
CRISPR/Cas9介导的KIFC1基因敲除对宫颈癌细胞增殖及凋亡的影响
2023-12-14 09:36:02范晓静魏雅岚叶洲杰朱丽萍王心睿
肿瘤防治研究 2023年11期
范晓静,魏雅岚,叶洲杰,朱丽萍,王心睿,3
0 引言
驱动蛋白14家族成员C1(Kinesin family member C1, KIFC1, NP_002254)是一种从微管正极端向负极端运动的驱动蛋白,KIFC1可以促进微管间的交联、滑动和捆绑,在有丝分裂过程中参与纺锤体极体的聚集和组装,并在含有多个中心体的癌细胞中参与中心体的聚集。关于KIFC1的重要性尚存在争议,但在癌细胞中,KIFC1的过量表达是维持多个中心体肿瘤细胞正常分裂所必需的[1-2]。研究发现KIFC1在乳腺癌[3]、卵巢癌[4]、肝癌[5-6]、胃癌[7]、前列腺癌[8]、胰腺癌[9]等恶性肿瘤中特异性过表达,并可促进肿瘤细胞的增殖、迁移和上皮间质转化[4]。KIFC1通过调节PI3K/AKT信号通路调控子宫内膜癌进程,其表达量与子宫内膜癌的临床分期、病理类型及预后显著相关[10]。此外,KIFC1已被确定为三阴性乳腺癌患者预后不良的生物标志物之一[11]。KIFC1也可诱导多西他赛耐药,并与前列腺癌患者的不良预后有关[12]。敲低KIFC1可以抑制肝癌细胞增殖和侵袭,增加其对紫杉醇的敏感度[13]。因此,KIFC1作为肿瘤选择性治疗的靶点,值得进一步探讨。然而,关于KIFC1在宫颈癌发病机制中的作用研究较少,具体调控机制尚不清楚。
CRISPR/Cas9系统是目前最主流的基因编辑系统,具有效率高、成本低、操作简便等优点,广泛运用于基因敲除、基因敲入和多重基因编辑。本研究运用CRISPR/Cas9系统在人宫颈癌HeLa细胞中构建KIFC1基因敲除细胞系,探讨KIFC1表达缺失对细胞形态、细胞增殖、周期与凋亡的影响,为进一步研究KIFC1蛋白在宫颈癌发生发展中的作用提供细胞模型。
1 材料与方法
1.1 主要材料与试剂
人宫颈癌细胞系HeLa(来源于ATCC细胞库CCL-2编号的细胞系)、CRISPR/Cas9系统质粒pSpCas9(BB)-2A-GFP(简称PX458)源于本实验室保存,DMEM培养基、胎牛血清和0.25%胰酶均购自美国Gibco公司,兔抗人KIFC1和鼠抗beta-actin单克隆抗体均购自英国Abcam公司,辣根过氧化物酶(HRP)标记二抗购自美国Cell