
黄淮平原矿区土地复垦对微生物群落结构和功能的影响
2023-12-21 09:51:06董文雪朱燕峰尤云楠浮2
煤炭科学技术 2023年11期
董文雪 ,马 静,2 ,何 环 ,朱燕峰 ,尤云楠 ,陈 浮2,
(1.中国矿业大学 化工学院, 江苏 徐州 221000;2.河海大学 公共管理学院, 江苏 南京 211000;3.中国矿业大学 矿山生态修复教育部工程研究中心, 江苏 徐州 221116)
0 引 言
经济社会发展离不开能源的支撑,但化石能源开采破坏当地的生态环境[1],严重制约区域经济-社会-生态和谐和永续发展,尤其是黄淮平原矿区[2]。该区地下潜水位高、地质结构复杂、粮煤高度叠合[3],煤炭开采后地表快速沉陷,形成深1.0~4.0 m 不等的洼地[4],雨季积水成湖,改变区域水系和水循环,导致矿业和农田下泄水直排,极易造成水体富营养化,威胁区域生态和粮食安全[5]。东部地区土地资源稀缺、价值量高,一般采用重构土壤、改善排水、绿肥培土[2]等方式修复受损耕地,期望重建矿区生态、恢复土壤生产力[6]。然而,受地下高潜水季节变动和矸石充填的影响,复垦土壤团聚结构差、pH 值高、肥力低[7],导致生产效益不高,企业、政府和当地农户均缺乏复垦积极性。因此,提升复垦土地生产力,对保护黄淮平原耕地资源至关重要。
黄淮平原矿区缺少土源,一般复垦只能采用“挖深填浅+矸石充填”相结合方式[8]。但受土体构型、潜水淋洗和有害污染的影响,复垦土壤微生物区系发育受限[1],农田生态恢复缺少激发效应和恢复力,导致养分循环不畅、恢复缓慢。微生物是土壤物质循环和养分转化的重要催化剂,在腐植酸形成、有机质分解以及维持土壤生产力、抵抗非生物胁迫等过程中发挥着不可替代的作用[9]。土壤微生物作为土壤最活跃的组分,对外界环境变化十分敏感,并能迅速改变自身结构与组成,适应外界环境的变化[10]。因此,矿区土壤微生物群落结构和功能变化可作为判断矿山复垦土壤质量的指示性指标[4]。先前矿区复垦多关注土壤重构、农田平整、水利配套等工程技术[8,11],对土壤生态功能和微生物资源作用缺乏关注[12-13]。尽管有研究发现复垦能显著增加微生物多样性且与复垦时长呈正相关关系[14-15],但对复垦土壤微生物群落结构和功能缺少长序列监测和评估,导致复垦土壤微生物区系发育不清、功能及其演替的环境驱动机制不明,亟待开展深入研究。
为此,采用高通量测序技术及PICRUSt2 和FUNGuild 分析工具探测山东省邹城市东滩矿区不同复垦年限下土壤微生物群落结构与组成的变化,揭示土壤微生物区系演替规律及环境控制因子,为黄淮平原矿区废弃土地资源科学再利用提供科学依据和技术支撑。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于山东省邹城市东滩矿采煤沉陷复垦区(35°25′52″N、116°52′39″E),属暖温带季风气候,年均气温14.1 ℃,年均降水777.1 mm。