
不同虫态稻虱缨小蜂低温贮藏后的适合度
2023-12-25 11:45张居念吴春珠施龙清解振兴姜照伟占志雄
中国生物防治学报 2023年5期
张居念,连 玲,吴春珠,董 萌,施龙清,解振兴,姜照伟,占志雄
(福建省农业科学院水稻研究所,福州 350018)
水稻是世界上最重要的主粮作物之一,在世界各地普遍种植,全球半数以上的人口以稻米作为主食[1,2]。水稻主要害虫稻飞虱属半翅目Hemiptera,飞虱科Delphacidae,包括褐飞虱Nilaparvatalugens、白背飞虱Sogatellafurcifera和灰飞虱Laodelphaxstriatellus三种,在全球水稻产区常暴发成灾,严重影响全球粮食安全[3]。化学杀虫剂的大量持续、不合理施用等原因,使得稻飞虱对各类杀虫剂,如有机磷类、烟碱类、氨基甲酸酯类等农药的抗性逐渐增强[4,5],部分种群甚至达到成百上千倍的高水平抗性。化学农药的使用除导致害虫产生抗药性,污染环境和再猖獗的“3R”问题,还使田间自然天敌种群受到严重损害,天敌的缺乏致使害虫失去有效控制而暴发成灾。随着人们对无公害无污染及有机农产品需求量的增加,寻求有效、持久的治虫方法才是长久之计。
稻虱缨小蜂AnagrusnilaparvataePang and Wang 是稻飞虱的优势卵寄生蜂,稻田中稻飞虱卵的缨小蜂寄生率普遍很高(20.9%~83.3%)[6-8],对三种稻飞虱均有明显的控制作用,但由于稻虱缨小蜂种群的发展滞后于稻飞虱,在稻飞虱迁入稻田早期,稻虱缨小蜂群体较小,不足以控制稻飞虱的繁殖。若用人工饲养的稻虱缨小蜂进行田间释放,弥补自然种群的不足,则可望实现生物防治达到控害的目的。
在生物防治中,积累足量的天敌昆虫是大规模释放时的前提条件[7]。因此,储藏技术对天敌昆虫的规模化生产极为重要[8,9]。低温冷藏是积累和保存天敌行之有效的方法[10]。冷藏可以积累天敌的数量,同时也可以减少维持天敌种群的成本[9]。稻虱缨小蜂在饲养过程中,经常遇到羽化高峰期与寄主飞虱的产卵期不相遇,造成成虫没有可寄生的寄主卵,造成浪费。通过冷藏调节缨小蜂的发育进度,使得每一批寄主飞虱卵都可以与缨小蜂的羽化期相吻合,可提高繁蜂效率,确保天敌种群数量的充足和稳定,还可以延长货架期保障长距离运输,并且可在稻飞虱迁入产卵期实现同步释放。
天敌昆虫在低温条件下长时间贮藏,会发生冷害和负面作用,甚至导致天敌昆虫死亡[10],因此寻找天敌昆虫在低温条件下的适合虫态显得尤为重要。目前国内外有关稻虱缨小蜂人工饲养方面的研究较少,特别是冷藏方面无相关报道。本研究以羽化率、性比、雌成虫寄生飞虱卵数量、成虫寿命、繁殖力作为指标,测定不同虫态稻虱缨小蜂对不同低温、不同冷藏时间的耐受反应,探索合适的低温冷藏条件和方法,为稻虱缨小蜂的规模化饲养提供理论依据。
1 材料与方法
1.1 供试材料
稻虱缨小蜂于2018 年采自福建省农业科学院水稻研究所试验稻田,用褐飞虱卵为寄主连续继代繁殖50 多代。寄主灰飞虱由福建农林大学植物保护学院病毒研究所提供,用抗寒小麦品种鲁原118(网购)繁殖10 多代。
