
毛木耳育种与菌种扩繁技术研究进展
2023-12-20 07:01李小林杨学圳张小平
农学学报 2023年10期
叶 雷,李小林,张 波,杨学圳,谭 伟,张小平
(1四川农业大学资源学院,成都 611130;2四川省食用菌研究所,成都 610066)
0 引言
毛木耳(Auricularia cornea)为木耳属(Auricularia)的大型真菌,主产亚洲,曾用学名A.polytricha和A.nigricans,俗称耳子、大木耳、厚木耳、沙耳、猪耳等,产品名分为黄背木耳和白背木耳[1-3]。据中国食用菌协会统计,四川、山东、福建、河南等至少17 个省(自治区、直辖市)有毛木耳栽培,2021 年鲜产达220.69 万t,以四川最高为85.72 万t,中国已成为最大的毛木耳生产和消费国。毛木耳的栽培始于驯化,经过了从段木到代料栽培的发展过程。中国毛木耳从段木栽培转向利用农林业副产物的代料栽培始于20世纪70年代末80年代初[4-5]。毛木耳栽培技术体系的进步,与适应性品种的催生相配套,一代代“蘑菇人”通过系统驯化选育、杂交育种、原生质体融合技术等技术手段育成系列高产优质、高抗性、颜色各异的栽培品种,实现品种的更新迭代。形成了四川主栽黄背木耳‘上海1号’、‘781’、‘川琥珀木耳1号’等,福建主栽白背木耳‘漳耳43-28’和‘43012’等,江苏主栽‘苏毛3 号’,河南主栽黄背木耳‘951’、‘781’和‘丰毛6号’等的区域性分布格局。
优良的品种决定了产业的发展未来。近40 a的毛木耳育种工作中,形成了驯化(系统)选育、杂交育种、诱变育种和分子育种等育种技术,并成为毛木耳新菌株选育主要手段,但鲜见关于毛木耳育种工作的总结报道。近年来,现代分子育种技术和新型育种设备高速发展,为毛木耳育种提供了新思路、新方法,因此有必要对毛木耳育种技术和菌种扩繁内容进行总结、回顾,对最新研究进展进行介绍。笔者基于国内毛木耳育种现状,对毛木耳驯化(系统)选育、单单杂交、单双杂交、多孢杂(自)交、原生质体融合育种、诱变育种和分子育种等主要育种技术进行概述,提出毛木耳育种技术与菌种扩繁发展方向,供业内人员交流和讨论。
1 毛木耳育种技术与新品种
1.1 驯化(系统)选育
驯化(系统)选育是毛木耳新菌株选育最常用和最传统的方法,是一种从自然获取优良菌株的方法,一直沿用至今,是国内毛木耳人工栽培新菌株主要来源。驯化(系统)选育出发菌株为野生菌株或人工栽培特异株,经过菌株初筛、复筛、区域试验和生产试验,对典型株产量、品质和抗性等进行记录,最终获得亲本来源和选育过程清楚、遗传性状稳定、田间表现整齐一致优良新菌株的过程[6]。目前该选育方法仍在食用菌育种中大规模应用。