该区为典型平原地貌,海拔为(45.2±1.0) m,沉陷前为耕地,土壤类型为棕色潮土,砂粒、粉粒和黏粒质量百分含量分别为223.3 g/kg、658.7 g/kg 和118.0 g/kg,土壤密度1.48 g/cm。2001 年后采用矸石充填(200~400 cm)+表土回填(60~80 cm)复垦工艺+二次机械压实作业方式间隔实施4 次,分别为复垦9 a(编号R9)、复垦12 a(编号R12)、复垦15 a(编号R15)、复垦18 a(编号R18),每次复垦采用工艺和客土来源相同,且复垦为耕地,每个复垦地块面积约2.0 hm2,复垦后耕地由矿业集团发包给农业种植公司统一轮作耕作,种植小麦和大豆,一年两熟。小麦单季施常规复合肥600.0 kg/hm2作底肥,拨节时施氮肥150.0 kg/hm2,机械化收割时留茬高20~25 cm。大豆单季施量常规复合肥450.0 kg/hm2作底肥,机械化收割时自动粉碎还田。并选取附近未受采矿破坏,由农业种植公司统一经营的农田作为对照(CK),整个研究区除复垦时间不同外,成土母质、地形条件、气候水文、种植模式和施肥管理均基本一致。
1.2 样品采集、处理与测试
2020 年8 月22-23 日,采用梅花五点混合采样法收集5 个处理组0~10 cm 表土样约1 000 g,每个处理设13 个重复,总共采集65 个土样。表土作为土壤养分和物质代谢最活跃的部分,是监测重构土壤质量变化的关键。样品采集后剔除石砾、动植物残体等杂质,充分混合后分成2 份。所有样品采用无菌聚乙烯袋密封包装,置于-20 ℃车载冰箱保存,运回实验室后迅速进行后续处理。一份室内风干,磨碎过2 mm 筛,测定土壤基本理化性质和部分土壤酶活性;另一份新鲜土样直接提取微生物总DNA,测定相关微生物信息。其中:pH 采用电位法(水土质量比为2.5∶1)(DDS-307A,上海雷磁),铵态氮(AN)采用氯化钾浸提-紫外分光光度法,硝态氮(NN)采用氯化钙浸提-比色法,有效磷(AP)采用碳酸氢钠浸提-钼锑抗比色法,土壤有机碳(SOC)采用重铬酸钾-比色法[16]。亮氨酸氨基肽酶(LAP)、磷酸酶(PO)、β-葡萄糖苷酶(BG)及蛋白酶(PRO)采用活性试剂盒(索莱宝生物科技,北京)可见分光光度计测定[17]。
1.3 DNA 提取、纯化、PCR 扩增和高通量测序
65 个样品总DNA 提取遵循Fast DNA SPIN kit(MP Biomedicals,美国)试剂盒说明书使用,将提取的总DNA 使用微量紫外分光光度计NanoDrop one 测定浓度和纯度,采用琼脂糖凝胶电泳检测完整性。用引物515F(5′-GTGCCAGCCGGTAA-3′)和907R( CCGTCAATTCMTTRAGTT) 对细菌 16S rRNA 的V4 和V5 区进行PCR 扩增;引物ITS1(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS1(5′-GCTGCGTTCTTCATCGATGC-3′)对真菌进行PCR扩增。PCR 扩增程序如下:① 94 ℃预变性5 min;② 94 ℃变性30 s,52 ℃退火30 s,72 ℃延伸30 s,持续30 个循环;③ 72 ℃最终延伸10 min,降温至4 ℃,实施扩增。