原种质资源研究与利用实验室,青海 西宁 810016)
辣椒(CapsicumannuumL.)属茄科辣椒属植物[1],冬春季辣椒生产主要在日光温室中,由于青海省为高原温带、高原亚寒带和寒带气候[2],所以辣椒易受低温影响。辣椒受到低温胁迫后,会出现生理代谢紊乱、形态发育失调、生活力下降、植株发育迟缓和生殖生长失调[3]。根据报道,当植物遭遇逆境胁迫时,细胞会产生大量的活性氧和自由基,其大量积累会对细胞产生毒害,为了确保生长发育,植物体会激活主动防御系统。
本试验选择了青海省广泛种植的两个辣椒品种:华美105和乐都长辣椒为试验材料,在人工气候培养箱中对其进行低温处理,测定其生长指标和生理指标中的酶活性变化,研究低温胁迫对辣椒果实形态及生理特性的影响,旨在为辣椒抗低温冷害育种和保花保果技术研究提供参考。
1 材料与方法
1.1 试验材料
选择两种耐低温性不同的辣椒品种材料:乐都长辣椒(耐低温结实性弱)和华美105(耐低温结实性强),其中乐都长辣椒的种子由青海省农林科学院园艺所提供,华美105种子由山东省春禾种业公司提供。每个品种育苗150株,在辣椒4叶1心期时转入人工气候箱中进行不同温度处理。
1.2 试验方法
1.2.1 试验设计
本试验共设2个品种4个处理,每处理设3次重复,每个处理样本量为50株;试验条件如表1。培养箱低温处理25 d后移栽到大田,进行后续观察。试验在青海省农业科学院园艺研究所3号试验园及青海省蔬菜遗传与生理实验室中开展。
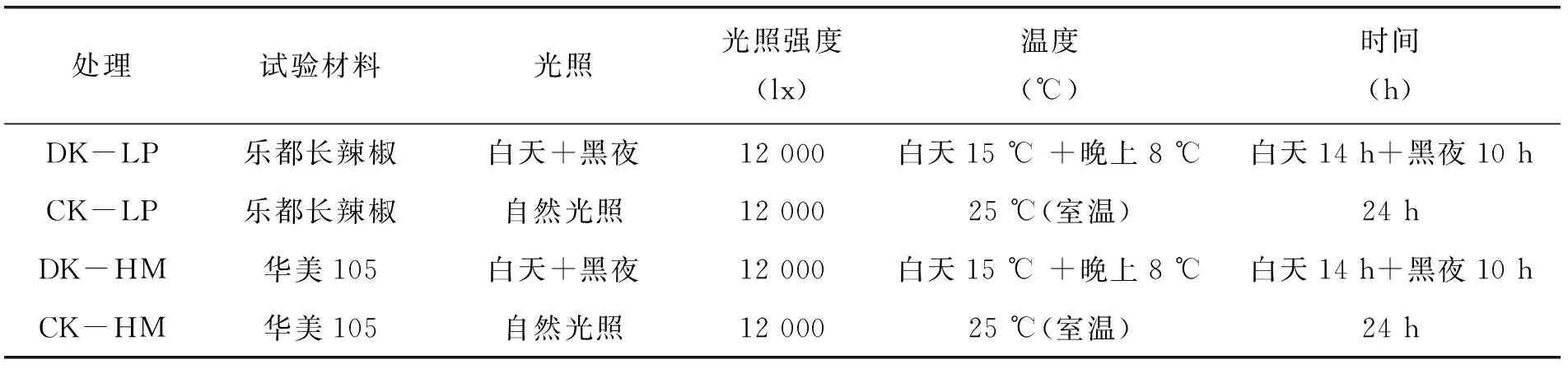
表1 试验设计
1.2.2 植物形态指标测定
株高用直尺(20 cm)测量;展叶数是对全株叶长大于2 cm的叶片数进行统计;鲜重用千分之一电子天平称重,将鲜株体置于电加热恒温鼓风炉中,在105 ℃下杀青30 min,然后在80 ℃下烘干,用电子天平称量植株干重。
1.2.3 辣椒花芽分化及花器官观察
花器官和幼果用OLYMPUS-SZX 7体视显微镜观察拍照。分别于初现蕾期、开花前3 d、开花当日、开花1日、开花3日和开花5日进行观察。
1.2.4 生理指标测定