水稻品种TN1 每7 d 播种一次,生长条件:温度(28±3)℃、光周期16L∶8 D、RH 80%±10%,生长30~40 d,TN1 用于褐飞虱的饲养,所得褐飞虱卵用于稻虱缨小蜂的大量繁殖。小麦抗寒品种鲁原118在温度(26±3)℃,光周期10L∶12D,RH 80%±10%条件下生长30~40 d,作为灰飞虱的产卵载体用于稻虱缨小蜂的冷藏。以褐飞虱或白背飞虱为寄主,以水稻苗为产卵载体的繁殖方法是稻虱缨小蜂理想的大量快速繁殖方法,但由于水稻苗不耐低温,水稻苗在长期低温条件下造成死亡,冷藏结束后取出,在常温下容易失水或发霉,造成水稻苗叶鞘内的稻虱缨小蜂也即失水而死亡,因此,水稻苗不能作为产卵载体用于稻虱缨小蜂的冷藏。小麦苗具有耐冷性,长期冷藏后可处于鲜活状态,从冷藏库取出后,在常温条件下,不会因失水造成小麦苗组织内的稻虱缨小蜂脱水死亡,是理想的冷藏材料,以灰飞虱为寄主,以小麦苗为产卵载体的冷藏方法能够大幅度延长稻虱缨小蜂的贮藏时间,便于虫源的积累,提高繁蜂效率,实现规模化人工饲养。
1.2 各发育历期的形态特征观察
卵:将雌成虫置于载玻片上,滴加生理盐水,用解剖针将雌成虫腹部剖开,将卵转移到另一载玻片上,盖上盖玻片,用显微镜观察并拍照。幼虫:解剖被稻虱缨小蜂寄生3 d、5 d、7 d 的灰飞虱卵,其中的幼虫即为对应的1 d、3 d、5 d 龄的幼虫,将幼虫转移到有生理盐水的凹载玻片上,在幼虫上滴加少许酒精,使其沉入水中,盖上盖玻片,进行观察和拍照。蛹、隐羽化期成虫、成虫:直接将蛹、隐羽化期成虫和成虫放在体视显微镜下观察和拍照。
1.3 各虫态历期的划分
参考罗肖南[11]缨小蜂研究的结果并结合前期的试验结果,在温度26 ℃、光周期14L∶10D,RH 80%±10%条件下,将寄生2 d 内的虫态划为卵期,寄生3~7 d 为幼虫期,寄生8~9 d 为蛹期,寄生10 d 为隐羽化期,羽化出壳后为成虫期。
1.4 各虫态冷藏后对适合度的影响
将小麦苗剥去黄叶老叶,接入怀卵灰飞虱(腹部明显彭大),每棵麦苗2 只,用直径20 mm,长20 cm亚克力圆管罩住,上端用100 目尼龙网封口,产卵1 d 后,移除灰飞虱成虫,接入初羽化稻虱缨小蜂1 对(雌、雄各1 只为1 对),寄生第2 d 去除成虫,分别在寄生1 d、2 d、3 d、4 d、5 d、6 d、7 d、8 d、9 d、10 d 后,放入温度为5 ℃、8 ℃、11 ℃、12 ℃、26 ℃(CK),光周期14L∶10D 培养箱中贮藏20 d、40 d、60 d 后取出,用0.02%高锰酸钾溶液表面消毒后,在体视显微镜下将麦苗中的灰飞虱卵用消毒过的镊子全部挑出,置于灭菌过的载玻片上,放在直径9 cm 垫有滤纸的培养皿里,加适量无菌水保湿培养。培养条件:温度26 ℃、光周期14L∶10D,RH 80%±10%。羽化后每天记载羽化数量和性比,并将当天羽化的稻虱缨小蜂1 对转移到前一天产过灰飞虱卵的新麦苗上,让其寄生,5 d 后剥查寄生数量(被寄生的卵为橙红色),调查子代繁殖力。其余缨小蜂转移到新培养皿里观察成虫寿命,每个处理调查5 株麦苗,并重复3 次。成虫的冷藏:用4.5 mL 四面透明的一次性比色皿收集刚羽化的成虫,用棉花塞住管口,置3 ℃、5 ℃、8 ℃、11 ℃、12 ℃培养箱贮藏10 d、15 d、20 d,统计存活率后,将存活的成虫转移至前一天产过灰飞虱卵的麦苗上,每株麦苗接种1 对成虫,5 d 后剥查寄生数量,调查冷藏后的繁殖力(未经冷藏的作为对照),每管比色皿装10 只成虫,每个处理3 管,重复3 次。为便于描述,以下将温度26 ℃、光周期14L:10D,RH 80%±10%条件下的培养称为正常培养。
1.5 数据处理与统计分析
用Excel 和SPSS 统计软件进行数据处理,数值采用平均值±标准误(mean±SE)表示,P值小于0.05表示差异达显著水平,图表中的不同英文小写字母表示在0.05 水平下差异显著(LSD 法)。
2 结果与分析
2.1 稻虱缨小蜂各虫态的形态特征