中国木耳栽培大约始于7 世纪,李时珍《本草纲目》“木耳”条引甄权《唐本草》的记述:“煮浆粥安诸木上,以草复之,即生蕈尔”。古代称木耳为“木栭”、“木菌”、“木蛾”,可能包括有黑木耳(A.heimuer)和毛木耳(A.cornea)2 个种,可见当时已认识到木耳的生活规律,发明了“原木自然引种法”[7-9]。毛木耳的人工栽培始于20 世纪70 年代末80 年代初[10](约1975 年),早期毛木耳的栽培菌株是野生驯化选育或引进[11-12]。在毛木耳产业发展初期,科研工作者们开展了大量的引种、驯化、栽培、病虫害防控等工作。育成‘紫木耳’[13]、‘乳白毛木耳’[14-15]、‘AP.4’[16]、‘Au897’[17]、白背毛木耳‘978’[18]、‘台毛43’[17]、‘Au119’[19]、‘桂毛7号’[20]、‘申耳4号’[21]、‘申耳1号’[22]等系列新菌株或品种并开展了品种配套应用技术研究工作。
1.2 杂交育种
毛木耳为担子菌,子实体成熟后会弹射孢子,一般36 h 后孢子萌发形成单核菌丝体[23],单核菌丝的形成为杂交育种提供了材料。杂交育种主要用于异宗结合食用菌,如毛木耳[23]、平菇[24]等,是将遗传特性不同的单核体进行单单杂交、单双杂交、多孢杂交、原生质体融合等,通过有性杂交(或人为介入)发生基因重新组合后,在杂交一代中筛选杂种优势(杂合子)的过程。毛木耳交配类型为四极性异宗结合[25],单核菌丝不能完成其生活史,2 种极性不同的单核菌丝间配对(杂交)产生双核菌丝(具锁状联合)才能完成其生殖过程(图1)。杂交育种对毛木耳亲本配对过程进行人为干预,其杂交类型一般可分为有性单核体杂交和无性单核体杂交,且根据亲本遗传距离关系,可进一步细分为自交、近交、异交、远交等[26]。
1.2.1 单单杂交育种 单孢杂交是采用单孢分离或原生质体单核化获得毛木耳单核菌丝体,再通过单核菌丝杂交配对组合获得杂交子,并通过锁状联合等确定异核体,经出菇(耳)性鉴定具有出菇(耳)能力,遗传区别于亲本的新菌株,进一步进行熟料袋栽初筛、复筛和区域试验,最终获得优良新菌株(表1)。孙华瑜[27]等介绍,Barnett在1937年应用单孢组合新技术证明同为木耳属的黑木耳为单因子异宗交配系统。小松光雄等[28]通过白色毛木耳单核菌丝杂交试验表明,白色毛木耳的颜色性状是由基因决定的,通过与有色野生型菌株杂交,可产生有色子代。王波等[29]采用黄背木耳分别与‘781’、‘951’和‘243’单孢杂交,育成了‘毛木耳1号’、‘毛木耳4号’和‘毛木耳5号’。四川省食用菌研究所专家采用单孢杂交育成了经全国食用菌品种认定委员会认定的品种(以下简称“国认品种”)‘川耳1号’、‘川耳7号’等。冀宏等[30]以江苏丰县的白背毛木耳‘丰2’为亲本进行种内杂交(近交),证明毛木耳种内杂交可产生具结实能力的杂交后代。李婧[25]以毛木耳褐色菌株‘黄耳10号’、‘AU2’和白色菌株‘APW822’为亲本,采用单孢杂交育种方法选育获得产量高、抗性强、性状优良的‘AU55’菌株。巫鹏飞[31]以毛木耳变异菌株‘AP2010’及3 株白背毛木耳菌株(‘4328’、‘43012’和‘AP201’)为亲本,进行单核菌株杂交育种,并结合RAPD分子标记和ISSR聚类分析等进行研究,获得农艺性状较好的‘AP2010-4’和‘43012-12’菌株。袁滨等[32]采取单孢杂交育种技术,以福建‘43012’白背毛木耳和四川‘AP2010’毛木耳白色突变种为亲本,选育出了高产优质耐高温的白背毛木耳新菌株‘杂10’。姚方杰等[33]以毛木耳栽培菌株ZP(浅褐色)和野生菌株YZ(白褐色)为亲本,单单杂交最终得到5个深色的杂交新菌株,且耳片宽度、厚度和产量等性状优于亲本。