扩增结束后用2%琼脂糖凝胶电泳进行PCR 扩增产物检测,用E.Z.N.A.Gel Extraction Kit(Omega,美国)凝胶回收试剂盒对PCR 混合产物进行回收,TE 缓冲液洗脱回收目标DNA 片段。利用Illumina 公司(Illumina,美国)开发的NEBNext® UltraTM II DNA Library Prep Kit 制备测序文库。构建好的文库经Qubit 和Q-PCR 定量确认文库合格后使用Illumina Nova 6000 平台对扩增子文库进行PE250测序(美格基因,深圳)。
1.4 功能预测分析
使用PICRUSt2 比对16S rRNA 测序数据获取的物种信息,推测组成。参考KEGG(https://www.kegg.jp/)数据库,得到KO(KEGG Orthology)功能丰度预测表及KEGG 代谢途径丰度表。采用FUNGuild数据库解析真菌的基因序列信息和生态功能之间关系,借助生态功能群从分类学上划分生态营养型。
1.5 数据统计与处理
利用美格基因云平台计算处理α 多样性指数、β 多样性分析(NMDS)和菌群功能预测等微生物信息分析[18-19]。土壤理化性质和酶活数据采用SPSS 26.0 软件(IBM,美国)进行单因素方差分析(ANOVA)。基于R-project(MathSoft,美国),利用ggplot2 包绘制箱线图,利用vegan 包分析和ggplot2 包绘制NMDS、RDA 图,利用pheatmap 包和ggplot 包完成功能预测分析图。
2 结果与分析
2.1 不同处理组土壤理化性质和酶活性特征变化
不同处理组土壤理化性质和酶活性存在显著性差异(图1)。复垦土壤pH 与CK 呈显著差异(P<0.05),呈先升后降趋势且均高于CK,均表现为弱碱性。除R15 外,R9、R12 和R18 等3 个处理间差异不显著。AN 随复垦年限增加呈上升趋势,除R9 外,其他3 个处理已高于CK,但并无显著差异。NN、AP 和SOC 随复垦年限增加呈下降趋势,复垦处理显著低于CK,R18 分别为CK 的68.5%、26.0%、51.0%。LAP 活性变化趋势与pH 值几乎一致,R15含量最高,约为CK 一倍。PO 活性随复垦年限增加逐渐升高,但仍低于CK 且差异显著(P<0.05)。BG和PRO 活性随复垦年限增加呈下降趋势,R15 时最低,分别下降了19.43%和53.18%。
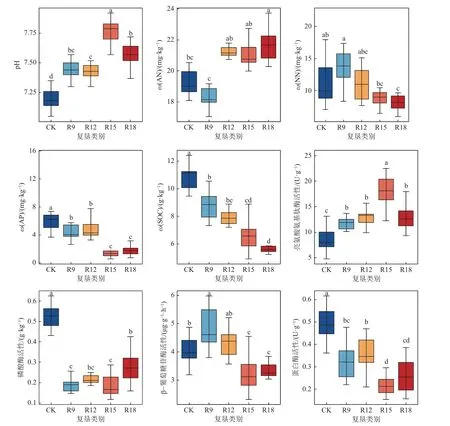
图1 不同处理组土壤理化性质和酶活性变化Fig.1 Changes of soil physico-chemical properties and enzyme activities under different treats
2.2 不同处理组土壤微生物群落结构和组成特征变化
2.2.1 不同处理组土壤微生物群落多样性变化
