
miR-335-5p过表达对胃癌基因表达谱的影响及筛选基因的验证研究*
2023-12-21 05:31王宏伟赵雪灵高夏青陈凤琴李海龙
国际检验医学杂志 2023年23期
王宏伟,赵雪灵,王 蒙,高夏青,陈凤琴,宁 月,李海龙△
1.甘肃中医药大学附属医院检验科,甘肃兰州 730020;2.甘肃中医药大学第一临床医学院,甘肃兰州 730101;3.甘肃中医药大学附属医院功能科,甘肃兰州 730020
微小RNA(miR)-335位于染色体7q32.2,在胃癌的发生、发展中发挥了重要作用[1]。有报道指出,miR-335-5p在胃癌组织中表达下调,与其启动子甲基化有关[2]。miR-335-5p抑制胃癌细胞的增殖和迁移,并诱导细胞凋亡,把胃癌细胞周期阻滞在G1/S期[1]。临床病理分析显示,miR-335低表达可能与胃癌发生发展有关,可一定程度评估患者预后[3]。为深入探明其调控胃癌细胞生物学行为的机制,本研究构建了miR-335-5p过表达慢病毒载体并感染胃癌细胞,然后选择人类全基因组表达谱芯片筛选过表达miR-335-5p的胃癌细胞与对照细胞之间差异表达的mRNA,并采用实时荧光定量PCR(qRT-PCR)和蛋白质印迹法(Western blot)验证。
1 材料与方法
1.1材料来源 胃癌细胞MGC-803购自中科院上海细胞库。靶基因环腺苷酸反应元件结合蛋白1(CREB1)、苏氨酸激酶3(AKT3)、外被体包被蛋白β2亚基(COPB2)、芳香烃受体核易位蛋白2(ARNT2)、转化生长因子(TGF)-β2、Kirsten大鼠肉瘤病毒癌基因(KRAS)、血小板源性生长因子-β(PDGF-β)、GAPDH、辣根过氧化物酶标记的山羊抗兔IgG(二抗)购自Immunoway生物科技有限公司。
1.2仪器与试剂 电泳和湿转仪(美国Bio-rad公司、DYCZ-24D),酶标仪(美国Molecular Devices公司、SpectraMax i3X),凝胶成像仪(广州博鹭腾生物科技有限公司、GelView 6300Plus),PCR扩增仪(美国ABI公司、ABI7500)。BCA试剂、配胶试剂购自北京索莱宝科技有限公司;增强型ECL发光试剂购自上海熠圣生物科技股份有限公司;miR-335-5p的过表达和阴性对照慢病毒载体由上海吉凯基因技术有限公司采用GV369载体合成,克隆位点为BamHNheⅠ;miDETECTA Track miRNA qRT-PCR starter kit购自广州锐博生物有限公司;逆转录试剂盒购自上海翊圣生物科技有限公司;高糖DMEM培养基(美国Hyclone公司),胎牛血清(杭州四季青公司),青链霉素双抗(北京索莱宝科技有限公司)。
1.3方法
1.3.1细胞培养 在含10%胎牛血清的高糖DMEM培养基中加入青霉素(100 U/mL)、链霉素(100 U/mL),在37 ℃、5%CO2的细胞培养箱中常规培养。每隔2~3 d换完全培养基,待细胞密度达90%左右时按1∶2比例传代。取对数生长期的细胞用于下一步实验。