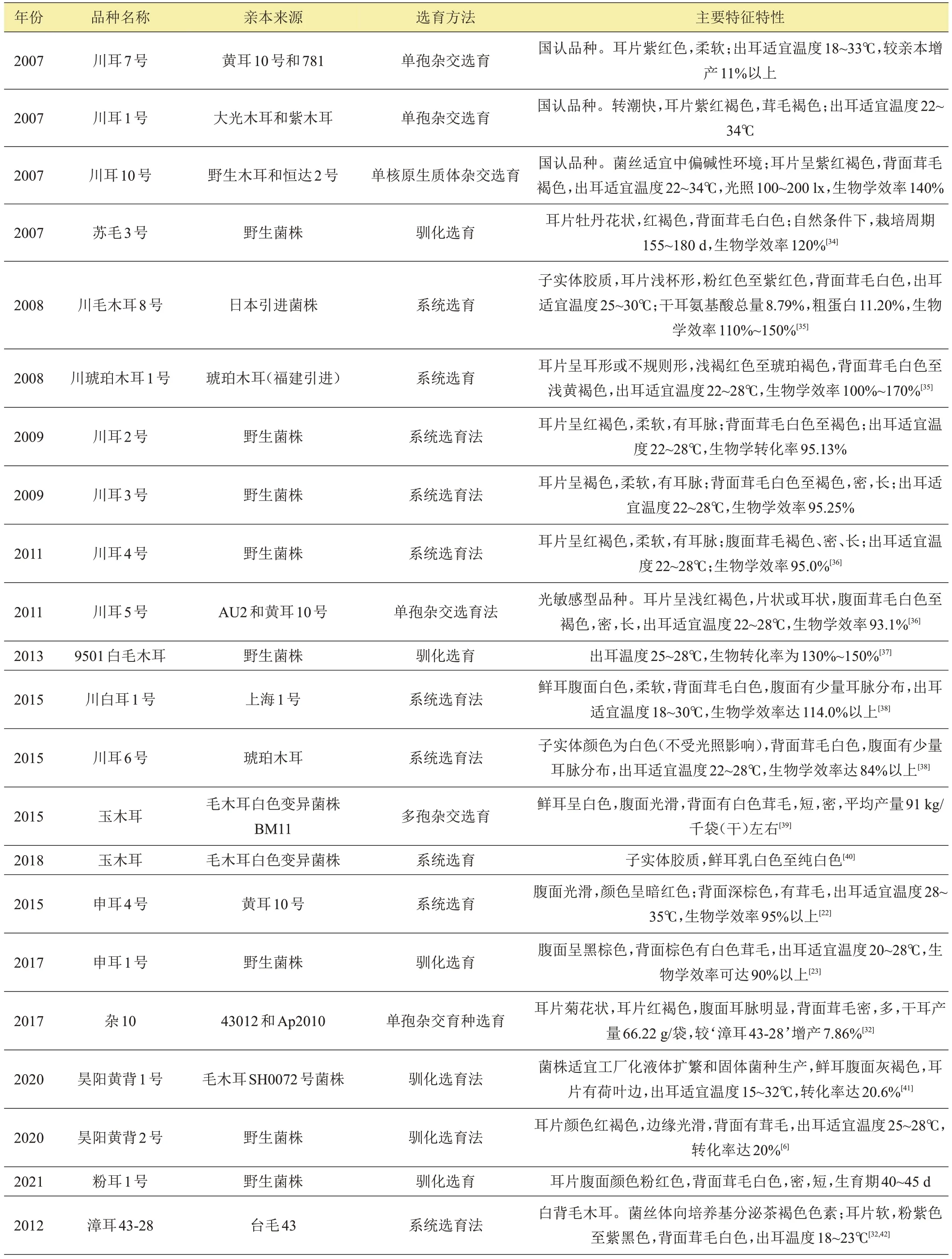
表1 部分选育新品种
此外,国内较多学者采用单孢杂交技术选育出食用菌新品种。赵光辉[44]以草菇(Volvariella volvacea)‘V0045’、‘V0062’、‘V0073’、‘V0032’、‘V0053’为杂交亲本,通过出菇试验、RAPD和ISSR分子标记验证6个草菇优势新菌株。张树强[45]以引自江苏高邮的鸡腿菇(Coprinus comatus)‘特白36’和新疆阿拉尔的野生鸡腿菇为亲本,优选得到3 株杂交鸡腿菇新菌株。王波等[46]采用金针菇(Flammulina filiformis)单单杂交选育新品种。张敏等[47]以滑菇(Pholiota microspora)‘PN06’和‘PN08’为亲本,育成杂交新菌株‘辽滑菇1号’。谭伟等[48]以‘姬菇258’和‘金凤2-1’为亲本,选育出了4 个杂交姬菇新菌株。林锋[49]以3 株草菇高产菌株和耐低温菌株‘VH3’为亲本,通过单孢杂交技术并结合RAPD 技术,选育出了草菇耐低温的高产优良菌株‘VV093’。王艳[50]以灰树花(Grifola frondosa)野生菌株‘梯灰1 号’和栽培品种‘庆灰151’为亲本,各取10 个孢子单核体进行单单杂交,通过对杂交菌株平板拮抗试验和菌丝锁状联合观察,初筛出65 个杂交子,并通过出菇试验、ISSR 和SRAP 分子标记鉴定,获得了7 个杂交新菌株。目前,单单杂交选育新品种技术已经非常成熟,已大量使用并成为毛木耳杂交育种首选方法。
1.2.2 单双杂交育种 单双杂交育种是将亲本单核菌丝体与双核菌丝体转接至同一培养基上进行杂交选育新菌株的方法。关于毛木耳采用单双杂交选育优良菌株的研究几乎未见报道。但有学者采用单双杂交系统成功选育出香菇(Lentinus edodes)优良菌株(‘D100FP’和‘D97FP’)[51],育成四川栽培面积最大的黄色金针菇‘川金3号’[52]、白色金针菇单双杂交子‘F374506’[53]、滑子菇‘丹滑16 号’[54],以及糙皮侧耳[55]和黑木耳[56]优良杂交子菌株。
1.2.3 多孢杂(自)交育种 多孢杂(自)交是将(不)同亲本多孢子混合物放置在同一培养基上培养,后挑取单菌落菌株进行锁状联合观察,再与其他挑取菌株和亲本进行拮抗,最终通过出菇试验获得优良菌株的方法。多孢自交本质上是一种单单自交。贺建超等[57]将毛木耳‘C14’和黑木耳‘C21’进行多孢杂交,结果获得6株与亲本具有明显拮抗效应的子代且产量和抗逆性明显高于黑木耳亲本。李婧[25]对毛木耳F1杂交种和F2多孢自交分离种进行遗传多样性分析,表明F1杂交种的组织分离种遗传稳定性高于F2多孢分离种。任梓铭[58]以毛木耳‘MZP01’(红褐色)和毛木耳‘MZP03’(白色变异株)为亲本进行多孢杂交,结果选育获得‘MZF02’与‘MZF05’2个菌株用于复筛。