稻虱缨小蜂的卵呈椭圆形,一端细长,柄状。长(含柄)121~186 µm,宽22~28 µm。幼虫前期(孵化2 d)体色透明,头部前端钝平,两侧有一对无色突起,口器退化,仅一对褐色上颚清晰可见;胸部3节,腹部分节不明显,尾部有一对无色突起,幼虫后期(孵化3~5 d)形体与前期相似,体内油球增多,颜色逐渐加深,呈橙红色或黄绿色(图1B、C、D)。蛹为离蛹,橘红色,头胸腹分化明显,单眼紫红色,位于两复眼中间,三角形排列(图1E)。隐羽化期为羽化未出壳期,此时虫体与成虫无异,触角、口器、复眼、单眼、翅、产卵器等均清晰可见(图1F)。雌成虫体长0.54 mm,橙红色,复眼黑褐色,单眼红色。触角9 节,梗节扁状,侧面梨形,内侧具若干平行横斜纹,第3 节最短,短于梗节的一半,棒节纺锤形,长为宽的3 倍。前翅较后翅宽,后翅狭窄。产卵器向前达腹部基部,向后伸出于腹部末端(图1G)。雄成虫形态与雌成虫相似,体形略小,触角13 节鞭状,第Ⅲ节稍短于其远端的各节,第IV~Xlll 各节基本等长(图1H)。
2.2 卵期不同冷藏处理对适合度的影响
2.2.1 不同冷藏处理对卵孵化发育的影响 经三因素方差分析,其结果表明,冷藏温度(F=1414.064;P=0.00)、冷藏时间(F=505.445;P=0.000)和冷藏温度×冷藏时间(F=59.949;P=0.000)对稻虱缨小蜂卵的孵化发育产生显著影响,冷藏卵龄(F=0.963;P=0.331)、冷藏温度×冷藏卵龄(F=0.569;P=0.638)、冷藏时间×冷藏卵龄(F=1.977;P=0.149)和冷藏温度×冷藏时间×冷藏卵龄(F=0.442;P=0.847)均对孵化发育无显著影响。
灰飞虱卵被稻飞虱缨小蜂寄生后,正常培养7 d,在体视显微镜下,可以清晰地观察到被寄生卵由半透明基本无色转变成橙红色或浅黄绿色。在5 ℃条件下,冷藏20 d 以上,再正常培养7 d 后,未剥查到转色的被稻虱缨小蜂寄生的灰飞虱卵,说明稻虱缨小蜂的卵在上述条件下处理后,不能孵化或孵化后不能正常发育。在8 ℃条件下,冷藏40 d 内,有部分寄生卵可正常孵化和发育,相同冷藏时间处理的1 d 与2 d卵龄的正常发育数量之间无显著差异。冷藏20 d,寄生卵能正常发育的数量与对照相比,只有43.89%。冷藏60 d 后,寄生卵不能正常发育。在11 ℃条件下,冷藏60 d 内,均有部分寄生卵能正常发育,但随冷藏时间的延长,正常发育的数量逐渐下降。在12 ℃下冷藏,发育数有所增加,但冷藏时间最短的20 d,正常发育的数量也只有对照的76.82%。说明不适合在卵期进行冷藏(表1)。
2.2.2 卵期不同冷藏处理对成虫寿命、成虫性比和子代数量的影响 卵期不同冷藏处理经三因素方差分析,冷藏温度(F寿命=7.999;P寿命=0.002)和冷藏时间(F寿命=97.136;P寿命=0.000)对成虫寿命产生显著影响,卵龄(F寿命=0.205;P寿命=0.654)对成虫寿命无显著影响,冷藏温度×卵龄(F寿命=1.606;P寿命=0.216)和冷藏温度×冷藏时间×卵龄(F寿命=0.449;P寿命=0.720)也无显著影响。
冷藏20 d 内,只有12 ℃下的成虫寿命与对照相比有显著差异,8 ℃和11 ℃的成虫寿命与对照相比无显著差异。8 ℃~12 ℃范围内,成虫寿命随温度的升高而缩短,也随冷藏时间的延长而缩短,12 ℃冷藏60 d 后寿命与对照相比显著缩短,缩短了1.20 d。