Signaling Technology公司,TurboFect转染试剂(R0531)、T4 DNA连接酶(EL0016)和BbsⅠ酶(ER1011)均购自美国Thermo Scientific公司,感受态细胞DH5α(C502-03)、染料法qPCR预混液(Q711-02)、TA克隆试剂盒(C601-01)和Annexin V-FITC/PI凋亡检测试剂盒(A211-02)均购自南京诺唯赞公司,ExTaq酶(RR001Q)和反转录试剂盒(RR047A)均购自日本TaKaRa公司,PMSF(A610425)、RIPA缓冲液(C500005)、EdU细胞增殖检测试剂盒(E607204)和吖啶橙染色试剂盒(E607307)均购自上海生工公司,细胞周期与凋亡检测试剂盒(C1052)购自上海碧云天公司,细胞基因组DNA提取试剂盒(DP304)、无内毒素质粒小提中量试剂盒(DP118)、琼脂糖凝胶DNA回收试剂盒(DP219)、细胞总RNA提取试剂盒(DP451)均购自天根生化科技(北京)有限公司,引物合成和DNA测序由铂尚生物技术(上海)有限公司完成。
1.2 实验方法
1.2.1 靶向人KIFC1基因sgRNA的设计及表达载体的构建
在美国国家生物技术信息中心官网(https://www.ncbi.nlm.nih.gov/)检索Homo sapiensKIFC1基因序列(NM_002263.4),将序列输入到在线sgRNA设计网站(http://chopchop.cbu.uib.no/),根据预测结果设计两对sgRNA序列(见表1)和测序引物hKIFC1 target(见表2)。sgRNA与BbsⅠ酶切后的PX458质粒连接,转化到大肠杆菌,筛选阳性克隆,提取质粒鉴定。
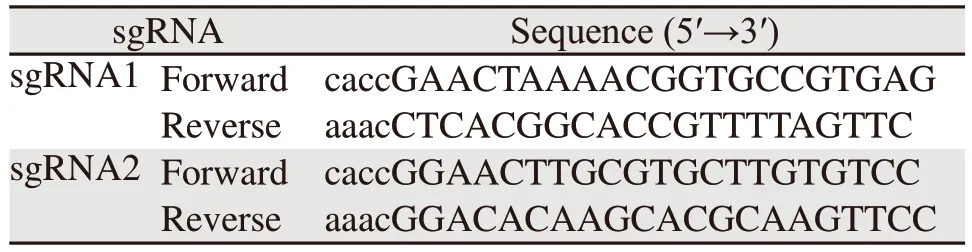
表1 sgRNA寡核苷酸序列Table 1 sgRNA oligonucleotide sequences
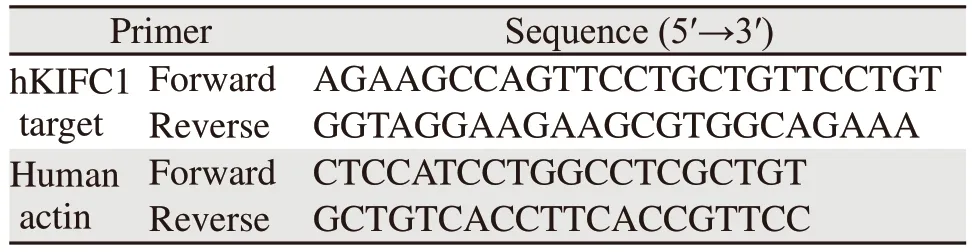
表2 测序及qPCR相关引物Table 2 Primers for sequencing and qPCR
1.2.2 HeLa细胞培养及转染
