
代谢工程改造大肠杆菌发酵生产β-烟酰胺单核苷酸
2023-12-14 12:45安俊侠王倩倩王昭颖徐庆阳范晓光
食品科学 2023年22期
安俊侠,王倩倩,王昭颖,刘 欢,徐庆阳,范晓光
(天津科技大学生物工程学院,工业发酵微生物教育部重点实验室,天津 300457)
β-烟酰胺单核苷酸(β-nicotinamide mononucleotide,β-NMN)属维生素B族衍生物,广泛存在于蔬菜、真菌、肉类和虾类等天然食物中,参与人体内多种生化反应,与免疫、代谢调控息息相关[1-2]。在人体中,β-NMN作为辅酶I NAD+的前体,直接参与NAD+的合成,并通过NAD+体现其功能[3-4]。多项研究发现,β-NMN对缓解老年退行性疾病、神经退行性疾病、代谢紊乱和衰老等方面具有较好的作用[5-6]。β-NMN的食品安全性较高,日本、欧美等国家已经批准其作为新食品原料[7-8]。与NAD+相比,β-NMN更容易进入细胞,因此适合在食品、饮料、保健品中使用。
根据2013年我国第五次国家卫生服务调查分析报告中的老年人生活照料调查统计[1]数据显示(以城市老年人生活照料分析为主),11.9%的在城市生活的老人在近30天的生活起居方面需要照顾时,49%由配偶照顾;47.4%由子女或孙子女照顾;0.8%由亲戚、朋友、邻居照顾;0.2%由社区提供照顾服务;1.8%是其他受助渠道获取照顾服务或者是没有人照顾,生活照顾需要自己独立完成,具体见表1。
目前,β-NMN的生产方法主要分为化学合成法和酶催化法。化学合成法工艺成熟,是β-NMN的主要生产方法,但存在合成路线繁琐、反应条件严苛、产物手性分离困难、使用有机溶剂等问题,导致产品售价较高[9-10]。酶催化法安全环保,产品旋光性单一,更容易被消费者接受[11]。酶催化法可以分为3 种路线,第1种是以烟酰胺核糖(nicotinamide riboside,NR)为底物,以ATP为磷酸供体,在烟酰胺核糖激酶的作用下生成β-NMN[12];第2种是以烟酰胺(nicotinamide,NAM)和5-磷酸核糖-1-焦磷酸(5-phosphoribosyl-1-pyrophosphate,PRPP)为底物,在烟酰胺磷酸核糖转移酶(nicotinamide phosphoribosyl transferase,Nampt)的作用下生成β-NMN[13];第3种是以烟酸和PRPP为底物,在烟酸磷酸核糖转移酶的作用下先生成烟酸单核苷酸(nicotinic acid mononucleotide,NaMN),然后在β-NMN合成酶的作用下生成β-NMN[14-15]。酶催化法的问题在于原料成本较高,酶活性和转化效率较低,因此限制了工业级应用放大[16]。