1.2.4 原生质体融合育种1967 年Strunk 等首次分离并观察到担子菌原生质体自发融合[59]。Devriers 等对裂褶菌(Schizophyllum commune)原生质体的分离成功,开启了原生质体(单核或双核)融合育种技术在食用菌研究领域的应用,约在20世纪80年代中后期达到研究高峰[60]。应用原生质体融合技术已实现食用菌科、属和种间原生质体的融合,克服物种间不亲和性,获得远缘融合新菌株。该技术属细胞工程育种新技术领域,关键技术环节在于原生质体的形成、融合、再生及融合子的获得。罗信昌[61]、肖在勤等[62]、彭卫红等[63-65]、王波等[66-67]、邱敦莲等[68]、郑林用等[69]、孙露等[70]、钱可晴等[71]对毛木耳、金针菇、凤尾菇、茯苓、灵芝(Ganoderma lingzhi)、长根菇、鸡枞菌、草菇、双孢蘑菇(Agaricus bisporus)和茶树菇(Agrocybe aegerita)等亲本原生质体的制备、再生条件以及细胞融合、融合核分裂等内容进行了研究。汪麟[72-73]采用黑木耳和毛木耳原生质体融合技术,育成了种间杂交黑木耳新品种‘冀杂2 号’和‘冀杂3 号’,李育岳等[74]进一步进行菌株特性研究,表明杂交菌株遗传稳定,在耐热性、抗霉性、产量和商品性等方面优势明显。1999年,四川省食用菌研究所专家研究成果“应用细胞融合技术选育食用菌优良新品种”获国家技术发明二等奖,突破了融合核分裂技术,解决了食用菌远缘融合异核体遗传不稳定、核配菌株形成子实体难的世界性难题[60]。贺建超等[75]获得黑木耳‘C22’热灭活和毛木耳‘C16’紫外灭活原生质体融合优良菌株。彭卫红[76]以毛木耳‘恒达2号’为亲本制备双核原生质体,在同一亲本再生菌株中性状差异较大,且获得与亲本产量相当的‘03-1’菌株,进一步与野生木耳单核原生质体杂交育成了国认品种‘川耳10 号’。近年来,关于毛木耳原生质体融合育种的研究鲜见报道。
此外,郑林用等[69]将金针菇和凤尾菇具锁状联合的菌丝制备原生质体后进行科间融合育种育成‘金凤2-1’。谭琦等[77]以香菇‘Le1’”和野生种0426 为亲本,单核原生质体杂交育成‘申香8号’。彭卫红等[78]应用原生质体不对称融合方法,筛选到金针菇白色优良菌株‘金白1 号’和‘金白2 号’。甘炳成等[79]以香菇单核原生质体进行杂交,筛选获得了优良菌株‘JW1’和‘JW3’。杨珊[80]获得了猴头菌(Hericium erinaceus)原生质体制备及再生最佳条件,以‘0605’和‘刺长’猴头菌株为亲本,采用单核体杂交和原生质体融合育种技术,并通过对融合子和杂合子的筛选、鉴定及出菇试验,对子实体活性物质积累、营养成分及农艺指标观测,获得了优良猴头菇菌株新材料。张鹏[81]以杏鲍菇(Pleurotus eryngii)和秀珍菇(Pleurotus geesteranus)为原生质体融合亲本,经高温灭活后利用聚乙二醇(PEG)诱导融合,并观察锁状联合、拮抗现象,结合RAPD、SRAP、ISSR 及SCAR 分子标记技术进行融合子鉴定、遗传分析,最终获得秀珍菇融合子‘R1-16’。