HeLa细胞在含10%胎牛血清的DMEM培养基中培养,待细胞密度达到70%时,按照转染试剂说明书分组转染,其中实验组为PX458-sgRNA1+PX458-sgRNA2,阴性对照组为PX458质粒,空白对照组为脂质体试剂。转染48和72 h后,使用倒置荧光显微镜(Nikon, Ts2R-FL)观察绿色荧光蛋白表达情况以确定转染效果。
1.2.3KIFC1敲除单克隆HeLa细胞株的筛选及鉴定
转染72 h后的HeLa细胞经胰酶消化吹打成单细胞悬液,流式细胞术分选收集绿色荧光蛋白表达阳性的细胞悬液。采用有限稀释法将单个细胞接种至96孔板中进行培养,待单孔细胞增殖至50%融合度时,传代并扩大培养获得单克隆细胞株。
提取细胞基因组DNA,进行PCR扩增。PCR扩增体系为:PCR程序为:95℃ 3 min;95℃ 15 s,55℃ 20 s,72℃ 30 s,33个循环;72℃ 10 min,4℃保温。胶回收PCR产物并进行TA克隆,挑选单克隆菌落进行菌落PCR验证及测序分析。最终以KIFC1敲除细胞株为实验组,野生型HeLa细胞为对照组进行后续实验操作。
1.2.4 Western blot检测KIFC1蛋白表达情况
使用含1%PMSF的RIPA缓冲液裂解细胞,提取细胞总蛋白。制备好的蛋白样品进行SDSPAGE,电转到PVDF膜上,5%脱脂奶粉封闭1 h,加入一抗4℃孵育过夜,HRP标记二抗孵育1 h,TBST洗涤3次,在膜上滴加显影液,放入显影系统(Bio-Rad,ChemiDoc MP)中成像。
1.2.5 RT-qPCR检测KIFC1基因转录水平
提取细胞RNA,进行反转录反应。以cDNA为模板,采用SYBR Green染料法进行实时荧光定量PCR检测。qPCR反应体系为:2×ChamQ Universal SYBR qPCR Master Mix 10 μl,ddH2O上下游引物各1 μl,模板cDNA 1 μl,补ddH2O至20 μl。qPCR反应程序为:95℃ 30 s, 95℃ 10 s, 60℃ 30 s,40个循环;融解曲线95℃ 15 s,60℃ 60 s,95℃ 15 s。
1.2.6 免疫荧光法检测敲除KIFC1对细胞形态结构的影响
将细胞接种于含12 mm爬片的24孔板中,待细胞贴壁生长后,吸弃培养基,PBS漂洗1次后,4%多聚甲醛固定20 min,PBS浸洗3次。0.25%Triton X-100/PBS透化10 min,PBS浸洗3次,用含3%BSA/PBST的封闭液室温孵育1 h,加入一抗4℃孵育过夜,二抗室温避光孵育1 h,两步分别用PBS浸洗3次去除多余抗体,DAPI与抗荧光淬灭剂等体积混匀封片,激光扫描共聚焦显微镜(Leica SP8)成像。