可溶性糖测定采用蒽酮比色法[4];叶绿素含量测定采用乙醇法[5];超氧化物歧化酶(SOD)活性测定和氧化氢酶(CAT)活性测定参考陈建勋等的方法[6-7];过氧化物酶(POD)活性测定参考《植物生理实验指导》的方法;抗坏血酸过氧化物酶(APX)活性测定参考赵世杰等的方法[8]。
1.2.5 数据分析
使用Microsoft Excel 2019对试验数据进行计算;采用Origin 2019 绘图。
2 结果与分析
2.1 低温胁迫对辣椒花器官显微结构的影响
如图1A所示,乐都长辣椒部分胚珠坏死,只有授粉管伸长正常的部位胚珠发育比较正常,其余胚珠大量坏死呈褐色。结合对照分析,乐都长辣椒花朵脱落是低温环境造成辣椒授粉不完全,胚珠发育受阻,导致大部分胚珠坏死,从而影响辣椒子房发育,造成结果期落果现象的发生。乐都长辣椒在低温条件下落花落果的可能原因是低温影响花粉萌发和花粉管伸长,导致胚珠受精不良而败育,进而引起子房体发育不良或停止。从图2和图3比较可以看出,在低温胁迫下,乐都长辣椒在开花后子房体发育缓慢,花瓣脱落晚(图2e,图2f),而华美105花瓣在开花后3日就萎蔫脱落(图3e),开花后5日子房体明显长大(图3f),二者的差异可能就来源于受精或花粉管生产刺激与否。由于华美105属于耐低温结实性品种,试验当季未发生明显的落花落果现象,而乐都长辣椒不耐低温结实,发生了落花落果。
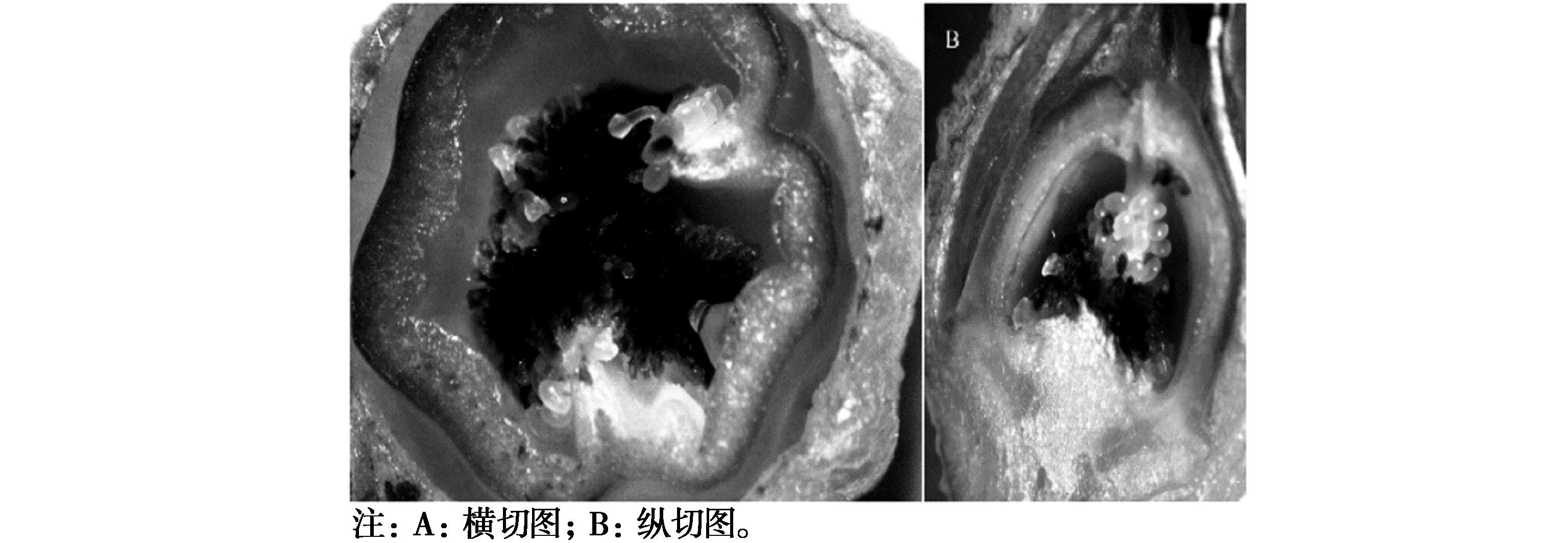
图1 低温处理下乐都长辣椒脱落花器官图Fig.1 Low temperature treatment led to the fall of the long pepper shedding flower organ map
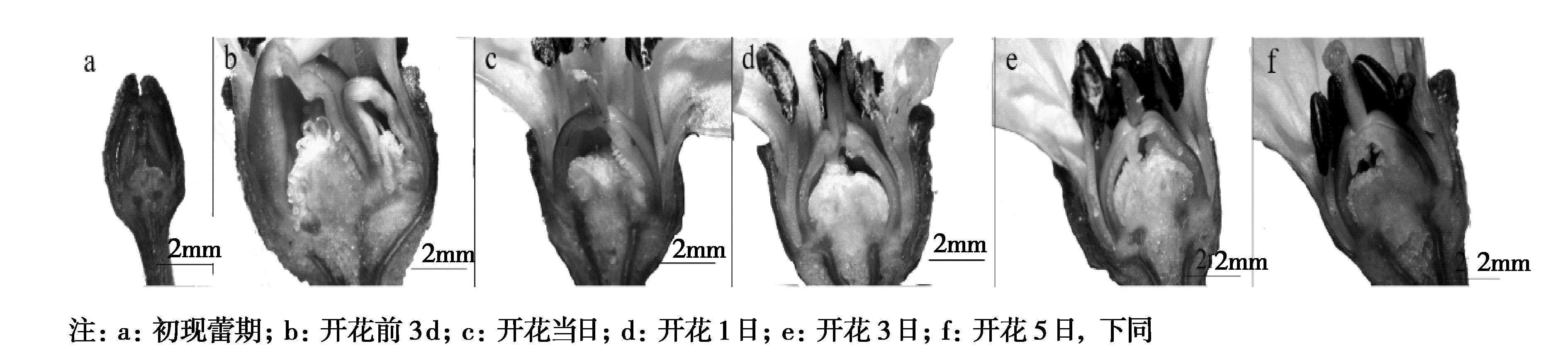
图2 低温胁迫下乐都长辣椒花器官结构Fig.2 Ledulong peppe flower organ structure under low temperature stress