1.3.2慢病毒载体感染胃癌细胞 培养胃癌MGC-803细胞至对数生长期,消化细胞后用含有10%FBS的高糖DMEM培养基重悬细胞后计数,调成单细胞悬液,接种在6孔板中,置于37 ℃培养箱中备用。待过夜后的细胞生长至40%~50%时,用感染增强液稀释配备1×108TU/mL的病毒液。取10 mg/mL polybrene以DMEM培养基稀释至终浓度为5 μg/mL(指1 mL感染时体积),备用。在6孔板中采用1×108TU/ mL的病毒液感染胃癌细胞,将培养体系混匀后继续培养8~12 h后,观察细胞状态,并更换为新鲜的培养基。感染72 h后,观察荧光表达情况并拍照,消化细胞接种至细胞培养瓶继续培养。待细胞生长至70%~80%时,消化传代培养,收集部分细胞,提取RNA,逆转录后,扩增并计算miR-335-5p水平,以确定感染后细胞内miR-335-5p水平。
1.3.3MTT法检测过表达miR-335-5p对胃癌细胞增殖的影响 将稳转miR-335-5p和阴性对照的MGC-803细胞培养至对数生长期,收集细胞后用不含有胎牛血清的高糖DMEM培养基重悬细胞后计数,调成单细胞悬液,以200 μL体积加入96孔板,使细胞数为5 000个/每孔,置于37 ℃培养箱过夜备用。待24、48、72、96 h后每组每孔加入MTT 20 μL,继续孵育4 h,吸去上清液,每孔加入二甲基亚砜150 μL,振荡10 min,用酶标仪以570 nm波长检测各孔吸光度(A)。按A值计算各组的平均值,实验重复3次。
1.3.4Transwell小室检测过表达miR-335-5p对胃癌细胞的迁移能力的影响 将成功转染miR-335-5p的胃癌细胞用不含血清的培养液制成5×104/mL的悬液,取200 μL加入Transwell上室中,在下室中加入500 μL含有10%胎牛血清的培养基。细胞在37 ℃下培养48 h。将小室内的培养基丢弃,加适量预冷磷酸盐缓冲液(PBS)将细胞洗涤2次,添加足够的4%细胞固定液将小室下侧细胞固定2 min,将固定液倒掉并用PBS洗涤2次,随后用100%甲醇固定,25 min后将甲醇倒掉并用适量的PBS将细胞洗涤2次,加0.1%的结晶紫在避光条件下进行20 min染色,结晶紫染液倒掉并用足量PBS洗涤细胞2次,随后用棉签小心将小室内侧细胞拭去,常温下放置30 min,待其风干后于光学显微镜下观察拍照。随机选取3个视野进行拍照,以穿过小室细胞数代表细胞数量的迁移能力。
1.3.5表达谱芯片筛选过表达miR-335-5p后胃癌细胞差异表达的基因 将稳转miR-335-5p和阴性对照细胞培养至对数生长期,收集细胞后提取总RNA,采用Agilent mRNA表达谱芯片杂交分析,使用晶芯®cRNA扩增标记试剂盒进行样品RNA的荧光标记,逆转录合成1st-strand cDNA和2nd-strand cDNA。基因芯片杂交反应在42 ℃温度下16~20 h完成,洗涤后,应用芯片扫描仪进行扫描。杂交芯片在扫描后,使用Agilent G2565CA Microarray Scanner软件扫描图像(TIFF格式),扫描后的TIFF格式图片数据采用Feature Extraction提数软件进行预处理分析,然后采用GeneSpring GX软件,写入分组等参数信息。进行每个样品的数据归一化和QC分析,然后计算基因表达差异和P值。