1.2.7 细胞生长曲线绘制及EdU染色
细胞生长曲线绘制:各组细胞用胰酶消化成单细胞悬液并进行计数,调整细胞浓度为5×104/ml,接种至24孔板中正常培养。24 h后开始计数,连续计数8 d,绘制细胞生长曲线。
EdU染色:细胞爬片过夜培养后,按EdU染色试剂盒说明书进行染色,共聚焦显微镜(Leica SP8)获取图像,检测细胞增殖。
1.2.8 流式细胞术检测细胞周期
收集各组细胞,使用预冷的PBS洗涤和重悬,缓慢加入预冷的75%乙醇,4℃固定过夜。次日,PBS洗涤,加入碘化丙啶(PI)37℃避光染色30 min,上流式细胞仪(BD LSRFortessa)检测,记录激发波长488 nm处的红色荧光。
1.2.9 流式细胞术及吖啶橙染色法检测细胞凋亡
流式细胞术:细胞接种至6孔板中培养48 h后,胰酶消化收集细胞,按Annexin V-FITC/PI凋亡检测试剂盒说明书操作,1 h内上流式细胞仪(BD LSRFortessa)检测。
吖啶橙(AO)染色法:细胞消化后,PBS洗涤两次,按吖啶橙染色试剂盒说明书进行染色,荧光显微镜观察成像。
1.3 统计学方法
所有实验均重复3次,采用GraphPad Prism 7.0和Microsoft Excels软件进行数据分析,结果以均值±标准差表示,两组间比较采用t检验,P<0.05为差异有统计学意义。
2 结果
2.1 KIFC1在HeLa细胞中的时空表达定位分析
为评估KIFC1在HeLa细胞分裂中的功能,我们对KIFC1在细胞间期与分裂期的表达情况及亚细胞定位进行分析,利用免疫荧光技术对HeLa细胞的细胞核、微管及KIFC1蛋白进行荧光标记,测量荧光强度和相对细胞位置,见图1A。结果显示,在细胞间期,KIFC1分散定位在细胞质中或大量转运进入细胞核,见图1B。分裂期,尤其是在有丝分裂后期,KIFC1大量定位在中央纺锤体与星极纺锤体区,参与牵拉染色体的分离,见图1C。
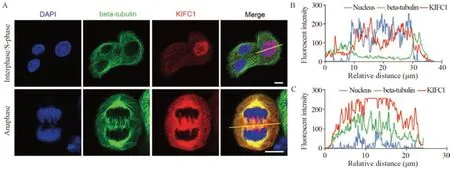
图1 KIFC1在HeLa细胞间期与分裂期的表达定位Figure 1 Location of KIFC1 in HeLa cells during interphase and mitosis
2.2 构建sgRNA-Cas9重组载体