从65 个样品中共检测到7 335 604 条有效序列,在97%相似度水平上聚类为316 607 个OTUs,不同处理组的有效序列和OTU 数量之间无显著性差异。从表1 可以看出:随复垦年限增加,不同处理组细菌Chao1 指数和Shannon 指数表现为相似趋势,先增加后减少,R12 时最高。Pielou 均匀度指数和Simpson指数表现为相反的变化趋势,R12 时Pielou 均匀度指数最高,Simpson 指数则最低,且与CK 呈显著差异(P<0.05)。真菌群落R9 和R12 处理Chao1 指数低于CK,Simpson 指数高于CK,但R15 和R18 处理Chao1 指数已高于CK,Simpson 指数低于CK。所有复垦处理的Shannon 指数和Simpson 指数均高于CK。所有复垦处理的Shannon 指数和Simpson指数均高于CK,表明复垦显著地促进真菌物种相对丰度的变化,R12 时真菌物种丰富度和多样性最高。
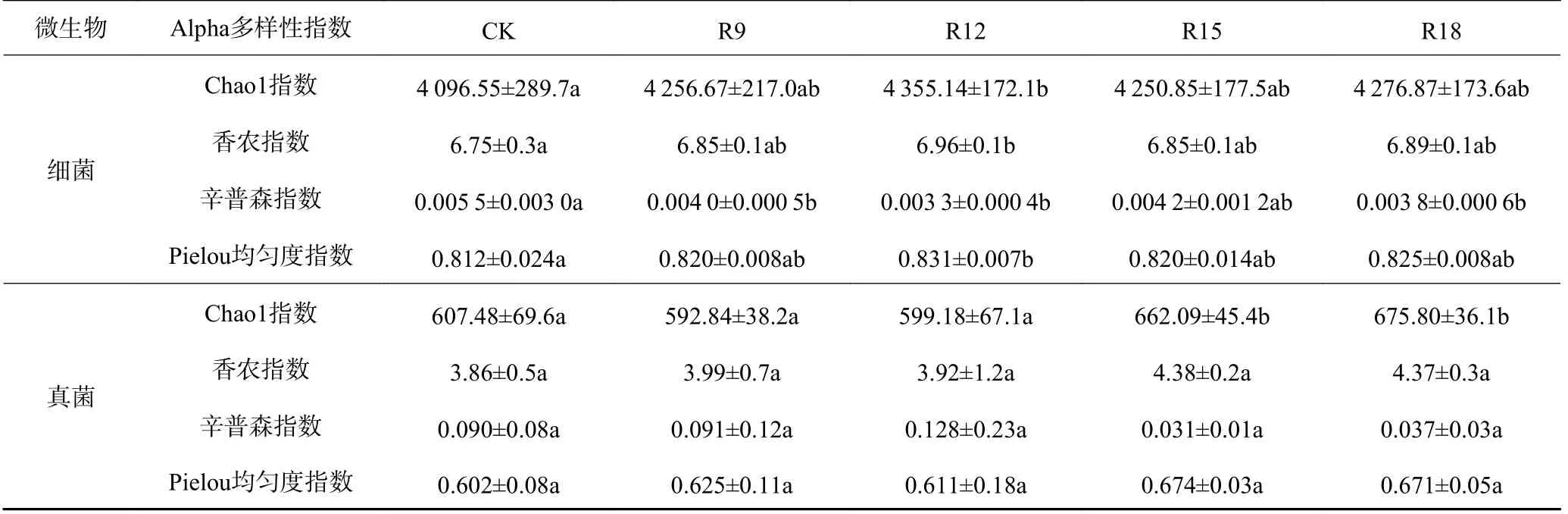
表1 不同处理组土壤微生物群落Alpha 多样性Table 1 Alpha diversity of soil microbial community under different treats
从图2 可知:不同处理细菌Beta 多样性组间分离、组内紧密,形成明显的聚类(图2a)。其中,横坐标(NMDS1)和纵坐标(NMDS2)为样本间差异解释度最大的两个坐标。利用胁强系数(stress 值)衡量NMDS 分析结果,检验结果是否能反映数据排序的真实分布。图2 中stress 值均小于0.2,由此表明分析结果具有一定的解释意义。所有复垦处理沿x轴与CK 分离,表明复垦与CK 之间群落结构存在显著差异。R9 和R12 沿y轴与R15 和R18 之间呈分离趋势,R9 与R18 不重叠,R15 与R18 差异不明显。但R18 聚类优于其他组,充分表明复垦组间差异和相似度。不同处理真菌群落NMDS 分析显示复垦处理沿x轴与CK 之间存在距离(图2b),表明复垦与CK 之间群落存在明显的分异。R9 和R12 与R15和R18 之间在y轴上产生距离,但两两之间系统发育更接近,表明不同复垦年限及CK 之间群落Beta多样性差异显著。
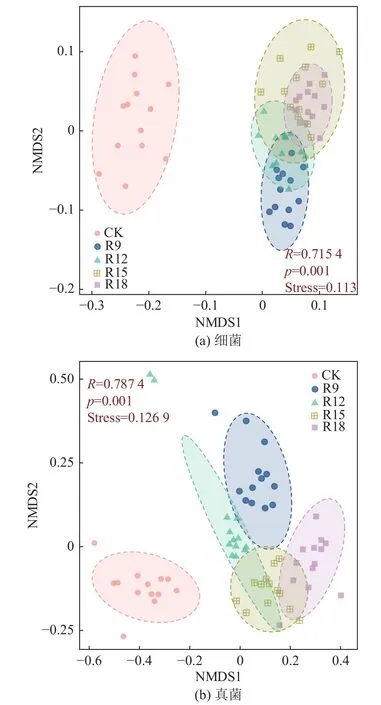
图2 不同处理组土壤细菌、真菌Beta 多样性Fig.2 Beta diversity analysis of soil bacteria and fungi under different treats
2.2.2 不同处理组土壤微生物群落组成变化
所有处理土壤细菌群落主要菌门保持一致(图3a),变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、拟杆菌门(Bcateroidetes)、放线菌门(Actinobacteria)和绿弯菌门(Chloroflexi)为优势菌门,相对丰度占比大于80.0%。其中,变形菌门最丰富,相对丰度为32.5%±3.3%,表明它在维持矿区复垦土壤复杂生态环境中起着重要作用。