张亚娇[82]将白灵菇(Pleurotus nebrodensis)与杏鲍菇进行远缘杂交,得到了杏白灵双向核迁移杂交子菌株。谢宝贵利用杏鲍菇和白灵菇杂交,育成了具有杏鲍菇特性的‘杏白030’和‘杏白081’等杂交新菌株[83]。此外,还获得了白灵菇原生质体制备和再生最佳条件,并采用原生质体再生无性系及其紫外诱变育种技术,并经RAPD 验证,筛选到经紫外照射10 s 的优良变异菌株‘10s n29’[82]。综上,原生质体融合育种技术在食用菌育种领域应用具有开拓性,在多种菌类育种上是行之有效的,实现了细胞间远缘融合(杂交),可获得双亲优势互补新菌株或不育株,在食用菌遗传育种中具有重要意义。
1.3 诱变育种
诱变育种是人为采用物理、化学诱变剂等直接或间接地诱发出发菌株DNA碱基发生可遗传突变(碱基错配、点突变等),使遗传特性发生变化,再从突变体中筛选新菌株的过程。毛木耳的诱变育种鲜见报道。
物理诱变中,传统的诱变剂方法是采用紫外线、辐射等,但随着生物技术的发展,近年常压室温等离子体(atmospheric room temperature plasma,ARTP)作为新型物理诱变方法,在菌类育种中具有较多应用。在毛木耳传统物理诱变中,诱变体可从单(双)核菌丝体、原生质体、孢子等获得,结合其他杂交育种技术进行新菌株选育。陈文良[84]将大木耳孢子液经紫外线诱变(15 W,30 cm 处),育成‘北京大木耳’。杨新美等采用60Co 射线辐射诱变光木耳和琥珀木耳获得9 株营养缺陷型单核菌株。在ARTP 诱变育种中,目前仅见刘微[43]以‘毛木耳3 号’为出发菌株,采取ARTP诱变仪进行物理诱变育种,在316株杂合菌株中初筛获得26株与出发菌株有明显拮抗现象的突变株,再经复筛、ISSR 分子标记和栽培研究,育成毛木耳新菌株‘新河大SL205’。据介绍,该菌株可有效利用黑木耳菌糠,生物学效率达126.29%。作为新兴诱变育种技术,ARTP 在滑子菇[85]、灵芝[86]、北虫草[87]、花脸香蘑[88]和草菇[89]等菌类育种中快速应用,成效显著。相信在接下来的5~10 a,ARTP 在毛木耳育种中将发挥更大作用。
1.4 分子育种
分子育种包括分子标记辅助育种(marker-assisted breeding,MAB)、基因工程育种、基因编辑和分子设计育种、全基因组选择(genomic selection,GS)等,具有快速、准确、抗干扰强的优点。与大作物相比菌类作物的分子育种起步较晚,但近年取得了一定成效,特别是分子标记辅助育种的应用。
分子标记辅助育种将传统育种技术与现代分子技术相结合,利用分子标记与特定效应基因或性状(耳片颜色、菌丝生长速度、产量等)基因紧密连锁,从而通过检测分子标记而准确鉴别不同后代个体基因型[90]。较广泛应用的标记有随机扩增多态性(RAPD)、相关序列扩增多态性(SRAP)、内部简单重复序列(ISSR)、SSR、目标区域扩增多态性(TRAP)、特征序列扩增区域标记(SCRA)以及第三代分子遗传标记(SNP)[91]。