卵期不同冷藏处理经三因素方差分析,冷藏温度(F♀=0.110;P♀=0.896)、冷藏时间(F♀=2.547;P♀=0.094)和卵龄(F♀=0.124;P♀=0.727)对成虫的性比无显著影响,雌雄比接近1∶1;冷藏温度(F子代数=0.318;P子代数=0.729)、冷藏时间(F子代数=0.458;P子代数=0.637)和卵龄(F子代数=0.539;P子代数=0.468)对子代数也无显著影响(表2)。
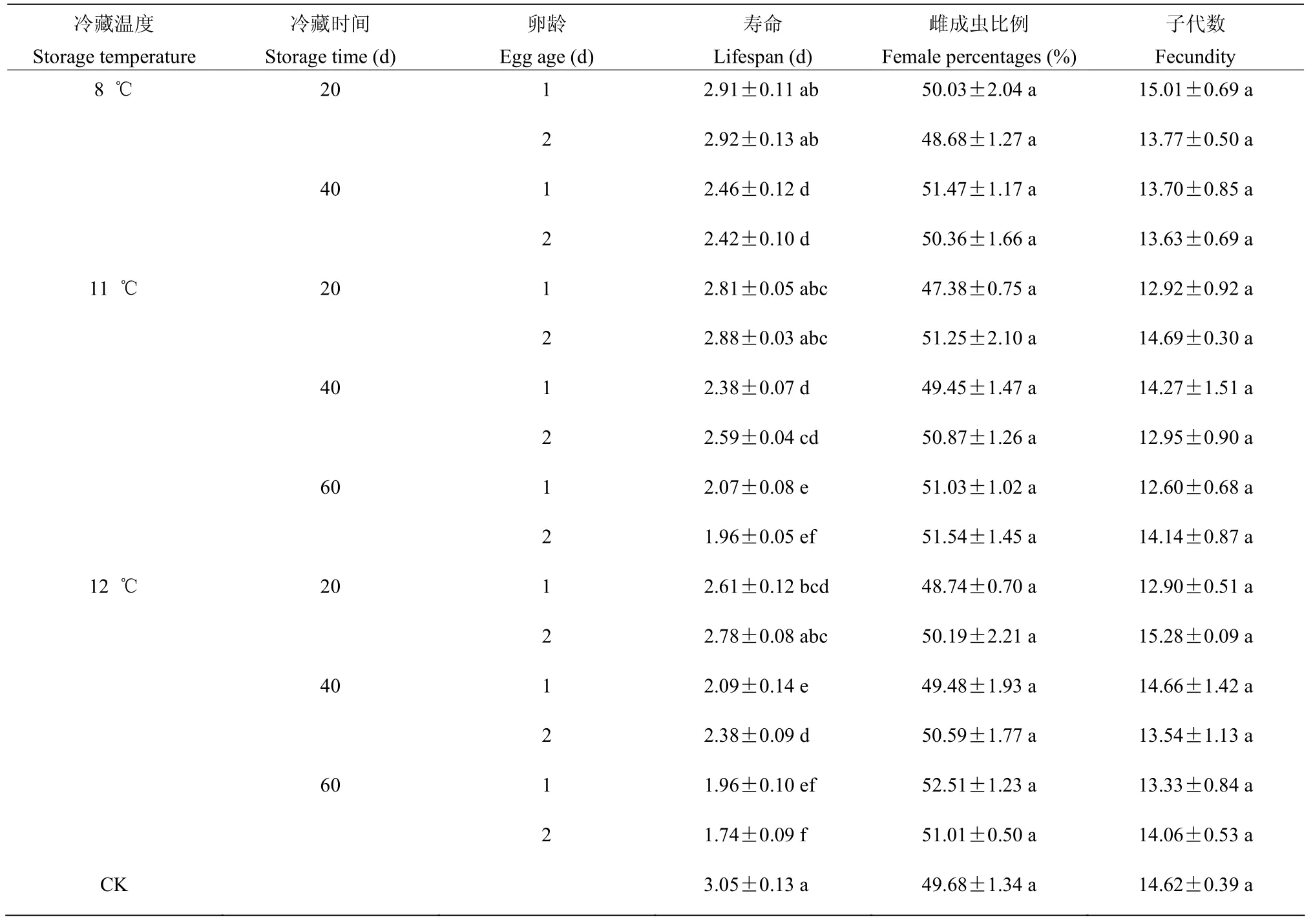
表2 卵期不同冷藏处理对稻虱缨小蜂成虫寿命、成虫性比和子代数的影响Table 2 Effects of cold storage on Anagrus nilaparvatae adult lifespan, sex ratio and fecundity at egg stage
2.3 幼虫期不同冷藏处理对适合度的影响
2.3.1 幼虫期不同冷藏处理对羽化率的影响 稻虱缨小蜂幼虫期的冷藏处理,经三因素方差分析,冷藏温度(F=486.942;P=0.000)和冷藏时间(F=1780.897;P=0.000)对羽化率产生显著影响,虫龄(F=1.285;P=0.280)对羽化率无显著影响。冷藏温度×冷藏时间(F=44.823;P=0.000)对羽化率产生了显著影响,冷藏温度×虫龄(F=0.745;P=0.705)、冷藏时间×虫龄(F=0.683;P=0.706)和冷藏温度×冷藏时间×虫龄(F=0.778;P=0.757)均对羽化率无显著影响。
羽化率随着冷藏时间的延长而下降,但在不同温度下,下降幅度有所不同,温度越低,下降幅度越大。在相同冷藏温度下,幼虫期不同虫龄之间的羽化率差异不显著。在5 ℃~12 ℃范围内,11 ℃和12 ℃冷藏的羽化率受低温时间的影响最小,冷藏20 d 的平均羽化率高于80.00%,11 ℃和12 ℃冷藏40 d 的平均羽化率分别为64.85%和64.95%,两者相当,但12 ℃冷藏60 d 后的平均羽化率大幅度下降,为34.93%,11 ℃冷藏60 d 后的平均羽化率还可保留有50.69%,因此,从羽化率这个因素考虑,11 ℃是合适的冷藏温度(表3)。
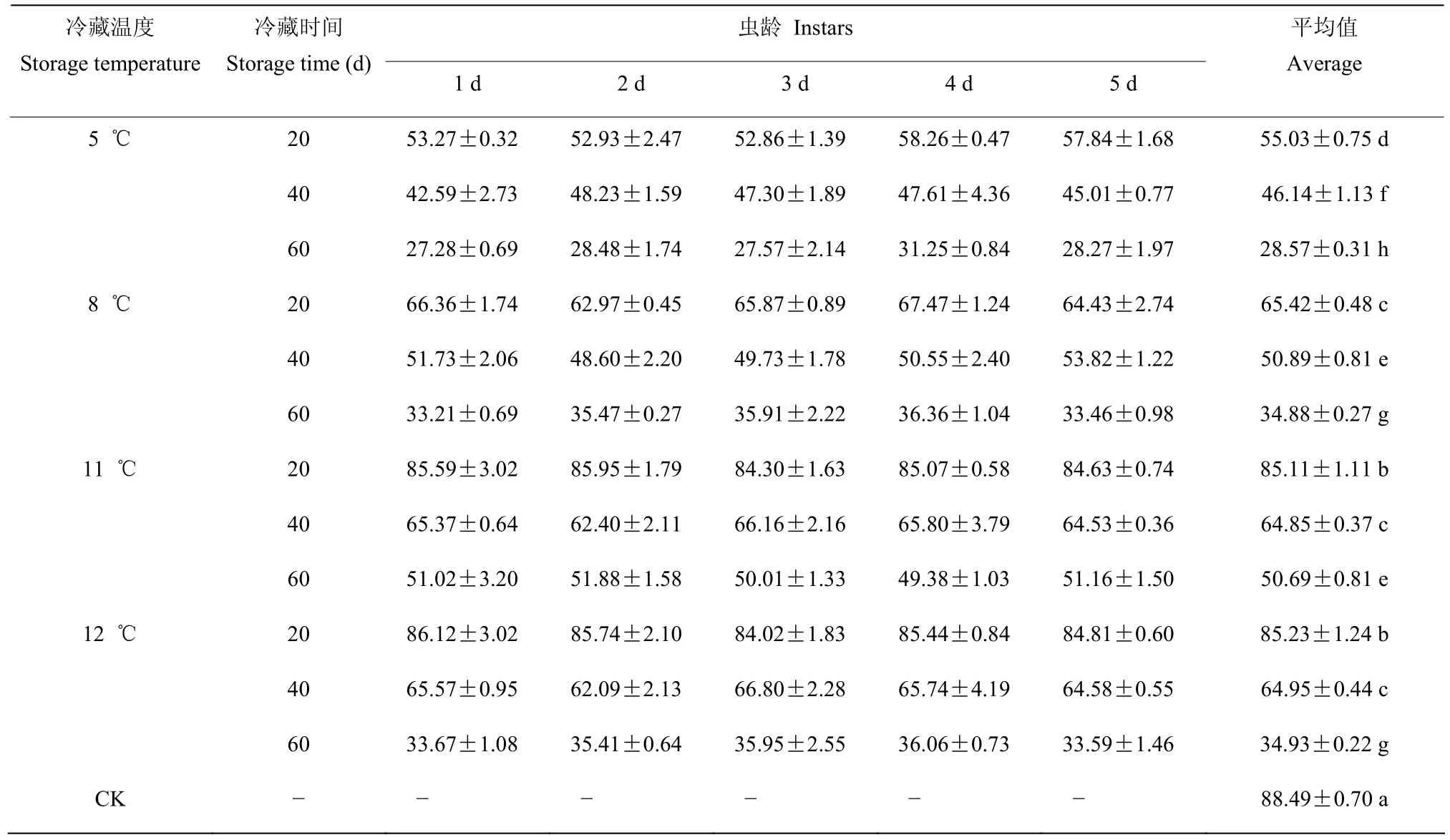
表3 稻虱缨小蜂幼虫期各虫龄冷藏后的羽化率Table 3 The average eclosion rate of Anagrus nilaparvatae after cold storage at larvae stage (%)
2.3.2 幼虫期不同冷藏处理对成虫性比的影响 经三因素方差分析,其结果表明,冷藏温度(F=0.883;P=0.452)、冷藏时间(F=0.302;P=0.740)、虫龄(F=0.531;P=0.713)、冷藏温度×冷藏时间(F=1.445;P=0.203)、冷藏时间×虫龄(F=0.718;P=0.675)和冷藏温度×冷藏时间×虫龄(F=0.518;P=0.968)均对成虫的性比无显著影响。同样,所有处理的雌虫比例与对照组均无显著性差异(表4),雌雄比例接近1∶1。由此可见,幼虫期低温贮藏并不会对稻虱缨小蜂的成虫性比产生影响。

表4 稻虱缨小蜂幼虫期各虫龄冷藏后的雌成虫比例Table 4 Female percentages of Anagrus nilaparvatae after cold storage at larvae stage (%)