大部分微生物可以通过自身代谢合成PRPP,因此理论上通过系统代谢改造,可以实现以NAM或烟酸为前体发酵生产β-NMN[17]。本研究以生长周期较短、遗传背景清晰的野生型大肠杆菌(Escherichia coli)W3110作为出发菌株,设计模块化代谢改造策略(图1)。首先,分析并减弱底盘细胞中与NAM和β-NMN降解相关酶的表达,减少底盘细胞对前体和产物的额外消耗;其次,引入NAM输入蛋白(BcNiaP)、β-NMN输出蛋白(BmPnuC)、PRPP合成酶(PRPP synthetase,Prs)和Nampt,敲除调节蛋白PurR,实现了β-NMN的初步积累;此后,考察了不同来源的Nampt,筛选得到了酶活性较高且对底盘细胞负担较小的催化用酶;最后,强化β-NMN输出蛋白和Prs的表达水平,大幅提升了β-NMN的发酵产率。本研究通过基因组编辑和组成型质粒表达方式,构建1 株遗传背景清晰、无营养缺陷、无需诱导的大肠杆菌基因工程菌,以期实现NAM为前体发酵生产β-NMN。
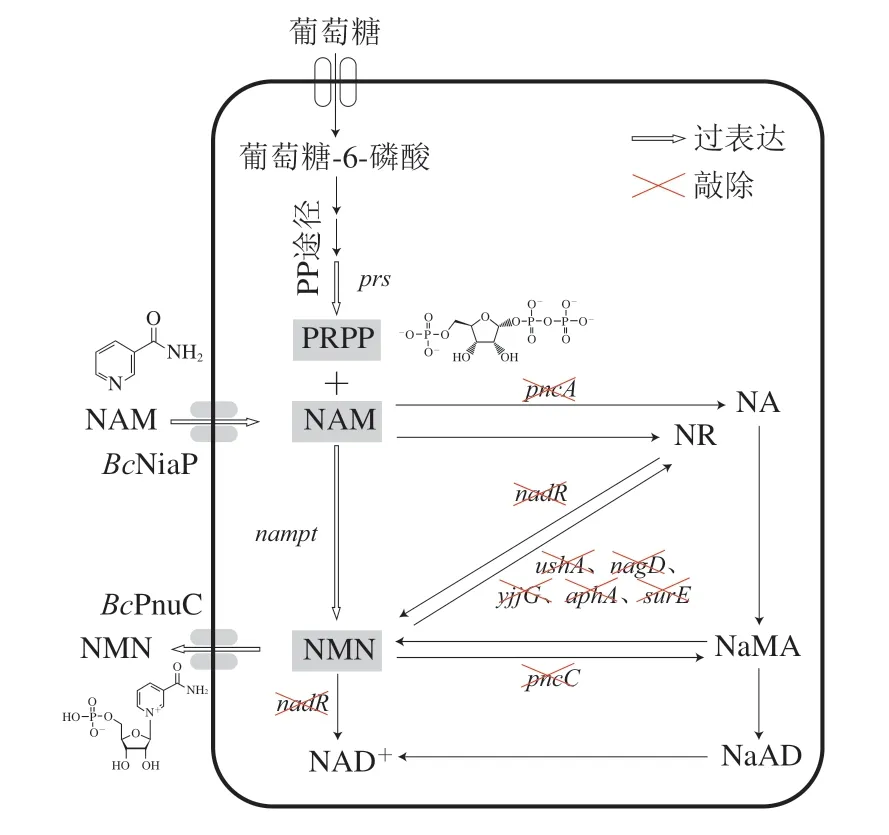
图1 代谢工程改造大肠杆菌发酵生产β-NMN策略Fig.1 Metabolic engineering strategies for β-NMN fermentation in E.coli
1 材料与方法
1.1 材料与试剂
1.1.1 菌株与质粒
综上所述,经过长期的发展,政府引导基金在撬动社会资本、政府意愿体现、政府参与公司治理能力和多阶段投资引导能力等方面为新兴产业发展进行了以FOF为代表的制度创新,但现有模式在考虑新兴产业融资的多阶段性、行业不对称性等方面仍有较大不足,推动新兴产业发展需要对融资特征进行准确定义,并在此基础上进一步构建创新融资模式。
本研究使用的菌株和质粒分别见表1、2。其中,E.coliDH5α用于质粒构建,E.coliW3110 ΔlacI作为出发菌株用于构建β-NMN生产菌株。
翻转课堂是常用的教学方法,高中物理实验教学过程中,也采用了翻转课堂的方法,这种教学方法相比于传统的教学方法有很多优点.首先,它把课堂的大多数时间都归还给学生,学生成为课堂的重心.翻转课堂的提出,改变了传统的授课模式,最大限度划分课堂时间,合理配置教学资源,更好地为学生服务.这种教学模式在高中物理实验中的应用,不仅提高了学生的学习效率,而且还促进了教育事业的改革.