利用BbsⅠ酶对PX458质粒进行线性化处理,将合成的sgRNA序列与线性化PX458质粒连接,经转化、涂板、测序验证,构建sgRNA-Cas9重组载体,见图2。该载体含有人U6启动子,带有Cas9和EGFP标签,并具有氨苄青霉素抗性。琼脂糖凝胶电泳结果显示,酶切成功的质粒在相应位置出现明显条带,见图3A。
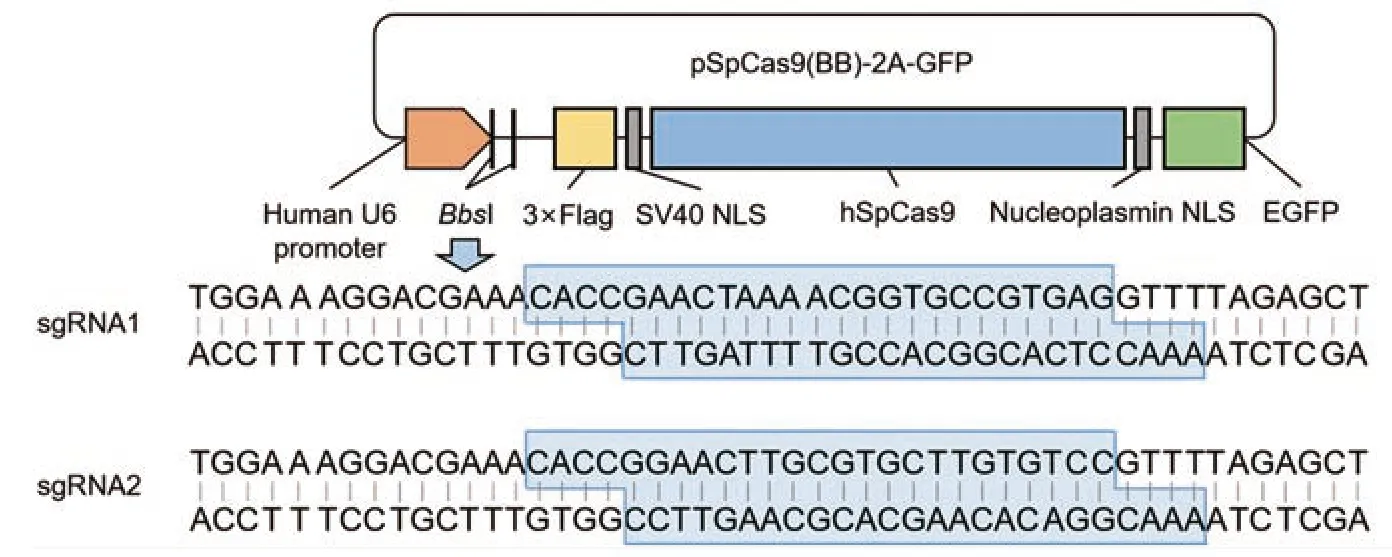
图2 sgRNA-Cas9重组载体构建模式图Figure 2 Schematic diagram of sgRNA-Cas9 recombinant vector construction
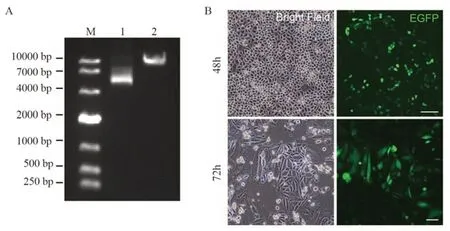
图3 转染质粒后EGFP阳性细胞形态F i g u r e 3 E G F P p o s i t i v e c e l l morphology after plasmid transfection
2.3 sgRNA-Cas9重组载体成功转染至HeLa细胞
将PX458-sgRNA1和PX458-sgRNA2质粒共转染至HeLa细胞中,使用倒置荧光显微镜观察转染效果,转染48 h后可见多数细胞成功表达EGFP信号,72 h后大量EGFP阳性细胞表现出巨核或多细胞核的表型,见图3B。
2.4 流式细胞术分选EGFP阳性细胞
通过荧光流式细胞分选技术,收集EGFP强信号的细胞群,检测其转染效率为28.6%,见图4A,收集该部分细胞群。经过荧光显微镜镜检,可以观察到所有分选后的细胞都携带绿色荧光,见图4B。
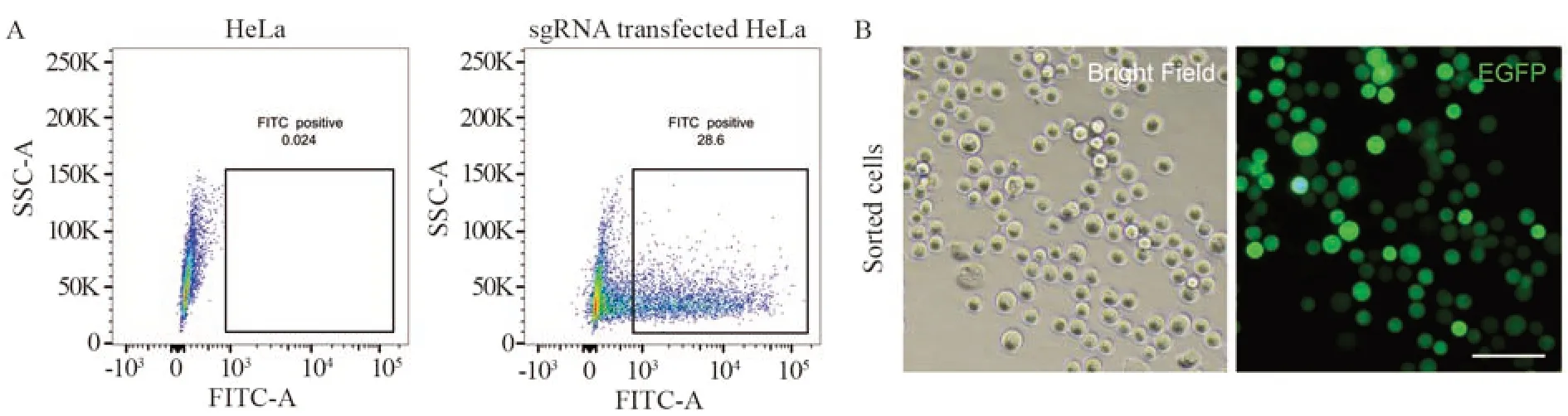
图4 流式细胞术分选EGFP阳性HeLa细胞群Figure 4 Flow cytometry sorting of EGFP-positive HeLa cells
2.5 成功构建KIFC1敲除单克隆HeLa细胞株
靶序列测序分析显示,KIFC1敲除细胞在386 bp和563 bp位点各缺失1个鸟嘌呤(G),在564 bp位点缺失1个胸腺嘧啶(T),见图5A,这些碱基缺失可以造成KIFC1蛋白截断翻译。与对照HeLa细胞相比,敲除细胞株中KIFC1蛋白表达量和转录水平明显下降,见图5B~D,与预期结果一致,证明HeLa细胞KIFC1基因敲除成功。