1.3.6miRNA靶基因预测和靶基因富集信号通路分析 使用miRSystem网站(http://mirsystem.cgm.ntu.edu.tw/index.php)中集成的DIANA、miRanda、miRDB、miRWalk、PICTAR、PITA、RNA22、Targetscan软件对miR-335-5p的靶基因进行预测,采用京都基因与基因组百科全书进行信号通路富集分析,选取至少3个软件支持的靶基因作为下一步分析的候选基因。并与1.3.5所述表达谱芯片筛选基因取得交集基因,然后将交集基因开展靶基因富集信号通路分析。
1.3.7筛选基因PCR扩增检测 采用Trizol试剂从细胞中提取总RNA,检测RNA浓度和纯度(A260/A280在1.8~2.0),以1%琼脂糖凝胶电泳鉴定其完整性(28 S和18 S RNA条带比值≥2.0)。提取完成后,进行逆转录反应,反应条件如下:37 ℃ 15 min,85 ℃ 5 s。然后进行qRT-PCR,反应条件:预变性95 ℃ 10 s,变性95 ℃ 15 s,退火60 ℃ 20 s,40个循环。扩增反应结束后进行溶解曲线分析,判断产物是否有非特异性扩增;分析扩增曲线,计算Ct值,以β-actin作为内参基因,通过2-ΔΔCt法计算各组间mRNA水平差异(模型组基因水平设为1)。实验重复3次,引物合成由大连宝生物工程有限公司合成,引物序列见表1。

表1 qRT-PCR引物
1.3.8Western blot 收集各组处理的细胞,提取总蛋白,加入蛋白裂解液(RIPA∶PMSF为100∶1)提取总蛋白,将蛋白煮沸变性后,以十二烷基硫酸钠-聚丙烯酰胺凝胶电泳将蛋白分离,湿转至聚偏氟乙烯膜上,以5%脱脂奶粉封闭1.5 h,一抗孵育过夜(CREB1、AKT3、COPB2、ARNT2、TGF-β2、KRAS、PDGFB、GAPDH 1∶500~1∶1 000稀释),TBST洗膜3次,二抗孵育2 h(羊抗兔,1∶2 000稀释),TBST洗膜3次,加入ECL于Bio-rad凝胶成像仪中进行反应,实验重复3~5次,采用Image J软件对Western blot结果进行定量分析,灰度值以累积吸光度值表示,结果以目的蛋白与GAPDH的累积吸光度的比值表示。
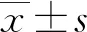
2 结 果