2.3.3 幼虫期不同冷藏处理对成虫寿命的影响 经三因素方差分析结果表明,冷藏温度(F=230.218;P=0.000)、冷藏时间(F=124.196;P=0.000)和冷藏温度×冷藏时间(F=8.745;P=0.000)对稻虱缨小蜂的成虫寿命产生显著影响,虫龄(F=1.042;P=0.388)、冷藏温度×虫龄(F=1.255;P=0.255)、冷藏时间×虫龄(F=0.722;P=0.671)和冷藏温度×冷藏时间×虫龄(F=1.097;P=0.358)均对寿命无显著影响。
在5 ℃~12 ℃范围内,成虫寿命随着温度的升高而缩短,随冷藏时间的延长而缩短,不同虫龄在相同温度相同冷藏时间的寿命差异不显著。虽然不同温度和不同冷藏时间之间差异显著,但下降幅度较小,寿命最短的11 ℃冷藏60 d 的寿命2.46 d,只比对照短了0.66 d(表5)。

表5 稻虱缨小蜂幼虫期各虫龄冷藏后的成虫寿命Table 5 Adult lifespan of Anagrus nilaparvatae after cold storage at larvae stage (d)
2.3.4 幼虫期不同冷藏处理对子代数量的影响 经三因素方差分析结果表明,冷藏温度(F=0.670;P=0.572)、冷藏时间(F=0.792;P=0.455)、虫龄(F=1.289;P=0.278)、冷藏温度×冷藏时间(F=0.378;P=0.892)、冷藏温度×虫龄(F=0.219;P=0.997)、冷藏时间×虫龄(F=0.399;P=0.919)和冷藏温度×冷藏时间×虫龄(F=1.090;P=0.366)均对子代数量无显著影响,所有处理的子代数量与对照均无显著性差异(表6),由此可见,幼虫期低温贮藏不会对稻虱缨小蜂的子代数量产生显著影响。
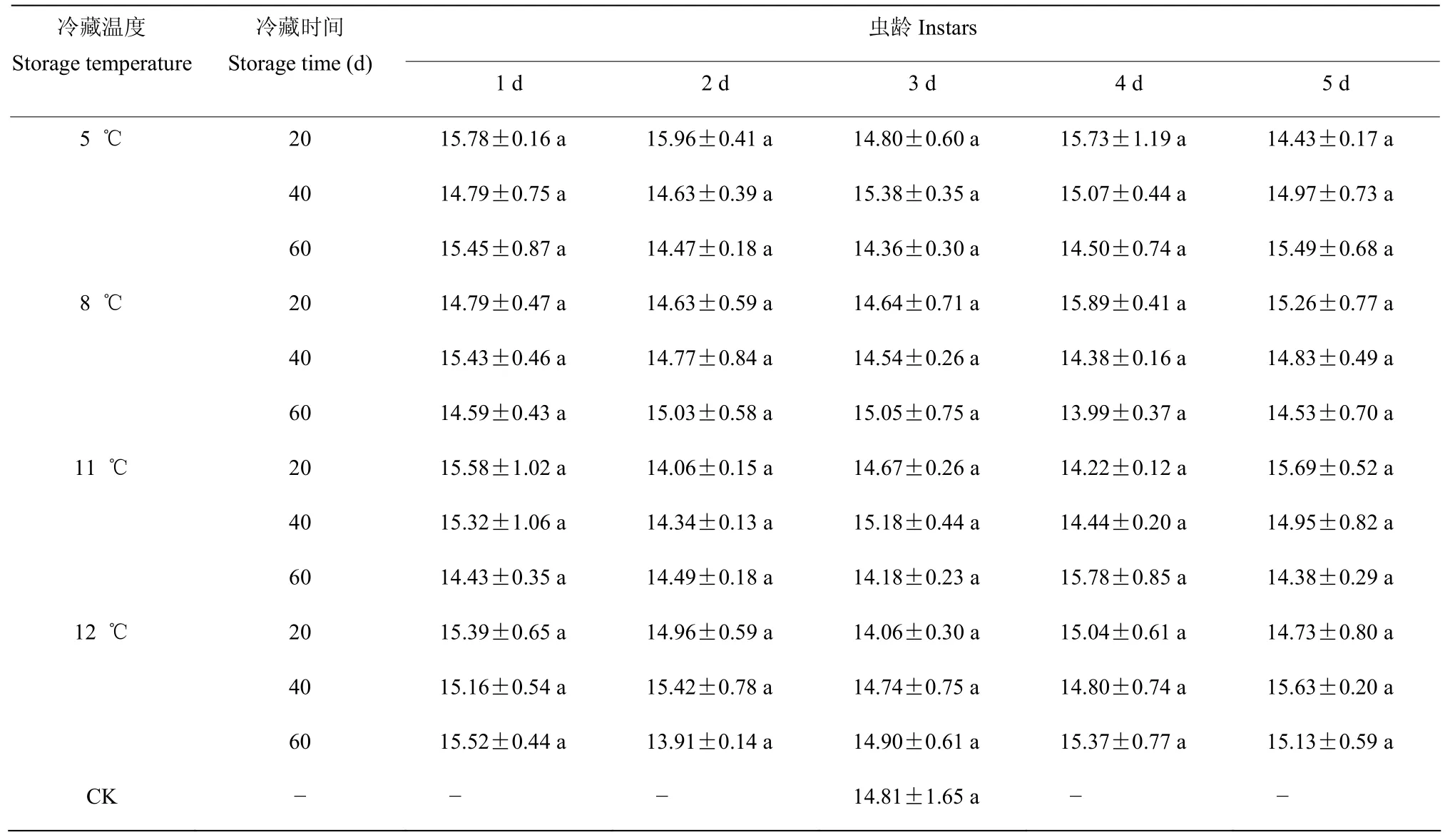
表6 稻虱缨小蜂幼虫期各虫龄冷藏后的子代数量Table 6 Fecundity of Anagrus nilaparvatae after cold storage at larvae stage
2.4 蛹期不同冷藏处理对适合度的影响
2.4.1 蛹期不同冷藏处理对羽化率的影响 经三因素方差分析结果表明,冷藏温度(F=156.211;P=0.000)、冷藏时间(F=643.982;P=0.000)和冷藏温度×冷藏时间(F=4.264;P=0.002)对羽化率产生显著影响,蛹龄(F=0.343;P=0.561)、冷藏温度×蛹龄(F=0.205;P=0.893)、冷藏时间×蛹龄(F=1.878;P=0.164)和冷藏温度×冷藏时间×蛹龄(F=1.087;P=0.384)均对羽化率无显著影响。
蛹期冷藏后的羽化率随温度的降低而下降,随冷藏时间的延长而下降,不同蛹龄之间的羽化率没有显著差异,在11 ℃下冷藏20 d 的羽化率达82.81%(两个龄期的平均值),冷藏60 d 的羽化率也较高,为46.68%(表7)。

表7 稻虱缨小蜂蛹期冷藏后的羽化率Table 7 The average eclosion rate of Anagrus nilaparvatae after cold storage at pupal stage (%)
2.4.2 蛹期不同冷藏处理对成虫性比的影响 经三因素方差分析结果表明,冷藏温度(F=0.507;P=0.679)、冷藏时间(F=0.743;P=0.679)、蛹龄(F=0.806;P=0.374)、冷藏温度×冷藏时间(F=0.112;P=0.995)、冷藏温度×蛹龄(F=0.998;P=0.402)、冷藏时间×蛹龄(F=0.611;P=0.547)和冷藏温度×冷藏时间×蛹龄(F=0.125;P=0.993)均对成虫性比无显著影响(表8)。