尽管群落中主要菌门相似,但相对丰度变化明显。复垦处理组变形菌门的相对丰度显著低于CK(P<0.05),但酸杆菌门则相反。随复垦年限增加,变形菌门和放线菌门的相对丰度逐渐上升,酸杆菌门和拟杆菌门则逐渐下降,并均呈向CK 靠近的趋势。所有处理组中子囊菌门(Ascomycota)、被孢霉门(Mortierellomycota)及担子菌门(Basidiomycota)为真菌群落主导菌门(图3b),相对丰度分别为28.9%±2.5%、16.5%±6.7%和9.7%±6.8%,占比超过一半。子囊菌门随复垦年限增加呈上升趋势,与CK 相比,R15 和R18 中子囊菌门分别上升了115.0%和127.02%。总体来看,随复垦年限增加,细菌菌门类型变化更为复杂,真菌菌门丰度变化更加显著。
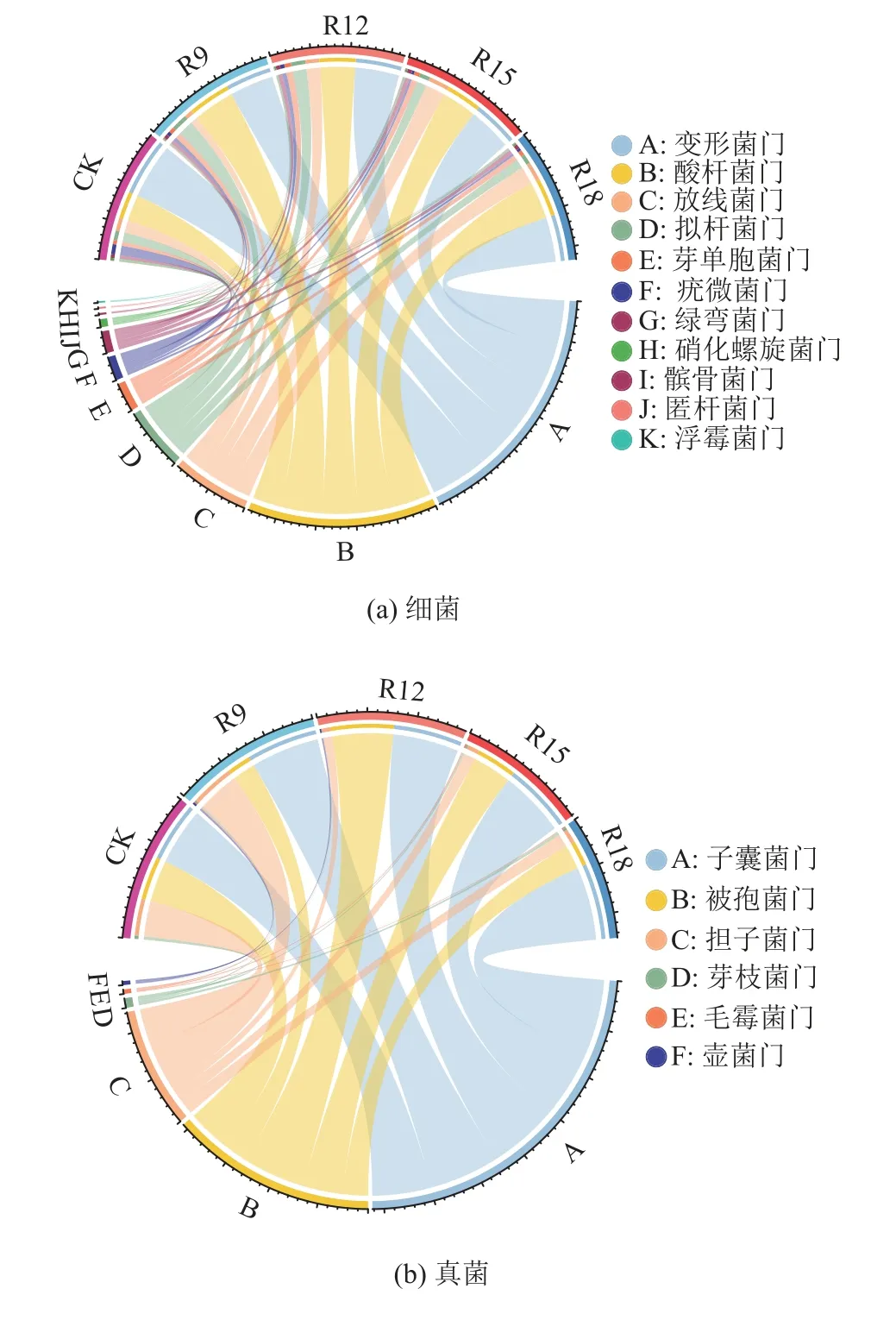
图3 不同处理组土壤细菌、真菌主导菌门相对丰度Fig.3 Relative abundance of soil bacteria and fungi under different treats at phylum level
2.3 微生物群落与环境因子间的相互作用
利用R 软件vegan 包计算OTUs 水平除趋势对应分析的lengths of gradient(LG),细菌和真菌OTUs 水平LG 值分别为1.218 7 和3.861 2,均小于4.0,可采用冗余度分析(RDA)。通过RDA1 和RDA2 表示环境因子与排序轴之间的相关性。图4 显示环境因子分别解释了细菌和真菌群落结构77.3%和56.5%的总变异(细菌RDA1 和RDA2 的解释率分别为58.92%和18.35%,真菌RDA1 和RDA2 的解释率分别为29.08%和27.43%),RDA 前两轴已较好地解释了土壤微生物与环境因子之间的关系。RDA分析结果显示,不同处理间差异明显,复垦组与CK分别形成明显的聚类,表明复垦对土壤微生物群落结构作用显著,且受不同复垦年限土壤理化性质变化影响。pH、AN 和PO 为土壤微生物群落结构变化的主导因子,复垦处理土壤微生物群落结构与pH、AN 和LAP 表现为正相关关系,与PRO、SOC 和AP表现为负相关关系。
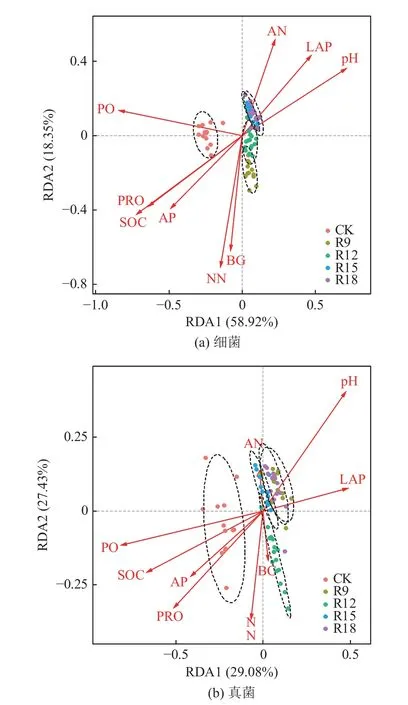
图4 OTUs 水平土壤细菌、真菌和环境因子间RDA 分析Fig.4 RDA analysis of soil bacteria and fungi and environmental factors at OTU levels
2.4 不同处理组土壤微生物群落功能预测分析
为预测不同处理组土壤细菌功能变化,基于KEGG 数据库,采用PICRUSt2 进行功能预测分析。结果显示:不同处理组细菌群落表现为相似的基因功能,包含3 个一级、15 个二级及50 个三级功能层(图5a)。三级功能层主要包含代谢、复制和修复、翻译及细胞生长与死亡等功能。复垦处理在丙酮酸代谢、糖酵解/糖异生、丙酸代谢、丁酸代谢等18 种功能上与CK 存在差异,它们随复垦年限增加不断增强。但叶酸一碳库、叶酸生物合成、生物素代谢等10 种功能则随复垦年限增加有所减弱。