图3 低温胁迫下华美105花器官结构Fig.3 Huamei 105 flower organ microstructure under low temperature stress
2.2 低温胁迫对辣椒表型及生物量的影响
由图4可知,低温对辣椒表型有明显的影响,在低温处理下,两种材料生长变缓。华美105辣椒在处理25 d后株高、完全功能叶片数、鲜重、干重较对照分别降低63%、39.3%、63.5%、73.7%;乐都长辣椒在低温处理25 d后株高、完全功能叶片数、鲜重、干重较对照分别降低50%、30%、64.9%、71.4%。但是华美105在低温条件下生物量积累优于乐都长辣椒,其它指标乐都长辣椒受温度影响比华美105较小。由此看见,低温条件下两个品种的株高、功能叶片数、鲜重、干重均低于对照。
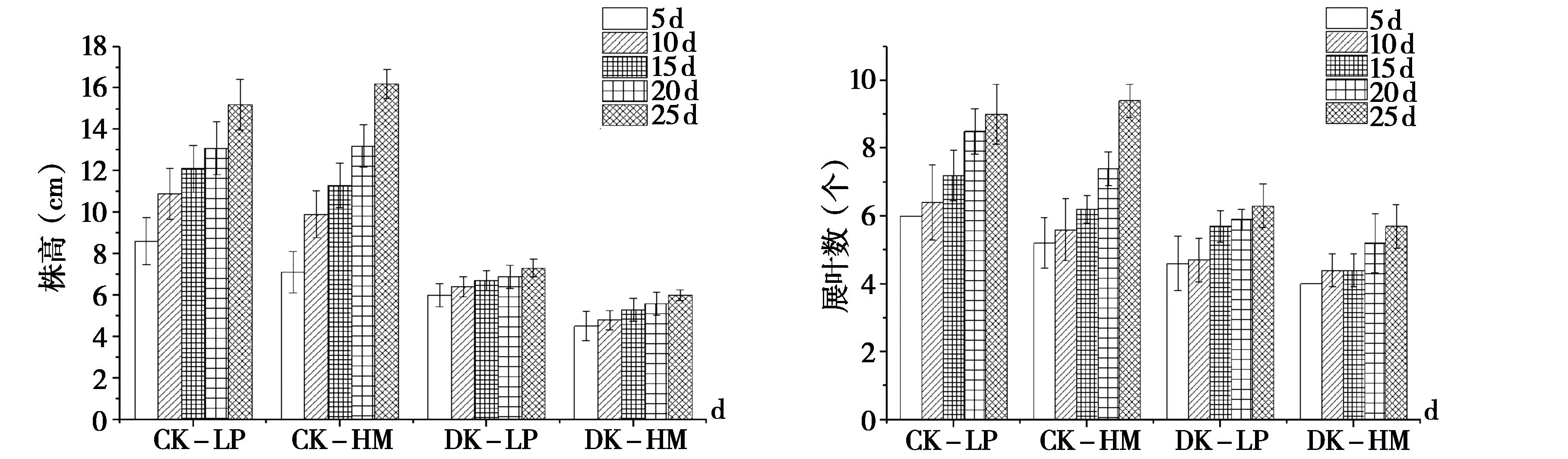
图4 不同温度处理对辣椒株高、完全功能叶数、鲜重、干重的影响Fig.4 The effects of high,leaf,fresh weight,and dry weight of chili pepper were treated at different temperatures
2.3 低温胁迫对辣椒叶绿素含量及可溶性糖含量的影响
由图5a可知,常温条件下两个品种叶绿素含量均随时间延长不断增加,25 d时都达到最大值。低温处理下两个品种叶绿素含量呈先降低后升高的趋势,在低温处理的第10 d时两个品种的叶绿素含量值均最低,然后又随着时间延长叶绿素含量逐渐增加,但是低温处理的叶绿素含量始终低于常温条件下两个品种叶绿素含量,且华美105的叶绿素含量始终高于乐都长辣椒。
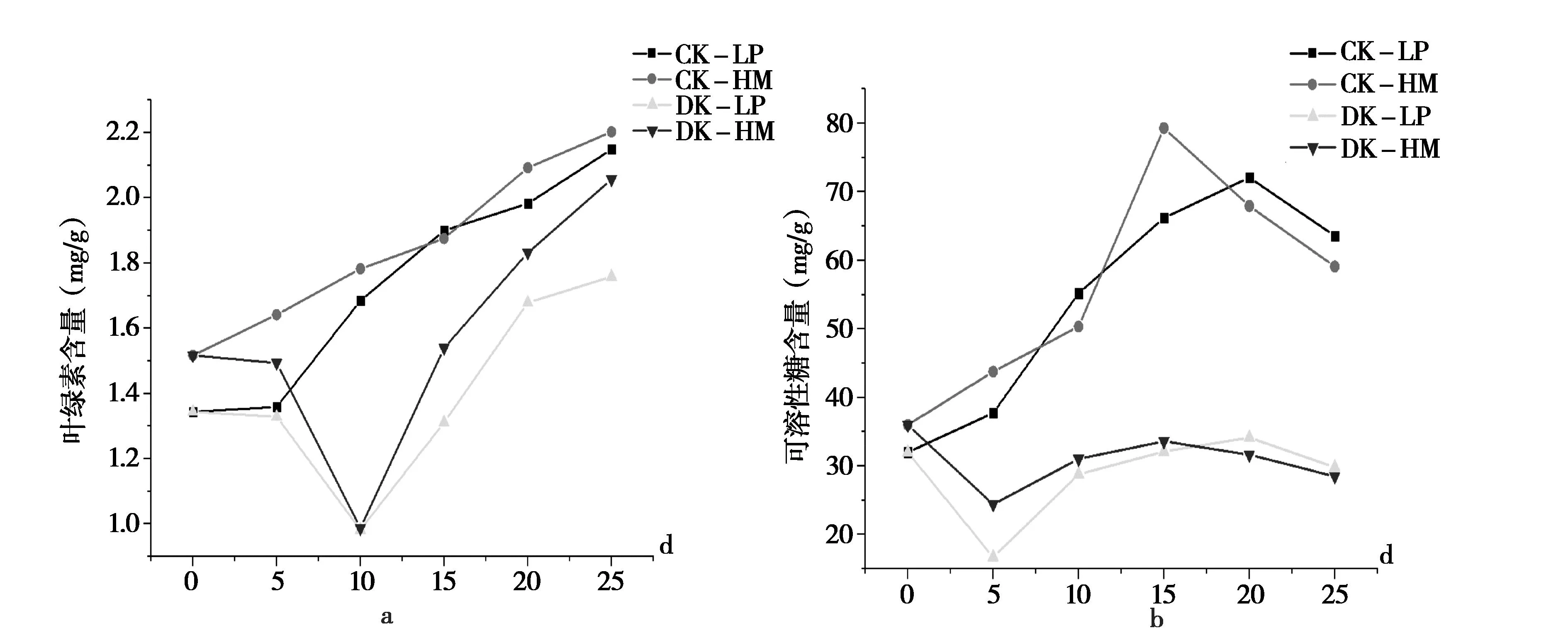
图5 不同温度处理对辣椒叶绿素和可溶性糖含量的影响Fig.5 Effects of different temperature treatments on the content of capsaicin and soluble sugars