图5 敲除细胞KIFC1转录与蛋白表达水平验证Figure 5 Verification of KIFC1 transcription and protein expression levels in knock-out cells
2.6 KIFC1基因缺失影响HeLa细胞的形态结构维持
在KIFC1敲除株中,大量细胞核表现出多核、微核、染色质异常包装和微管骨架装配错误,见图6。细胞骨架异常延伸并过度组装,但细胞的贴壁能力明显变弱,细胞的增殖能力降低,部分细胞在生长过程中表现出明显的凋亡或死亡,见图7A。
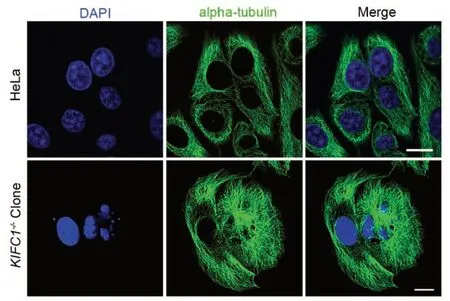
图6 KIFC1敲除细胞微管骨架形态分析Figure 6 Morphological analysis of microtubule cytoskeleton in KIFC1 knockout cells
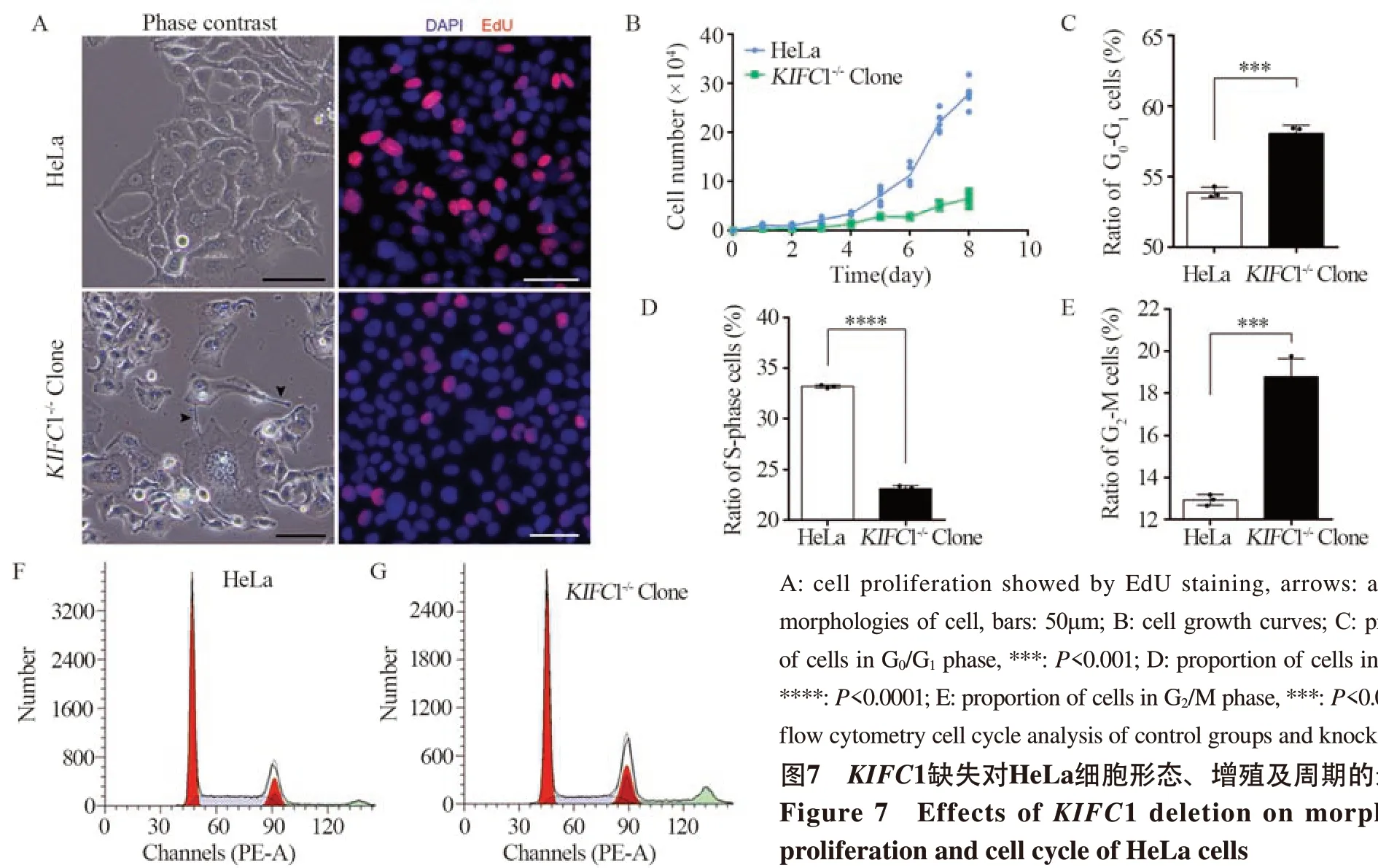
图7 KIFC1缺失对HeLa细胞形态、增殖及周期的影响Figure 7 Effects of KIFC1 deletion on morphology,proliferation and cell cycle of HeLa cells
2.7 敲除KIFC1抑制HeLa细胞增殖并影响细胞周期调控
EdU染色结果显示,与对照组相比,KIFC1敲除组中EdU标记的细胞明显减少,且DNA复制强阳性信号的细胞数量较少, 见图7A。通过分析细胞生长曲线,KIFC1敲除组细胞生长速率显著低于对照组,见图7B。以上结果均表明KIFC1敲除可使HeLa细胞增殖能力下降。
流式细胞术检测细胞周期分布结果显示,细胞周期调控发生显著的变化。其中,KIFC1敲除组G0/G1期和G2/M期细胞比例均显著高于对照组(P<0.05),而S期细胞比例则明显低于对照组,提示细胞DNA合成受到抑制,见图7C~G,进一步印证了KIFC1敲除导致HeLa细胞的增殖水平降低。
2.8 敲除KIFC1可促进HeLa细胞凋亡
KIFC1敲除组细胞经吖啶橙染色后,通过荧光显微镜可观察到不同程度的细胞凋亡情况,凋亡细胞核呈致密浓染的黄绿色,而对照组细胞的细胞核表现为均匀的绿色荧光,见图8A。

图8 KIFC1缺失对HeLa细胞凋亡的影响Figure 8 Effects of KIFC1 deletion on HeLa cell apoptosis