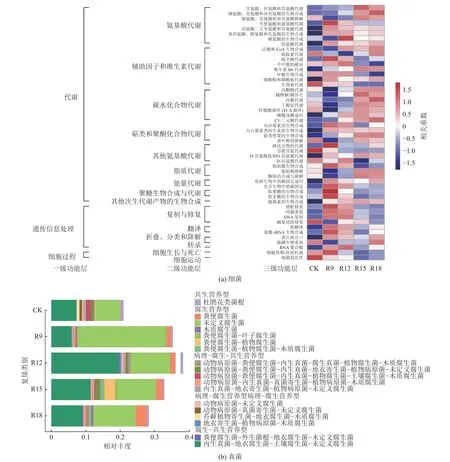
图5 不同处理组微生物功能基因预测相对丰度Fig.5 Map of relative abundance of microbial predicted functional genes under different treats
采用FUNGuild 对土壤真菌进行功能预测分析(图5b),结果显示:复垦土壤真菌群落中包含2 个单营养型(腐生营养型、共生营养型)和3 个混合营养型(病理营养型-腐生营养型-共生营养型、病理营养型-腐生营养型、腐生营养型-共生营养型)。此外,包括4 个单一功能群和14 个混合功能群。不同处理组真菌群落以未定义的腐生真菌为主,其次是内生菌-垃圾腐植生物-土壤腐植生物-未定义的腐植生物和粪便腐生生物,不同处理组功能差异显著。
3 讨 论
3.1 复垦年限对土壤理化性质和酶活性的影响
pH 是土壤微生物多样性和丰富度最主要的控制因素,影响养分有效性[20]。本研究发现随复垦年限增加,pH 先升后降。这可能与季节性潜水变动有关,初期下层充填煤矸石中部分重金属和盐离子溢出,发生毛细现象被带至表土层,这与先前研究的结果一致[3]。但随复垦年限延长,土壤中一部分养分受到淋溶作用向下迁移,并与重金属和盐离子发生化学反应,导致后期土壤pH 下降。土壤中重金属累积对土壤微生物群落结构、组成乃至功能产生了重要影响,导致微生物群落结构与pH 间呈显著相关[21]。土壤SOC 降低与作物吸收、土壤动物和微生物分解消耗相关[11],随复垦年限增加,微生物介导的土壤呼吸作用不断增加,不利于SOC 的积累[22]。这与CHAUDHURI 等[23]的研究结果不同,但与张宇婕等[11]的研究成果相一致,主要可能与气候、植被、成土母质的差异有关。土壤有效磷是可被植物吸收的磷组分,有研究发现AP 随复垦年限增加逐渐降低[24],这与本研究结果完全一致,可能主要归结于复垦年限增加,土壤质量得以改进,作物生产力得以提升,需要吸收更多磷素,以满足其生长需要。且高潜水位地区降雨量变化致使的土壤中离子和矿物含量变化,亦是能够影响土壤磷元素的化学形态。AN 随复垦年限呈上升趋势,主要原因为随复垦年限增加,作物根系和凋落物等进入土壤越多,酶活性提高,加速养分释放[13],这也是SOC 持续下降的原因之一。与此同时,有机肥和化肥输入也越多,外源氮素逐渐累积。此外,随复垦年限增加,有机氮矿化作用增强,硝化作用减弱,导致AN 上升,NN 下降[22]。微生物利用分泌胞外酶将复杂化合物降解为水溶性分子供自身或作物利用[25],因此酶活性变化反映了复垦对土壤生态环境的影响。如LAP 参与土壤氮循环[26],本研究显示LAP 先升后降,与豆科植物早期的固氮作用有关,增加氮可利用性,酶活性增强。后期AN 不断积累又抑制LAP 活性,这与先前发现LAP 与AN 之间存在负反馈调控相一致[27]。PO 活性逐渐升高可能与复垦增加酶作用的底物从而间接提高酶活性,也可能与AP 含量下降有关,呈一定的负反馈调节[28]。BG 参与多糖分解,PRO 参与有机含氮化合物水解,两者酶活性下降与复垦土壤耕作扰动和SOC 下降有关,反映土壤微观环境质量状况[29]。