表8 稻虱缨小蜂蛹期冷藏后的雌成虫比例Table 8 Female percentages of Anagrus nilaparvatae after cold storage at pupal stage (%)
2.4.3 蛹期不同冷藏处理对成虫寿命的影响 经三因素方差分析结果表明,冷藏温度(F=42.790;P=0.000)、冷藏时间(F=182.843;P=0.000)和冷藏温度×冷藏时间(F=2.590;P=0.030)对成虫寿命产生显著影响,蛹龄(F=0.004;P=0.950)、冷藏温度×蛹龄(F=1.552;P=0.213)、冷藏时间×蛹龄(F=0.084;P=0.420)和冷藏温度×冷藏时间×蛹龄(F=0.911;P=0.495)对成虫寿命无显著影响。
蛹期在5 ℃~12 ℃范围内冷藏后,成虫寿命随冷藏温度的升高而缩短,随冷藏时间的延长而缩短。1 d和2 d 两个蛹龄之间的成虫寿命差异不显著,5 ℃冷藏20 d 与8 ℃冷藏20 d、40 d 和11 ℃冷藏20 d 的成虫寿命与对照相比差异不显著(表9)。
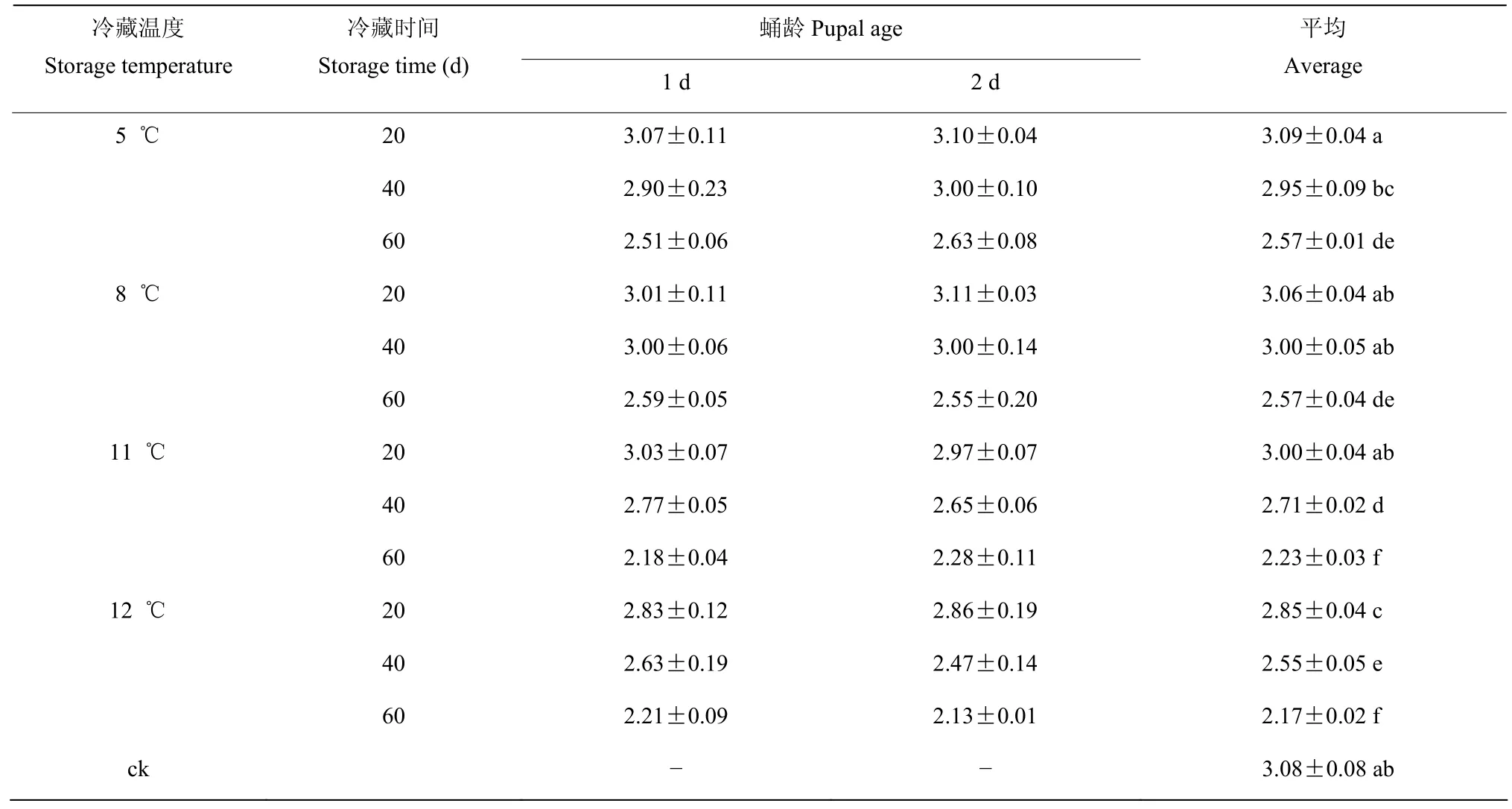
表9 稻虱缨小蜂蛹期冷藏后的成虫寿命Table 9 Adult lifespan of Anagrus nilaparvatae after cold storage at pupal stage (d)
2.4.4 蛹期不同冷藏处理对子代数的影响 经三因素方差分析结果表明,冷藏温度(F=1.127;P=0.348)、冷藏时间(F=0.822;P=0.446)、蛹龄(F=0.309;P=0.581)、冷藏温度×蛹龄(F=0.970;P=0.414)、冷藏温度×冷藏时间(F=0.956;P=0.465)、冷藏时间×蛹龄(F=0.135;P=0.874)和冷藏温度×冷藏时间×蛹龄(F=0.536;P=0.778)均对子代数量无显著影响(表10)。
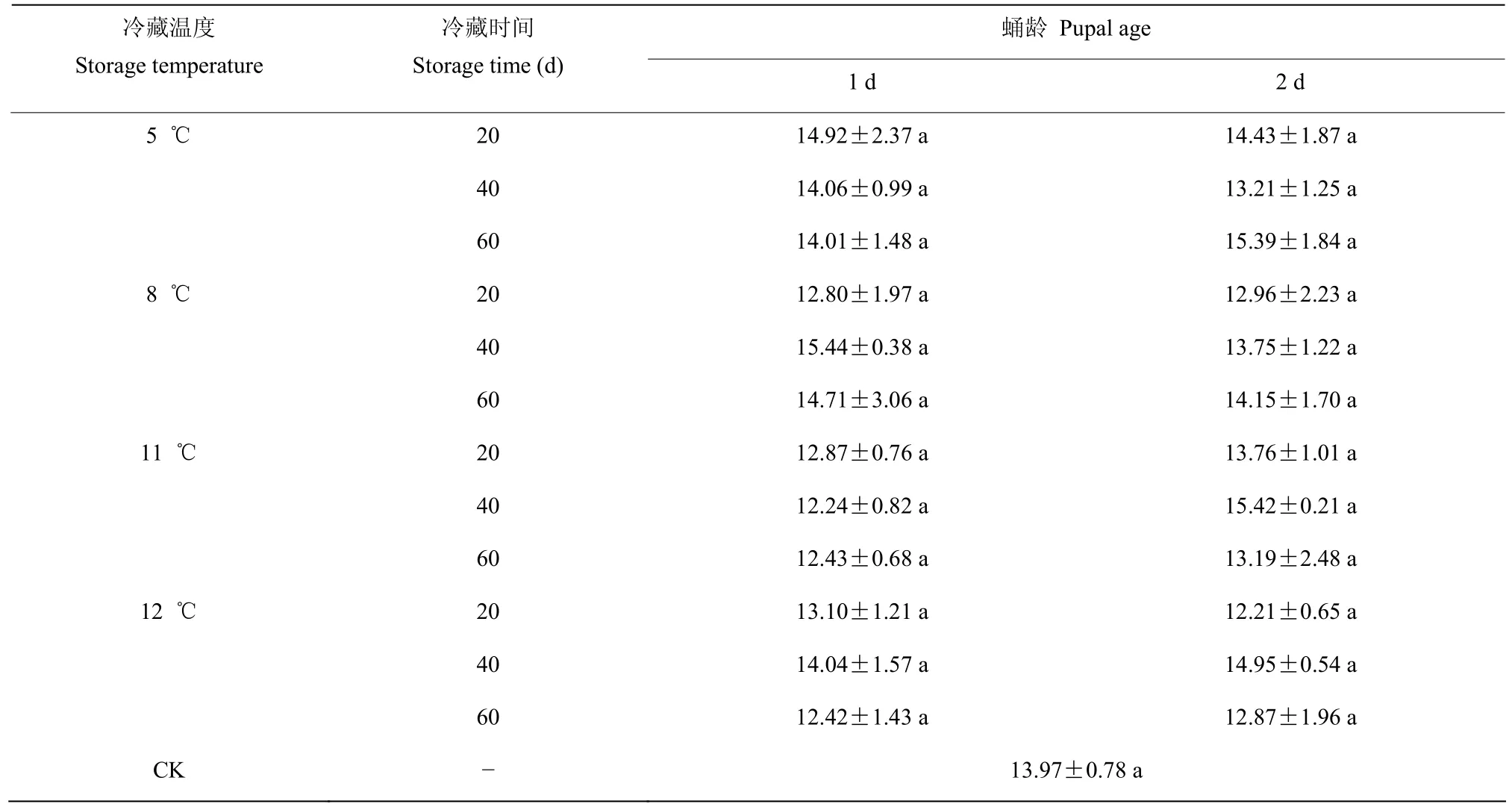
表10 稻虱缨小蜂蛹期冷藏后的子代数量(只)Table 10 Fecundity of Anagrus nilaparvatae after cold storage at pupal or cryptoeclosion stage
2.5 隐羽化期不同冷藏处理对适合度的影响
隐羽化期不同冷藏处理经两因素方差分析,冷藏温度(F羽化率=46.523;P羽化率=0.000)、冷藏时间(F羽化率=380.253;P羽化率=0.000)和冷藏温度×冷藏时间(F羽化率=20.889;P羽化率=0.000)对羽化率产生显著影响。隐羽化期冷藏后,羽化率随着冷藏时间的延长而下降,在5 ℃~12 ℃范围内,与对照相比,羽化率均显著低于对照,说明在隐羽化期冷藏,羽化率易受低温影响,不宜在此阶段进行冷藏。在不同冷藏温度下,羽化率下降幅度有所不同,相同冷藏时间条件下,11 ℃的羽化率下降幅度最小,5 ℃的下降幅度最大。
经两因素方差分析,冷藏温度(F♀=0.306;P♀=0.821)和冷藏时间(F♀=0.210;P♀=0.812)对成虫的性比无显著影响,雌雄比接近1∶1;冷藏温度(F子代数=1.426;P子代数=0.260)和冷藏时间(F子代数=0.340;P子代数=0.715)对子代数也无显著影响。