2.1慢病毒感染效果的观察及检测 慢病毒miR-335-5p感染MGC-803细胞72 h后在倒置荧光显微镜镜下观察,结果显示绿色荧光细胞占全视野细胞在80%以上,见图1。qRT-PCR结果显示,慢病毒miR-335-5p感染MGC-803细胞72 h后,MGC-803细胞中miR-335-5p水平(130.74±2.67)较阴性对照组(1.00±0.11)明显升高(P<0.05),见图2。
注:A为明场下细胞(×100);B为暗场下细胞(×100)。

图2 各组miR-335-5p水平比较
2.2过表达miR-335-5p对胃癌细胞增殖的影响 MTT结果显示,与阴性对照组比较,miR-335-5p组细胞的48、72、96 h 的A值均明显降低,差异有统计学意义(P<0.05),表明miR-335-5p可降低胃癌细胞活力、抑制其增殖,见图3。

注:与同时段阴性对照组比较,aP<0.05。
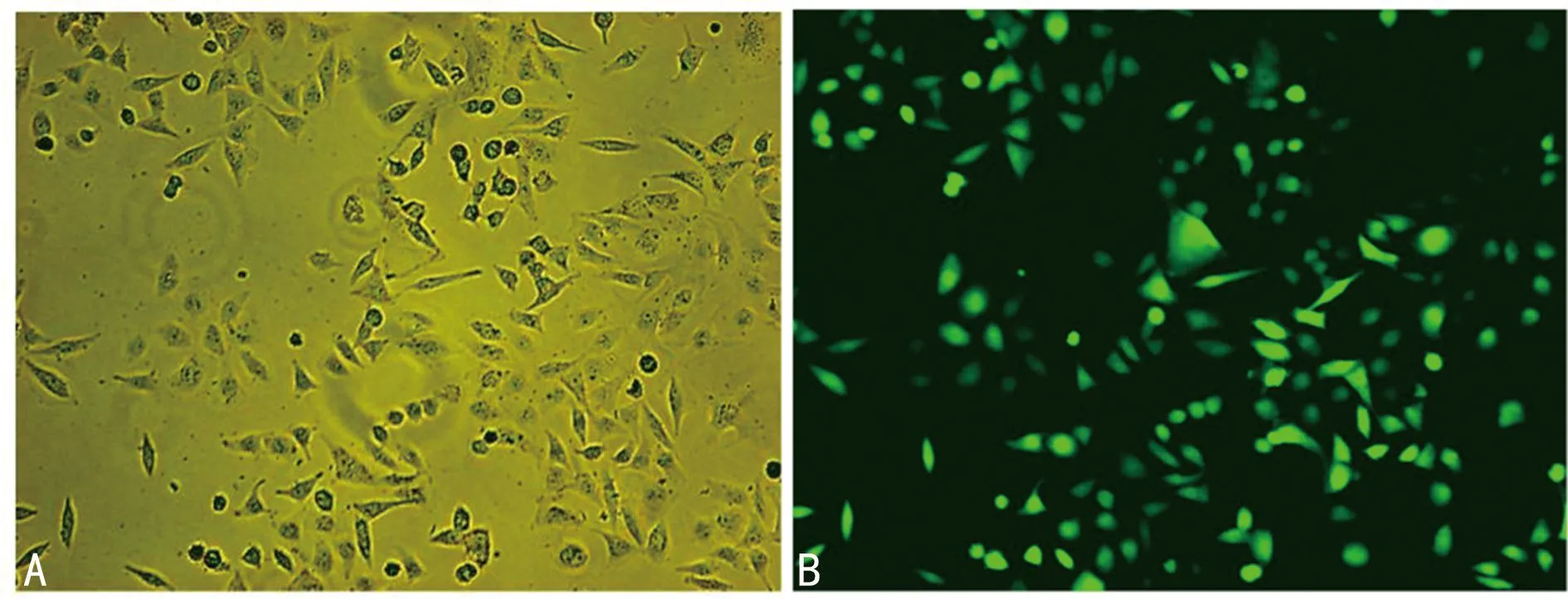
2.3过表达 miR-335-5p 对胃癌细胞侵袭能力的影响 Transwell结果显示,与阴性对照组比较,miR-335-5p组细胞数明显减少,差异均有统计学意义(P<0.05),表明过表达miR-335-5p可抑制胃癌细胞的侵袭能力。见图4。
注:A为阴性对照组;B为miR-335-5p组。
2.4表达谱芯片筛选差异基因和富集信号通路分析 表达谱芯片研究发现,miR-335-5p稳转细胞和阴性对照细胞之间差异表达的基因(FC大于2.0倍)有1 713个。生物信息学研究表明,经DIANA、miRanda、miRDB、miRWalk、RNAhybrid、PICTAR4、PICTAR5、PITA、RNA22和Targetscan软件共同预测miR-335-5p的靶基因,其中至少3个软件支持的靶基因有3 013个,将所得基因与表达谱芯片筛选的差异基因取交集,获得交集基因212个,并将获得的交集基因运用DAVID 6.8软件进行富集分析,共富集到20条信号通路,其中绝大部分为与肿瘤生物学有关的信号通路,如TGF-β通路、mTOR 通路、凋亡通路、p53信号通路等均为调控肿瘤生物学表型的重要通路。见图5。

图5 富集在与肿瘤密切相关的信号通路中的交集基因
2.5靶基因CREB1、AKT3、COPB2、ARNT2、TGF-β2、KRAS、PDGF-β的mRNA表达情况 为验证感染慢病毒miR-335-5p和阴性对照的MGC-803后胃癌细胞中筛选靶基因在蛋白水平上的表达情况,选择表达谱芯片筛选后差异表达的核心基因进行验证。qRT-PCR实验验证结果表明,相对于阴性对照组,相关靶基因CREB1(0.53±0.08)、AKT3(0.60±0.05)、COPB2(0.43±0.05)、ARNT2(0.57±0.07)、TGF-β2(0.73±0.09)、KRAS(0.14±0.03)、PDGF-β(0.42±0.07)的mRNA水平降低,差异有统计学意义(P<0.05)。见图6。
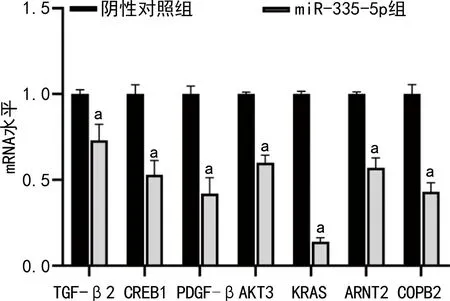