3.2 不同处理对微生物群落结构与组成的影响
本研究显示:随复垦年限增加,微生物群落丰富度和均匀度也上升(图2)。主要由于复垦消除了采矿对土壤环境的胁迫,农业生产和施肥又为微生物提供充足养分和碳源,激发特定功能菌类群发育,这与马静等[21]的研究结论相一致。有研究发现,随复垦年限增加,根系和凋落物源源不断输入,提高了土壤肥力,改善了理化性质[30],为微生物群落创造了适宜的生存条件,促进微生物恢复至原始或更高水平[31]。本研究NMDS 分析结果显示,CK 在y轴上与R18 处理组样点更接近,说明随复垦时间增加,微生物群落组成差异逐渐消失[9]。与此同时,本研究发现细菌群落多样性远高于真菌,这与李智兰[1]、周雪岩[32]等的研究结论一致。变形菌门和放线菌门相对丰度随复垦年限增加而增加(图3a),主要原因是黄淮平原矿区复垦土壤为弱碱性,它们对碱性土壤亲和力高。拟杆菌门和酸杆菌门相对丰度随复垦年限增加而下降(图3a),说明复垦对其生存带来了挑战,改变了环境营养状况和碳源利用潜力,尤其是属寡营养型的酸杆菌门[33]。子囊菌门和被孢霉门为复垦土壤真菌的主导菌门(图3b),子囊菌门相对丰度随复垦年限明显增加,说明复垦改善了环境胁迫,为偏爱高营养底物的腐生菌提供更适宜的生存环境[34]。
3.3 土壤微生物群落与环境因子的互作机制
复垦改变了地上植物群落和地下土壤环境,不可避免地引发微生物群落结构和功能的变化[35]。因此,微生物群落结构与土壤理化性质之间存在一定的相关关系[36]。本研究发现,复垦处理组pH、AN和LAP 与细菌和真菌群落结构表现为显著正相关关系,PRO、SOC 和AP 与细菌和真菌群落结构表现为显著负相关关系。LAP 促进有害物降解,PRO 增强微生物适应能力,SOC 分解提供更多的营养物质和能量,有效地提升了土壤微生物群落与环境因子间互馈机制。随复垦年限增加,微生物区系不断发育,功能逐渐完善。复垦土壤细菌形成了3 个一级功能层、15 个二级功能层和50 个三级功能层(图5a)。代谢通径为核心功能,对土壤-植被的碳氮磷循环至关重要[37-38],与复垦土壤特定环境胁迫防御、固氮、溶磷等互作密切相关。二级功能主要包含氨基酸代谢、碳水化合物代谢及辅助因子和维生素代谢等,为存活与繁衍服务[39]。复垦土壤中放线菌门拥有较多的促进动植物遗骸腐烂的好氧性腐生菌,变形菌门中存在许多代谢功能类群,为植物生长和自身发育提供物质和能量[40]。真菌生活史更为复杂,FUNGuild分析结果显示腐生营养型真菌在复垦土壤中最具优势(图5b)。它们可以腐生、寄生和共生等多种营养方式生存,既可分解动植物残体,又可腐生植物根茎叶等不同部位[41]。腐生菌多属子囊菌门,因此子囊菌门成为复垦土壤真菌最优势菌门。