由图5b可知,常温下随着时间的推移,两个材料的可溶性糖含量整体上均呈先上升后下降趋势;而低温下两种材料可溶性糖含量呈现先降低再缓缓增加后降低的趋势,5 d后降低到最小,但是两个品种低温处理的可溶性糖含量始终明显低于常温条件下可溶性糖含量。
由此可知,低温处理可使得叶绿素含量和可溶性糖含量下调,两个品种相比,华美105叶绿素含量和可溶性糖含量降幅相对较小,说明乐都长辣椒中叶绿素含量和可溶性糖变化对低温处理更敏感。
2.4 低温胁迫对辣椒抗氧化酶活性的影响
由图6可知,低温条件下,随着生长期延伸,两个品种辣椒叶片中 SOD活性均是先下降后上升的趋势,且培育15 d后达到最低值。与华美105相比,乐都长辣椒中SOD活性受温度影响变化更加明显,变化幅度更大。低温条件下, 两个品种的POD活性均是先升高后下降,低温处理的POD活性均高于常温的两个品种。两个品种CAT活性整体上表现出先降低后升高趋势,低温条件与常温条件相比CAT酶活性明显降低,且乐都长辣椒低温处理后变化幅度更加明显。在低温条件下,两个品种的APX活性在常温条件下表现出先下降后升高再下降的趋势。APX活性在处理的前10 d酶活性降低,之后与常温条件酶活性变化趋势一致。

图6 不同温度处理对辣椒抗氧化酶活性的影响Fig.6 Effects of different temperature treatmented on antioxidant enzyme activity of chili pepper
由此可知,低温处理下,除POD酶活性增强外,其它3种抗氧化酶活性均降低。整体上乐都长辣椒抗氧化酶活性对低温响应更敏感。
3 讨论与结论
综上所述,通过对两种耐低温辣椒品种进行低温胁迫处理发现,华美105不像乐都长辣椒,当季未发生明显的落花落果现象;两个品种的株高、功能叶片数、鲜重、干重均低于对照;华美105叶绿素、可溶性糖、SOD、CAT和APX含量降幅相对小,整体上乐都长辣椒对低温响应更敏感。