选择合适的分子标记才能构建高质量遗传连锁图谱。卢兴潮[92]基于毛木耳杂交菌株‘APM2-16’的123 个孢子单核体后代作为作图群体,构建转录组ESTs 分子标记遗传连锁图谱,并定位了菌丝体生长速度相关的QTLs,周雁等[93]对该菌株开发了系列基因内分子标记SSR、SCAR、CAPs、InDel 等,选用373 对InDel 引物对亲本单核体间群体基因型分型和多态性分析,获得了一张由14个连锁群组成,含有160个基因内标记的毛木耳遗传图谱。黄蓉梅等[94]对毛木耳‘43012H’白色突变体耳片颜色相关SNP 标记开发,获得SNP1(AC_B 特有)和SNP2(AC_W 特有)用于区分褐色(正常)和纯白色(突变)耳片性状。崔玮洁等[95]获得了玉木耳原生质体制备最佳条件,并以玉木耳单核全基因组为参考,对双核重测序基因组进行分析,发现1432175 个SNPs 和251663 个InDel 位点。Jia 等[96]发现蛋白磷酸酶2A 调控亚基(PP2A)和多聚泛素(UBQ)是毛木耳中一组可用的内控基因,可用于该物种未来的分析。黄艺宁[97]联合RAPD、ISSR分子标记对10个毛木耳菌株进行了鉴定和遗传多样性分析,为育种亲本选择奠定了基础。
关于毛木耳基因工程育种(转基因)、基因编辑、分子设计育种和GS方面研究几乎未见报道。关于重要功能基因的分离和克隆研究鲜见报道,且得到的功能基因大多仍处于功能研究阶段,育种实践中还未应用。贾定洪等[98]构建了毛木耳农杆菌介导转化的Mfcsg、Hpt潮霉素B抗性基因表达载体,为毛木耳多功能纤维素酶基因Mfc转化研究奠定基础。苏文英等[99]对玉木耳Cupredoxin 超家族基因Aclac漆酶基因(cDNA序列长度1734 bp)进行克隆及原核表达,为该基因在玉木耳生长过程中的表达及菌株遗传改良研究奠定基础。GS 策略由Meuwissen 等[100]在2001 年提出,是通过高密度分子标记,覆盖全基因组分子标记数据或单倍型数据和每个样本的表型数据来建立预测模型,估计每个标记的遗传效应,进而在后续的育种中预测个体的全基因组育种值(genomic estimated breeding value,GEBV),以此来进行高效选择育种的新方法。目前还未见该技术在毛木耳育种中的应用,可能是受测序成本、测序方法、数据处理慢(计算机配置要求高)、芯片技术、个体分型费用及分型准确性不高等因素限制。相信随着分子生物学技术的进步、成本的降低,基因编辑CRISPR/Cas9、GS 等新育种技术将在毛木耳分子育种中逐步应用,并取得显著成效。
2 毛木耳菌种扩繁技术研究
优良的菌株配套合适的扩繁方法才能发挥品种最大效力。毛木耳菌种扩繁分固体和液体培养。限于毛木耳特殊种性(中高温出菇、需氧量大、生育期长等),液体菌种对设备要求高、投资大、技术要求高等原因,毛木耳工厂化液体菌种生产没有得到大面积应用,实践中仍以固体菌种扩繁为主。
2.1 固体菌种扩繁研究