经两因素方差分析,冷藏温度(F寿命=204.158;P寿命=0.000)和冷藏时间(F寿命=100.609;P寿命=0.000)和冷藏温度×冷藏时间(F寿命=2.538;P寿命=0.048)对成虫寿命产生显著影响。冷藏后,在8 ℃~12 ℃范围内,成虫寿命随温度的升高而缩短,也随冷藏时间的延长而缩短。5 ℃冷藏20 d 内,成虫寿命与对照相当,无显著差异,其它处理的成虫寿命均比对照显著缩短,12 ℃冷藏60 d 的寿命缩短最多,缩短了1.42 d,与对照相比,缩短了53.90%(表11)。

表11 隐羽化期不同冷藏处理对稻虱缨小蜂羽化率、成虫寿命、成虫性比和子代数的影响Table 11 Effects of cold storage on Anagrus nilaparvatae eclosion rate, adult lifespan, sex ratio and fecundity at cryptoeclosion stage
2.6 成虫冷藏对适合度的影响
2.6.1 成虫冷藏后的存活率 经两因素方差分析结果表明,冷藏时间(F=2190.736;P=0.000)对存活率产生显著影响,冷藏温度(F=0.732;P=0.577)以及冷藏温度×冷藏时间(F=0.998;P=0.458)对存活率无显著影响。
在3 ℃~12 ℃范围内,相同冷藏时间不同冷藏温度之间的存活率无显著差异,相同冷藏温度不同冷藏时间之间的存活率差异显著。成虫冷藏后的存活率随冷藏时间的延长而降低,冷藏10 d 的存活率92%以上,冷藏15 d 的存活率82%以上,冷藏20 d 的存活率大幅度下降,存活率低于50%(图2)。
2.6.2 成虫冷藏后的繁殖力 经两因素方差分析结果表明,冷藏温度(F=20.517;P=0.000)、冷藏时间(F=1073.391;P=0.000)以及冷藏温度×冷藏时间(F=5.667;P=0.000)均对繁殖力有显著影响。
成虫在3 ℃~11 ℃范围内,不同温度相同冷藏时间的寄生数量之间无显著差异,相同温度不同冷藏时间的寄生数量之间存在显著差异。在3 ℃~12 ℃范围内,寄生数量随着冷藏时间的延长而显著下降,12 ℃下冷藏的寄生数量下降幅度大于其它各温度,并与其它各温度的寄生数量均存在显著差异,冷藏15 d的寄生数量大幅下降,寄生数量最多的11 ℃冷藏15 d 处理也只有对照的53.36%。在3 ℃~11 ℃范围内,冷藏10 d 的寄生数量与对照相比无显著差异,但在12 ℃下冷藏10 d 的寄生数量与3 ℃~11 ℃和对照相比均显著下降。在3 ℃~12 ℃范围内冷藏20 d,成虫寄生数量均为0,表明已完全丧失寄生能力(图3)。
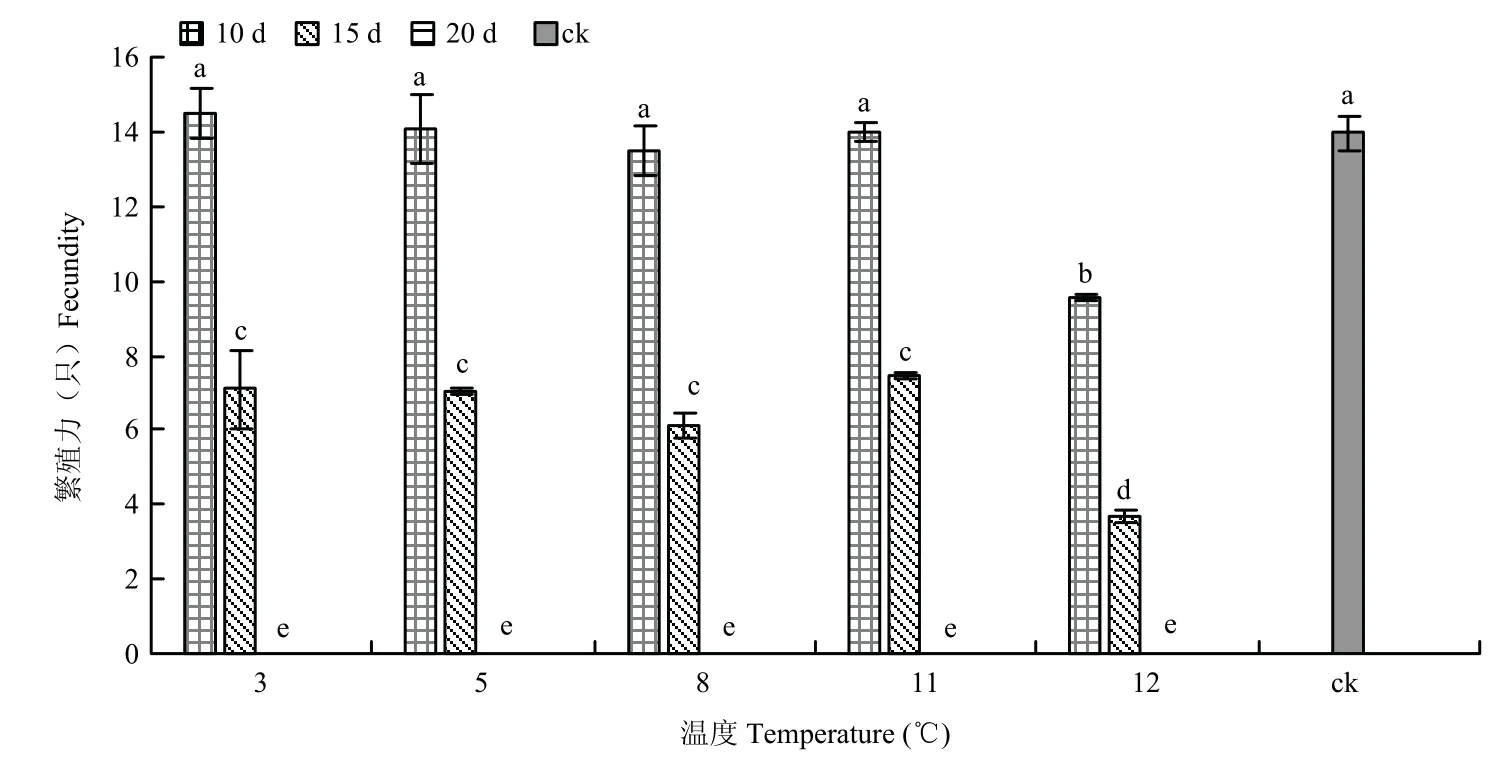
图3 稻虱缨小蜂成虫冷藏后的繁殖力Fig.3 Fecundity of Anagrus nilaparvatae after cold storage at adult stage
3 讨论
本研究稻虱缨小蜂雌成虫体长0.54 mm,短于庞雄飞等[12]记载的0.79 mm,差异较大。造成形体大小差异的原因是稻虱缨小蜂寄主较多,褐飞虱、白背飞虱、灰飞虱和小宽头飞虱等均是其寄主,成虫形体大小会因寄主卵的大小而发生变化,寄主卵大的其成虫体形相应较大。本研究中的稻虱缨小蜂是以灰飞虱为寄主饲养的,灰飞虱卵的个体比褐飞虱卵小,因此,文中所述成虫体长短于文献记载的情况是正常的。