通过本研究发现,低温敏感型品种乐都长辣椒和耐低温结实型品种华美105在花蕾期生长速度基本一致,开花以后到花后5 d二者的发育速度明显不一致,其表现为乐都长辣椒子房体发育迟缓,花瓣脱落期延后,华美105在花后3 d花瓣脱落,第5 d子房体明显长大。对乐都长辣椒低温落果解剖发现,子房体中只有少数胚珠颜色正常,其余胚珠全部褐化,由此可知低温敏感型辣椒品种在冬季低温设施条件下的落花落果可能是因为花粉萌发和花粉管伸长受阻,进而不能完成受精作用或对胚珠刺激而致使子房体发育畸形或停止,变现为果实变小皱缩和脱落。这一结果与王兴娥[9]的研究结果一致。
本研究发现,在低温胁迫下,乐都长辣椒和华美105的生长均受到一定程度的影响,整体上低温下华美105的生长量积累要高于乐都长辣椒,从叶片数和株高方面而言,乐都长辣椒受到影响表现较低,这一结果和宋静爽等[10]在辣椒苗期对低温胁迫的响应的研究结果存在一定的差异,可能与本试验中处理时间长度和处理温度设置有关,需进一步验证。研究发现低温处理会导致辣椒苗期叶绿素含量降低,这与常静等[11]研究发现低温弱光胁迫对辣椒叶片叶绿素光合荧光参数及部分生理特性均存在一定程度的影响一致。叶绿素作为光合作用的重要色素,其合成经常会受到长期亚低温处理和短期低温处理的影响。有研究表明,低温导致叶绿素含量下降的观点有两种:一是叶绿素的减少,既是因为色素的合成受到阻碍,又是因为低温对叶绿素的伤害;二是低温抑制了叶绿素的合成,而非叶绿素的破坏[12-14],本研究中低温敏感品种叶绿素降低幅度大于耐低温品种,结果一定程度上支持上述理论。本研究发现低温敏感品种的可溶性糖在低温条件下变化幅度很大,这可能是这类品种在低温胁迫初期的应激反应,因为可溶性糖含量增加有利于提高细胞的修复,所以低温敏感品种可溶性糖含量应激增加是为了保护细胞免遭低温损伤,使植物体能够完成正常的生理代谢;相对而言耐低温品种的可溶性糖变幅较小,其应激反应比较温和。
低温胁迫使植物体产生破坏蛋白质、脂质、膜结构和核酸等细胞成分的活性氧,植物体细胞通过生成SOD、POD、CAT等小分子抗氧化酶,消除自由基使之保持在一定水平,防止活性氧对植物造成过度伤害[15]。研究中发现两个品种低温胁迫下SOD和CAT活性是先降低后增高,POD和APX活性是先增高后降低,整体上华美105在低温胁迫下四种酶活性变化幅度均小于乐都长辣椒,反映了两种不同抗性辣椒品种之间的应激酶活性差异。