注:与阴性对照组比较,aP<0.05。
2.6靶基因CREB1、AKT3、COPB2、ARNT2、TGF-β2、KRAS、PDGF-β的蛋白表达情况 Western blot结果表明,与阴性对照组比较,相关靶基因CREB1(0.20±0.01)、COPB2(0.40±0.02)、AKT3(0.30±0.01)、ARNT2(0.49±0.01)、TGF-β2(0.21±0.00)、KRAS(0.19±0.01)、PDGF-β(0.21±0.01)的蛋白水平明显降低(P<0.05)。见图7。
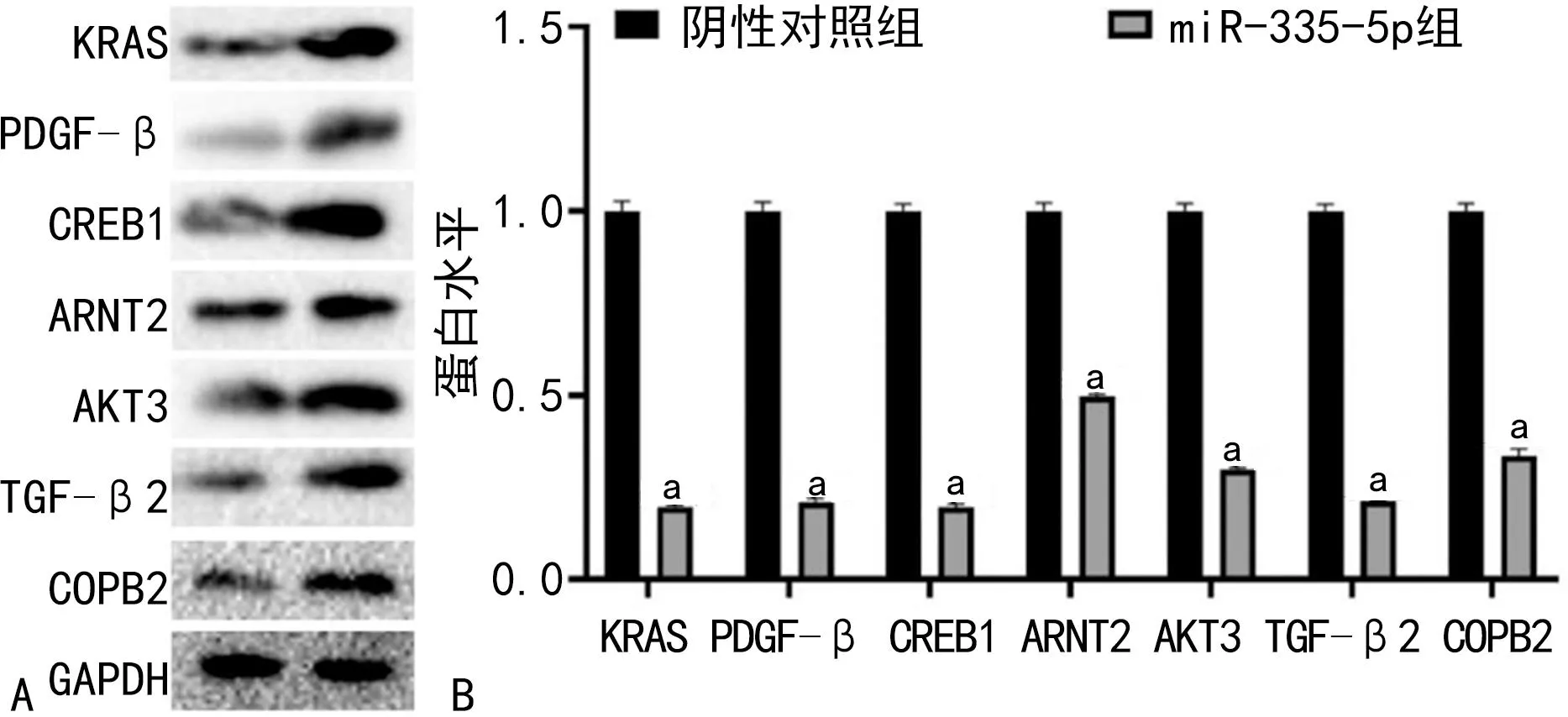
注:A为Western blot蛋白效果图;B为Western blot蛋白定量图;与阴性对照组比较,aP<0.05。
3 讨 论
胃癌是全球高发的恶性肿瘤之一,也是全球常见癌症相关死亡原因中排第2位的恶性肿瘤。目前胃癌治疗主要通过手术切除,但绝大多数胃癌在确诊时已属于进展期,已失去手术根治机会。因此,寻找早期诊断标志物是肿瘤研究的重大方向。目前,miRNA已成为胃癌诊断中的重要标志物。为进一步研究其生物学表型和功能意义,本研究通过慢病毒miR-335-5p过表达载体感染胃癌细胞的方式在胃癌细胞中上调其表达,从而观察对胃癌细胞生物学增殖、侵袭方面的影响,并通过表达谱芯片与预测靶基因取交集的方式研究部分靶基因和富集的信号通路。
本研究结果表明,所筛选的靶基因所富集的信号通路如TGF-β信号通路、mTOR信号通路、凋亡通路、p53信号通路等均为调控肿瘤生物学表型的重要通路。相关研究发现,以上通路均是与胃癌发生、发展密切相关的通路,这些通路的改变为深入探讨miR-335-5p的功能提供了依据。Western blot验证结果表明,过表达miR-335-5p可以下调胃癌细胞中CREB1、AKT3、COPB2、ARNT2、TGF-β2、KRAS、PDGF-β的表达。
AKT3是一种与细胞周期有关的丝氨酸/苏氨酸蛋白激酶。AKT激酶是响应胰岛素和生长因子的细胞信号的调节器,它们参与多种生物学过程,包括细胞增殖、分化、凋亡、肿瘤发生及糖原合成和葡萄糖摄取。该激酶已被证实可被PDGF、胰岛素和胰岛素样生长因子1(IGF1)刺激。HU等[4]研究表明,AKT3在胃癌组织中高表达。LIU等[5]研究发现,原发性胃炎是癌症的一种独特表型,生存率低,3个PI3K/AKT通路相关基因(PIK3R2、AKT3和IGF1)在原发性胃炎中表达上调。TAKAHASHI等[6]发现,AKT3表达是预测头颈部鳞状细胞癌免疫反应性和预后的潜在生物标志物。此外,AKT3的异构体特异性抑制可作为一种新型的癌症治疗方法,有效抑制PI3K/AKT/mTOR通路[5-6]。