通过Annexin/FITC共标的流式细胞术检测进一步发现,敲除KIFC1后正常细胞比例减少(P<0.01; HeLa, 95.60±0.29%;KIFC1-/-Clone,93.23±0.12%),见图8B~D,早期凋亡细胞比例有所增加,见图8E,晚期凋亡细胞比例显著增加(P<0.05;HeLa,1.05±0.23%;KIFC1-/-Clone,2.67±0.28%),见图8F。综上结果表明,敲除KIFC1可促进HeLa细胞凋亡。
3 讨论
驱动蛋白是一类沿着微管运输分子的马达蛋白,在细胞器运输、细胞分裂、组织器官发育、信号转导等过程中发挥重要作用[14]。驱动蛋白的异常表达影响染色体的正确分布,引起有丝分裂过程中遗传物质变化和异常纺锤体形成,促进癌症的发生发展。近年来,驱动蛋白已经成为癌症药物开发的潜在靶点[15-18]。其中KIFC1是多种癌症诊断、治疗和预后的潜在生物标志物。KIFC1蛋白通过聚集中心体在癌细胞有丝分裂中发挥关键作用[19]。研究表明,KIFC1蛋白可直接结合CEP215,促进中心体与纺锤体极的连接,从而提高中心体聚集[20]。KIFC1还可能参与染色体排列、细胞周期中细胞器的运输,调节细胞周期G2-M期的进程。KIFC1的过表达驱动了控制有丝分裂检查点基因的过量表达,从而产生非整倍体,加速癌症发展的进程[21]。KIFC1蛋白参与多条肿瘤形成相关的信号通路。在子宫内膜癌中,KIFC1通过结合HMGA1促进c-myc的表达,从而加快c-myc介导的下游糖酵解基因的表达[22]。在肝癌中,KIFC1通过激活gankyrin/AKT/TWIST1通路,促进上皮间质转化和癌症转移[23]。KIFC1还可以通过Akt/GSK3β信号通路加速膀胱癌细胞增殖并诱导上皮间质转化[24]。在多种类型癌组织中,KIFC1的表达随着肿瘤发展级别的增加而增加,且KIFC1高表达易引起耐药性,与癌症患者的不良预后相关[25-28]。近期研究表明,DNA修复途径的ATM和ATR激酶诱导KIFC1-Ser26磷酸化,促进中心体聚集,导致耐药性和肿瘤复发[29]。这些研究揭示了KIFC1可能多方面参与癌症的发生发展过程。