R9 中未定义的腐生真菌相对丰度值最高,说明复垦初期养分胁迫严重,腐生真菌分解有机物对植被生长和满足自身利用至关重要。土壤真菌的分解能力明显增加,有利于作物吸收土壤中的营养物质,促进植被的生长发育。综上,土地复垦能有效提高黄淮平原矿区土壤微生物活性,促进养分吸收和物质代谢循环(图6)。今后黄淮平原采煤沉陷区复垦应加强工艺控制,一是防止过分压实,减少毛细现象发生;二是适当增大降渍深度,防止充填煤矸石等重金属和盐离子上升,产生生物毒性;三是可适当添加功能微生物,加速微生物群落重建,快速改良重构土壤质量。
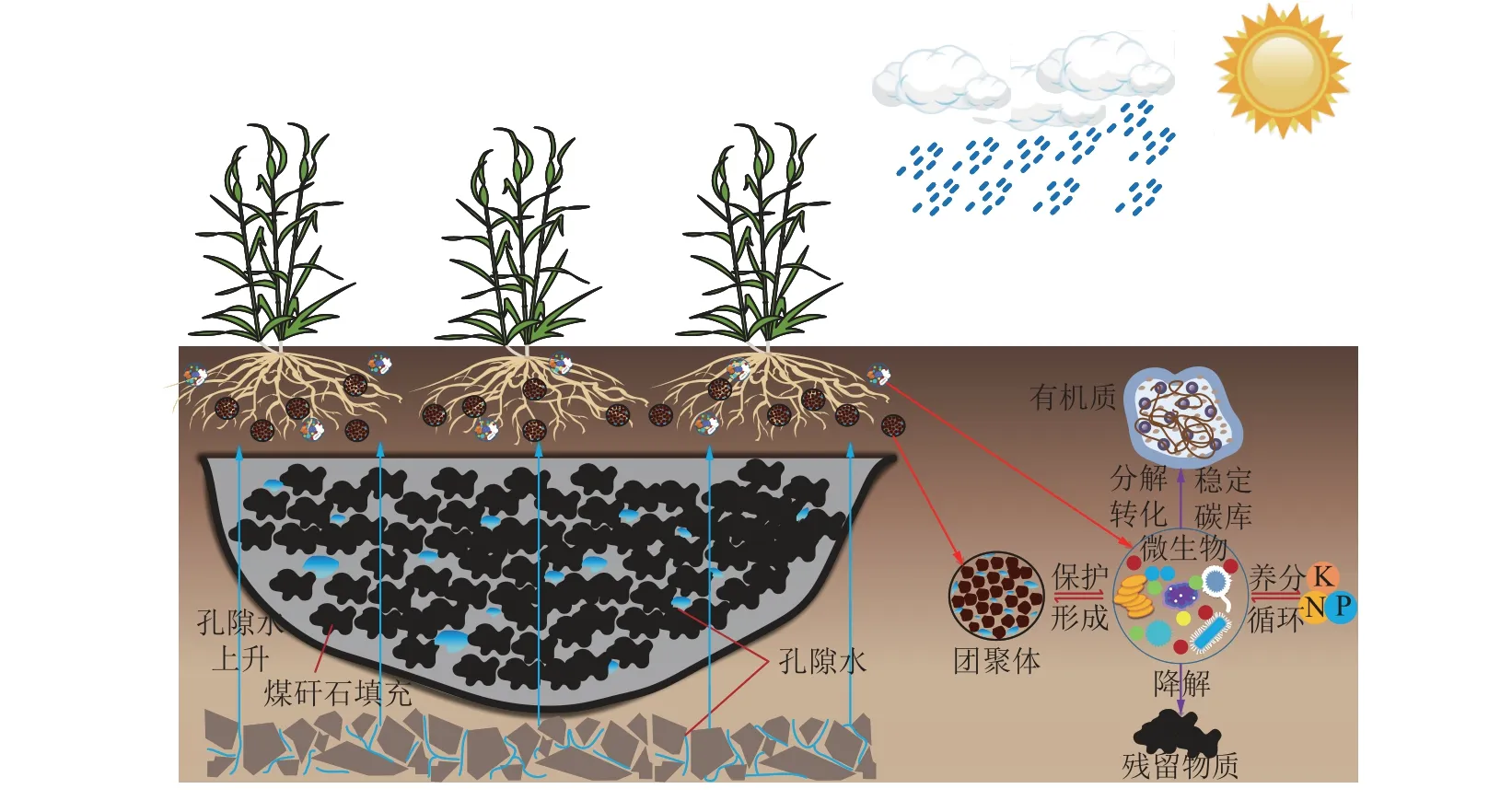
图6 微生物在高潜水位矿区复垦土壤中的作用过程和机制Fig.6 The microbial effecting process and mechanism in the reclaimed soil of high underground water mining areas
4 结 论
1)黄淮平原矿区复垦显著地改善土壤理化性质和酶活性。随复垦年限增加,AN 逐渐上升,NN、AP、SOC、BG 和PRO 活性下降,pH 和LAP 活性呈先升后降趋势。
2)复垦对土壤微生物群落结构与组成有显著影响。随复垦年限增加,细菌Chao1 指数和Shannon指数先增加后减少,Pielou 均匀度指数和Simpson 指数则相反。真菌群落Shannon 指数和Simpson 指数高于CK,但R9 和R12 处理Chao1 指数低于CK,Simpson 指数高于CK。所有处理土壤细菌和真菌群落的主要菌门保持一致,但相对丰度变化显著。
3)土壤微生物群落与环境因子之间存在特定的相关关系。土壤pH、AN 和PO 为微生物群落结构变化的主导因子,复垦土壤pH、AN、LAP 及PRO、SOC、AP 与微生物群落结构分别成正相关与负相关关系。
4)PICRUSt2 功能预测结果显示不同处理组细菌群落基因功能相似,包含3 个一级、15 个二级及50 个三级功能层,其中代谢功能活跃且与复垦年限呈正相关关系。FUNGuild 功能预测结果显示不同处理组真菌群落包含2 个单营养型和3 种混合营养型及4 个单一功能群和14 个混合功能群,腐生营养型最具有优势。
猜你喜欢
南京林业大学学报(自然科学版)(2022年4期)2022-11-29 11:51:32
淡水渔业(2022年3期)2022-05-25 07:16:04
食品安全导刊(2021年32期)2021-11-27 15:31:06
食药用菌(2021年5期)2021-10-15 06:23:12
食品科学(2020年23期)2020-12-31 01:31:42
林业科学(2020年10期)2020-11-30 00:36:56
中国比较医学杂志(2020年4期)2020-05-26 05:47:22
科学导报(2019年28期)2019-09-03 04:34:03
水生生物学报(2019年4期)2019-07-20 08:08:10
生物安全学报(2019年3期)2019-02-15 16:54:12