ARNT2是碱性螺旋-环-螺旋-PAS转录因子超家族的重要成员之一。编码的蛋白作为碱性螺旋-环-螺旋-PAS家族的几个传感器蛋白的伴侣,与传感器蛋白形成异二聚体,该异二聚体结合响应发育和环境刺激的基因中的调节性DNA序列。在缺氧条件下,编码的蛋白与细胞核中的缺氧诱导因子-1α组成复合物,该复合物与氧反应基因增强子和启动子中的缺氧反应元件结合。小鼠中高度相似的蛋白质与芳基烃受体和单一蛋白质形成功能复合物,表明编码的蛋白质在异源化合物的代谢和神经发生的调节中具有额外的作用。JIA等[7]研究发现,ARNT2在胃癌组织中低表达,且与肿瘤的浸润深度、分化程度和较差的生存率呈负相关。过表达ARNT2可抑制细胞增殖。LI等[8]研究发现,ARNT2在肝细胞癌中的表达明显高于癌旁组织,并与肿瘤的增殖、侵袭和转移密切相关。LI等[9]研究发现,ARNT2是乳腺癌中与侵袭转移有关的7个核心基因之一。以上研究表明,ARNT2在肿瘤的发生、发展中发挥了重要的作用,其在肿瘤中的表达具有组织特异性。
CREB1编码一种转录因子,是DNA结合蛋白亮氨酸拉链家族的成员。该蛋白以同型二聚体的形式与cAMP反应元件(八聚体回文组)结合。该蛋白被多个蛋白激酶磷酸化,并诱导基因转录以响应对cAMP途径的激素刺激。CREB1作为一种重要的原癌基因,作为蛋白激酶A(PKA)的下游效应器,参与组成cAMP/PKA/CREB信号通路,可以调节癌症细胞的生长、迁移、侵袭和代谢[10]。有研究表明,CREB在许多生长信号通路下游的位置暗示了其在肿瘤细胞中的致癌潜力[11]。肿瘤生长与CREB表达和活化增加有关,沉默CREB1可抑制肿瘤的生长[12-13]。
TGF-β2编码TGF-β蛋白超家族的分泌配体。该家族的配体结合各种TGF-β受体,导致调节基因表达的SMAD家族转录因子的募集和激活。编码的前蛋白被蛋白水解处理以产生潜伏相关肽(LAP)和成熟肽,并且以由成熟肽同二聚体、LAP同二聚体和潜在TGF-β结合蛋白组成的潜伏形式存在,或者以仅由成熟肽同二聚体组成的活性形式存在。成熟肽也可以与其他TGF-β家族成员形成异二聚体。TGF-β/SMAD途径的破坏与多种人类癌症有关。YANG等[14]研究表明,TGF-β2在胃癌中高表达,并与肿瘤的组织学分级、淋巴结转移、免疫细胞浸润和预后不良相关。
KRAS是哺乳动物ras基因家族的Kirsten-ras癌基因同源物,编码一种属于小GTPase超家族的蛋白质。一个单一的氨基酸取代是激活突变的原因。转化蛋白与多种恶性肿瘤有关,包括肺腺癌、黏液腺瘤、胰腺导管癌和结直肠癌。KRAS被认为是人类癌症中最常见的突变致癌基因[15]。研究证明,使用KRAS诱导系统可建立许多小鼠模型[16]。YOON等[17]研究表明,17%的胃腺癌发生KRAS的扩增和突变,敲低KRAS可抑制人胃腺癌细胞的上皮细胞-间充质转化和转移。