低温贮藏可以延缓昆虫天敌的发育进度或暂停发育,能够延长昆虫天敌的保存时间,实现天敌与寄主寄生期的相遇,有利于天敌的大量积累,保证释放期有充足的天敌。但冷藏对昆虫天敌有伤害作用,冷藏温度和冷藏时间均可对天敌产生不良影响,不同虫态对低温和冷藏时间的耐受能力也不同。有些研究表明,略高于昆虫发育起点温度(T0)的低温是最佳的冷藏温度,如Anagyrusananatis、Leptomastixdactylopii、Telenomuspodisi的适宜冷藏温度均略高于发育起点温度[13-16]。而有些天敌昆虫的最适冷藏温度却低于发育起点温度。Aphidiusgifuensis、Muscidifuraxraptor和Aphidiusmatricariae的适合冷藏温度均低于发育起点温度[14-18]。一般冷藏温度越低,死亡率越高[15,18],但也有一些寄生蜂的存活率与冷藏温度(一定温度范围内)无显著相关[18,24]。在适宜的冷藏温度下,冷藏效果一般随冷藏时间的延长而下降[15,16]。本研究表明,稻虱缨小蜂的适宜冷藏温度为11 ℃,略高于发育起点温度(稻虱缨小蜂的雌雄平均发育起点温度为10.61 ℃[17]),在此温度下,稻虱缨小蜂冷藏适合度的关键因子羽化率(虽然冷藏后成虫寿命随着温度的升高而缩短,随冷藏时间的延长而缩短,但对子代数并不造成影响,而且温度和冷藏时间对性比也没有显著影响,各虫龄之间也无显著差异,因此,冷藏适合度的关键因子为羽化率)受低温和冷藏时间的影响最小,冷藏60 d 的平均羽化率还可保留50.69%。稻虱缨小蜂冷藏后的羽化率除受冷藏温度和冷藏时间的影响外,冷藏载体小麦苗对稻虱缨小蜂的羽化率也有一定的影响,小麦苗在冷藏过程中部分老叶会枯死,致使其内的稻虱缨小蜂失水死亡而降低羽化率。因此,要提高羽化率,还需筛选更耐寒的小麦品种作为稻虱缨小蜂的冷藏载体。
此外,相关研究表明不同虫态的耐寒能力存在差异,草地贪夜蛾不同虫态的耐低温能力依次为卵<成虫<幼虫<蛹[18]。双委夜蛾老熟幼虫和蛹的耐寒性最强[19],Trichogrammapintoi在0 ℃~5 ℃范围内以预蛹期和前蛹期冷藏效果最好[20],卷蛾分索赤眼蜂Trichogrammabactrae在3 ℃~5 ℃下老熟幼虫最适合于冷藏[21],麦蛾茧蜂Habrobraconbebetor则以成虫更适合冷藏[22,23],有的寄生蜂在不同的温度下适合冷藏的虫态不同,如松毛虫赤眼蜂Trichogrammadendrolimi在4 ℃下最适合冷藏的虫态是幼虫期和蛹,在7 ℃下以卵和蛹期冷藏效果最佳[24]。本研究表明在卵期进行冷藏,受冷害较严重,部分卵不能正常孵化发育,损失较大,不适合在卵期进行冷藏;在隐羽化期进行冷藏其羽化率明显下降,而成虫冷藏会丧失寄生能力,说明隐羽化期和成虫也不适合于冷藏。综上所述,稻虱缨小蜂以幼虫期和蛹期进行冷藏效果最好,冷藏温度以11 ℃最为适宜。本研究探索了稻虱缨小蜂的适宜冷藏虫态和冷藏温度,为进一步优化稻虱缨小蜂的冷藏条件奠定了工作基础,为稻虱缨小蜂的积累和延长货架期提供了一定的理论依据。
猜你喜欢
云南农业(2024年3期)2024-03-11
中国水稻科学(2023年3期)2023-05-18
广西林业科学(2021年5期)2021-11-05
昆虫学报(2021年8期)2021-09-06
昆虫学报(2020年8期)2020-10-15
湖北植保(2017年4期)2017-08-31
小溪流(画刊)(2017年6期)2017-07-10
华南农业大学学报(2015年5期)2015-12-04
阅读与作文(小学低年级版)(2015年6期)2015-05-30
生物灾害科学(2015年3期)2015-03-11