目前KIFC1蛋白相关的研究通常使用基于RNAi机制的敲低细胞模型,我们利用CRISPR/Cas9技术在宫颈癌HeLa细胞中成功构建KIFC1敲除模型,可以更好地阐述KIFC1在宫颈癌中的作用机制。免疫荧光实验结果显示,在细胞间期,KIFC1大量定位在细胞核中,这与KIFC1尾部结构域的核定位序列(NLS)有关,KIFC1可与入核转运蛋白复合体中的Importin β结合,入核参与细胞周期的调控,并且其与核孔复合物NUP62相互作用可能介导核孔的重组[30]。在有丝分裂中后期,KIFC1与纺锤体共定位,表明KIFC1主要作用于纺锤体。在细胞分裂过程中,KIFC1参与微管交联、纺锤体两级形成、纺锤体组装和染色体分离等生命活动过程。之前的研究发现,KIFC1敲低可延长细胞的前中期,延缓细胞周期蛋白A的降解,进而破坏染色体凝集和排列,导致染色体错位、核型异常及微核的形成[31]。本研究观察到类似的结果,KIFC1基因的敲除可使细胞形态发生明显的改变,出现多核、微核等异常核结构,诱发微管骨架紊乱,还可以阻滞细胞周期,抑制DNA合成并诱导细胞凋亡,进而抑制HeLa细胞的增殖。这一结果提示作为宫颈癌治疗的潜在靶点,KIFC1具有广阔的发展前景。
综上所述,本研究以宫颈癌HeLa细胞系为研究模型,通过CRISPR/Cas9基因编辑技术敲除KIFC1基因,成功抑制KIFC1蛋白的表达。研究结果发现,KIFC1敲除可导致细胞骨架的异常装配,诱导细胞阻滞在G0/G1期、G2/M期,诱导HeLa细胞凋亡,从而减少细胞的增殖。未来,在KIFC1敲除细胞模型基础上可以结合临床实际问题如肿瘤治疗耐药性,深入研究KIFC1在宫颈癌中的潜在功能机制,为宫颈癌筛查、诊断和治疗提供新的思路。
利益冲突声明:
所有作者均声明不存在利益冲突。
猜你喜欢
中国医学科学院学报(2022年1期)2022-12-30 13:30:21
诊断学(理论与实践)(2020年2期)2020-06-28 12:41:12
国际呼吸杂志(2019年4期)2019-03-12 01:07:30
食品科学(2018年10期)2018-05-23 01:27:28
生物学教学(2017年9期)2017-08-20 13:22:32
中华老年多器官疾病杂志(2016年7期)2016-04-28 08:43:05
癌症进展(2016年10期)2016-03-20 13:15:43
西南医科大学学报(2015年1期)2015-08-22 13:01:46
医学研究杂志(2015年5期)2015-06-10 06:43:26
中国当代医药(2015年9期)2015-03-01 02:01:59