PDGF-β是由PDGF和血管内皮生长因子组成的蛋白家族成员。编码的前蛋白被蛋白水解处理以产生血小板衍生生长因子亚单位β,其可与相关的血小板衍生生长因子亚单位α同聚或异聚。这些蛋白结合并激活PDGF受体酪氨酸激酶,在广泛的发育过程中发挥作用。该基因突变与脑膜瘤相关。22号和17号染色体之间的相互易位,位于该基因和Ⅰ型胶原α1基因所在的位点,与皮肤纤维肉瘤(一种罕见的皮肤肿瘤)有关。有研究表明,PDGF-β在胃癌组织中高表达,并与肿瘤浸润深度、淋巴结转移及TNM分期呈正相关,检测PDGF-β水平可作为判断胃癌转移和预后的指标之一[18],PDGF/PDGFR信号通路的激活通过调节多种下游通路,包括磷脂酰肌醇3激酶/蛋白激酶B通路和丝裂原活化蛋白激酶/细胞外信号调节激酶通路,与癌症增殖、转移、侵袭和血管生成相关。因此,靶向PDGF/PDGFR信号通路已被证明是癌症治疗的有效策略[19]。
COPB2是典型的细胞质蛋白复合体的1个亚基,其与双赖氨酸基序结合,并与高尔基衍生的非网格蛋白包被的囊泡结合。在蛋白质生物合成过程中,COPB2以囊泡的形式将其他蛋白质从内质网运送至高尔基体[20]。COPB2参与调节多种肿瘤的生物学行为,可在肺癌[20]、乳腺癌[21]等多种恶性肿瘤中异常表达。AN等[22]研究表明,COPB2在胃癌中过表达,沉默COPB2可抑制细胞增殖,减少细胞集落的形成并诱导细胞凋亡。
本研究结果显示,miR-335-5p上调表达抑制胃癌细胞增殖和侵袭能力,表达谱芯片筛选结合生物信息学靶基因预测分析取得交集,共获得212个miR-335-5p的靶基因,并富集在20个肿瘤相关的信号通路中。mRNA和Western blot验证结果表明,靶基因CREB1、AKT3、COPB2、ARNT2、TGF-β2、KRAS、PDGF-β是miR-335-5p调控胃癌发生、发展及预后的重要癌基因。检索文献和数据库均显示,CREB1、AKT3、COPB2、ARNT2、TGF-β2、KRAS、PDGF-β广泛参与胃癌细胞的增殖、侵袭、转移表型。本研究发现,miR-335-5p上调表达可明显抑制胃癌细胞增殖和侵袭能力,所筛选的212个靶基因,包括CREB1、AKT3、COPB2、ARNT2、TGF-β2、KRAS、PDGF-β均富集于20个与肿瘤相关的信号通路中,CREB1、AKT3、COPB2、ARNT2、TGF-β2、KRAS、PDGF-β是经生物信息学方法表达谱芯片筛选的交集基因,并经PCR实验和Western blot实验验证,由此说明miR-335-5p作为胃癌中的抑癌基因,其抑制胃癌细胞增殖和侵袭能力是由靶向调控CREB1、AKT3、COPB2、ARNT2、TGF-β2、KRAS、PDGF-β所介导的。miR-335-5p上调表达抑制胃癌细胞增殖和侵袭能力与抑制靶基因CREB1、AKT3、COPB2、ARNT2、TGF-β2、KRAS、PDGF-β的表达有关。此外,表达谱芯片结合生物信息学手段是重要的组学筛选的方法,本研究中所发现的差异表达基因所富集的信号通路和筛选的核心基因均能说明miR-335-5p在胃癌中是影响表型的关键分子,是调控胃癌生物学表型的观察窗口。另外,通过表达谱芯片结合生物信息学分析发现的具有高度相关性的癌基因和信号通路为探讨miR-335-5p调控胃癌的机制提供了线索和依据。