毛木耳固体菌种采用人工合成培养基或原木进行菌株培养,具有运输方便、杂菌污染易观察、储存时间长的优点,一般扩繁三级。一级种培养基多用PDA或改良PDA 进行扩繁。黄艺宁等[101]报道了白背木耳较佳碳源为葡萄糖和蔗糖,较佳氮源为蛋白胨,且获得了生产建议改良PDA配方(马铃薯200 g/L、葡萄糖5 g/L、蔗糖15 g/L、琼脂20 g/L)。此外,马铃薯葡萄糖琼脂综合培养基、蛋白胨加富PDA 培养基(蛋白胨2 g、葡萄糖20 g、K2HPO41 g、KH2PO40.5 g、MgSO40.5 g、VB10.5 mg,水1 L)等均可用于一级菌种的培养[102]。二级和三级种多采用天然产物合成培养基,木粒、枝条[103]等原木进行扩繁。如‘台毛1 号’[104]用36%桑枝屑、36%棉籽壳、18%桉树木屑、7%麸皮、2%石灰、1%石膏。白背木耳‘43-28’[105]用棉籽壳30%、木屑50%、麦皮18%、石灰1%、碳酸钙1%。黄背木耳用棉籽壳46%、玉米芯14%、米糠18%、木屑14%、玉米粉3%、石灰3%、石膏1%和白糖1%。陈丽新等[106]基于广西桑枝屑、桉树屑和棉籽壳原料,获得4个质优价廉的毛木耳菌种扩繁配方。谭伟等[34]对二级和三级菌种扩繁原料选择、灭菌、接种和发菌关键环节进行了技术总结。近年来,四川为解决接种效率低、劳动强度大、接种技术人员老龄化等问题,研发了塑料袋(瓶)装菌种、扣盖菌种、木粒菌种及配套料袋封口装置,并通过“科研单位+龙头企业+协会+农场(专合社)+农户”的联动模式取得了较好的应用效果。
2.2 液体菌种扩繁研究
毛木耳液体菌种具菌丝萌发快、制种周期短的特点。主要研究内容集中在液体配方、培养条件(温度、转速、通气速率、三角瓶或发酵罐装液量、培养时间等)、菌种扩繁技术及液体发酵菌丝体功能活性研究[107]等方面。1986 年吴自强等[108]就将毛木耳液体菌种应用于稻草培养基接种,表现出菌丝长速快、污染率低和单产高的优点。高峰等[109]获得黄背木耳液体配方(葡萄糖3%、麦麸10%、MgSO4·7H2O 0.02%、KH2PO40.1%),且认为液体菌种比固体菌种成本低、发菌速度快。唐业刚[110]获得了毛木耳‘988’三角瓶液体发酵培养基及培养条件(马铃薯20%,蔗糖2%,玉米粉1.5%,蛋白胨0.6%,KH2PO40.1%,MgSO4·7H2O 0.02%。温度25℃,转速160 r/min)。刘孝利等[111]获得玉木耳最佳液体培养基配方(豆粉和玉米粉各2.5 kg,蔗糖和葡萄糖各5.5 kg,KH2PO41.0 kg,MgSO40.5 kg),按浓度2%~2.5%配制培养基,23℃培养96 h 时干菌丝量达0.85 g/100 mL,但没有对转速和通气速率进行介绍。刘敏等[112]获得白背毛木耳‘川耳1 号’液体培养基及培养条件(马铃薯20%、葡萄糖2%、酵母粉1.5%、MgSO4·7H2O 0.1%、KH2PO40.2%,温度25℃,转速180 r/min)。叶雷等[6,41,113]育成液体优良菌株‘昊阳黄背1 号’、‘昊阳黄背2 号’,并依托主产区龙头企业开展液体菌种生产应用[114]。随着中国种业振兴计划的实施,液体菌种在菌种(棒)集约化生产中将得到进一步应用。