吴琴等对抗氧化酶活性的研究发现,SOD能通过歧化作用将有害环境中产生的超氧自由基去除,避免植物体内O2-和H2O2结合,最终形成羟自由基-OH,对细胞造成损伤[16]。SOD催化歧化反应生成的H2O2可被其它酶去除,但CAT只存在于过氧化酶体中,而细胞质中的H2O2不会被CAT清除,所以在细胞质中存在的APX以抗坏血酸为底物来清除H2O2。在本试验中,POD活性在低温处理全过程中都表现为高于对照,SOD、APX和CAT酶活性在低温处理时均低于对照,说明辣椒可以通过抗氧化酶活性的提高来缓解低温初期损伤。在低温过程中两个品种的SOD活性在短暂下降后又表现出上升,说明在其它抗氧化酶作用变弱后SOD才启动保护机制,但随着胁迫作用的不断持续,POD和APX保护作用逐渐降低,而CAT作为抗氧化酶体中的保护性机制也同时启动,SOD的作用也持续加强,形成比较持久的保护机制。
笔者对低温处理后移栽至田间的辣椒落花现象进行观察统计发现:乐都长辣椒85%的门椒脱落,而对椒未受到低温影响表现正常生长;华美105门椒未脱落,说明低温胁迫早期容易造成落花落果,随着多个抗氧化保护机制的启动,植株在一定程度上可以适应低温,从而减少落花落果,耐低温结实品种表现更加优良。下一步将对低温胁迫下辣椒落花、假果形成过程中内源激素水平,以及不同品种相应低温关键基因表达水平方面开展研究,为今后的育种工作提供更加科学的理论支持。
猜你喜欢
Chinese Physics B(2023年2期)2023-03-13
今日农业(2022年1期)2022-06-01
扬子江(2018年4期)2018-09-26
小学生学习指导(低年级)(2017年9期)2017-08-07
兽医导刊(2016年12期)2016-05-17
电线电缆(2016年5期)2016-02-27
橡胶工业(2015年5期)2015-08-29
橡胶工业(2015年10期)2015-08-01
传奇故事(破茧成蝶)(2015年1期)2015-02-28
食品科学(2013年19期)2013-03-11