3 毛木耳育种技术与菌种扩繁发展方向探讨
3.1 毛木耳的杂交育种工作深入发展
在毛木耳长期育种发展史中,驯化(系统)选育、杂交选育是最成熟常用的育种手段。杂交选育可获得双亲优势互补的杂种优势后代,成为较多科技人员开展毛木耳菌株新材料创制的首选方案。在杂交育种技术领域仍然有较多科学问题需要回答[26],未来研究中高效、准确的从杂交后代定向预测获得目标典型菌株,减少育种工作量和提高育种效率,将分子标记辅助育种、基因编辑等分子育种技术结合进来,开展全基因组学选择,将促进杂交育种工作深入发展。
3.2 现代高效育种技术得到快速应用
近年,ARTP、GS、CRISPR/Cas9 和MAB 等现代高效育种技术在金针菇[115]、双孢蘑菇、草菇[116]、茶树菇[117]等育种领域得到快速应用,取得一定成效,但在毛木耳研究领域鲜见报道。毛木耳基因组、极性、功能基因挖掘、系统发育、配子体杂交、重要性状分子标记等育种相关基础研究已取得一定成效且处于发展阶段,现代高效育种技术的发展应用为毛木耳育种技术进步带来了前所未有的发展机遇和挑战。
3.3 毛木耳商业化育种得到积极倡导
育种工作是理论与实践的结合。毛木耳主要依托科研单位科技人员开展新品种选育示范工作,由于缺乏对市场的调研,育种者选育目标往往具有盲目性、随意性,科技成果转化率普遍不高,市场突破性品种缺乏。开展毛木耳商业化育种[118],能实现育种者以市场、经济、价值等问题为导向,育种目标与市场需求完美契合,菌株知识产权得到及时保护,市场开放获取资源,符合国家种业发展需求,应大力倡导。
3.4 毛木耳菌种“育繁推”体系逐步成熟
国内毛木耳菌种生产仍以传统农业式生产为主,但菌种的现代化、标准化、专业化生产是必然。有研究显示,毛木耳菌种产业链普遍存在种源复杂(同物异名)、菌株权益维护难、越级生产、设备设施落后、菌种质量参差不齐(无标准)、菌种检验不合格、管理不规范和缺乏资质企业牵头制种的问题[119-122],不利于毛木耳产业健康发展。近年曲靖种业走上“育繁推一体化”[123],贵州食用菌打造“育繁推”体系实现种业创新[124],弥补了菌种产业链不足。2022 年,四川在毛木耳“育繁推”一体化良种繁育基地建设方面已取得成效,实现20 余万瓶原种、40 余万瓶栽培种的集中繁育和300 余万料袋集中制袋和灭菌。在《中华人民共和国种子法(2022修订版)》和种业振兴行动方案的指导下,菌种“育繁推”一体的产业体系和生产主体将得到大力培育,更好地满足产业发展需求。
猜你喜欢
今日农业(2021年10期)2021-07-28
女士(2017年10期)2017-11-01
食用菌(2017年5期)2017-10-19
作文大王·低年级(2017年2期)2017-02-16
首都医科大学学报(2015年4期)2015-12-16
大连工业大学学报(2015年4期)2015-12-11
特产研究(2015年1期)2015-04-12
食品工业科技(2014年23期)2014-03-11
无机化学学报(2014年12期)2014-02-28
无机化学学报(2014年7期)2014-02-28