表1 本研究所用菌株Table 1 Strains used in this study

表2 本研究所用质粒Table 2 Plasmids used in this study
1.1.3 引物
业务流程优化指中小出版企业通过自主研发和外购系统,实现采、编、印、版、售等技术创新,聚焦于出版产业全链数字化辅助技术,促进绿色印刷业务的开拓。“四维传媒”基于“云计算”,将数字出版技术与移动平台技术顺利嫁接,形成每个在线服务技术模块,实现远程创意、中央图库、在线编辑、远程批注、色彩管理、远程打样、绿色印刷以及在线全媒体阅读等全链整合,极大地提高了数字出版效率。截至2017年年底,“四维传媒”的“绿色出版”系统已经获得多项海外机构认证,并与欧美4个国家建立了战略合作伙伴关系。“龙源数媒”不断创新和迭代触控阅读和互联网技术,提高了产品“按需出版”的能力,为新增公共文化产品线预留了出版空间。
在细胞中实现β-NMN的发酵生产,需要考虑3 个方面的问题,一是前体NAM和产物β-NMN的转运;二是前体PRPP的供应;三是催化β-NMN合成关键酶Nampt的活力。Shoji等[27]发现Burkholderia cenocepacia来源的烟酸转运蛋白(BcNiaP)以及Bacillus mycoides来源的NR转运蛋白(BmPnuC)可以分别提高大肠杆菌对NAM的摄取和β-NMN的外排。BcNiaP和BmPnuC均为膜蛋白,表达过强将会影响菌体生长,因此将niapBc和pnuCBm基因整合至工程菌N-8基因组ilvG和ygaY位点上,并使用组成型Ptrc启动子控制其表达,获得工程菌N-9和N-10。
斜面培养基:葡萄糖5 g/L,蛋白胨10 g/L,酵母粉5 g/L,牛肉膏10 g/L,NaCl 5 g/L,琼脂20 g/L。
摇瓶种子培养基:葡萄糖20 g/L,蛋白胨3 g/L,酵母粉5 g/L,无水柠檬酸2 g/L,甲硫氨酸0.5 g/L,KH2PO42.5 g/L,MgSO4·7H2O 0.5 g/L,FeSO410 mg/L,VB1、VB3、VB5、VB12各1 mg/L,pH 7.0~7.5。
摇瓶发酵培养基:葡萄糖20 g/L,蛋白胨4 g/L,酵母粉6 g/L,无水柠檬酸2 g/L,甲硫氨酸0.3 g/L,KH2PO45.5 g/L,K2HPO45 g/L,MgSO4·7H2O 2 g/L,FeSO420 mg/L,VB1、VB3、VB5、VB12各2 mg/L,苯酚红8 mg/L,pH 7.0~7.5。
NAM 是β-NMN 合成的重要前体,大肠杆菌中pncA编码的烟酰胺酶可以催化NAM脱酰胺生成烟酸,是NAM 的主要支路代谢[19]。因此,首先敲除pncA基因,减少底盘细胞对NAM的额外消耗,构建出工程菌N-1。
1.1.2 培养基
引物由安升达(天津)生物科技有限公司合成。
1.1.4 试剂
蛋白胨、酵母粉 英国Oxoid公司;无水柠檬酸、甲硫氨酸、KH2PO4、K2HPO4、MgSO4·7H2O、FeSO4、(NH4)2SO4、NaCl 国药集团试剂有限公司;NAM、β-NMN 上海阿拉丁生化科技股份有限公司;乙腈(色谱纯)美国赛默飞世尔科技公司;基因组提取试剂盒、质粒提取试剂盒、胶回收试剂盒 美国Omega BioTek公司;限制性内切酶、Primer STAR HS DNA聚合酶 宝日医生物技术(北京)有限公司;单片段快速克隆试剂盒 南京诺唯赞生物科技有限公司。
1.2 仪器与设备
PTC-1148型聚合酶链式反应(polymerase chain reaction,PCR)仪 美国Bio-Rad公司;5 L自动控制发酵罐 上海保兴生物设备工程有限公司;U3000高效液相色谱仪、Hypersil™ ODS-2 C18色谱柱 美国赛默飞世尔科技公司;SBA-40D生物传感器分析仪 山东省科学院生物研究所。
1.3 方法
1.3.1 重组质粒构建
对于快递包装回收,国家有关部门制定了一个时间表。2017年,国家十部门联合发布的《关于协同推进快递业绿色包装工作的指导意见》明确,到2020年,可降解的绿色包装材料应用比例将提高到50%,基本淘汰重金属等特殊物质超标的包装物料,基本建成专门的快递包装物回收体系。主要快递品牌协议客户电子运单使用率达到90%以上,平均每件快递包装耗材减少10%以上,推广使用中转箱、笼车等设备,编织袋和胶带使用量进一步减少。基本建立快递业包装治理体系。2019年1月1日实施的《电商法》中第五十二条规定,快递物流服务提供者应当按照规定使用环保包装材料,实现包装材料的减量化和再利用。
本研究所用质粒pTrc99a自身含有表达乳糖操纵子阻遏物的lacI基因,在表达该质粒时,需添加异丙基硫代半乳糖苷(isopropyl-β-D-thiogalactoside,IPTG)进行诱导,但IPTG易影响菌体生长,因此构建组成型质粒QI-pTrc99a。通过设计两端携带酶切位点ApaI的引物,利用PCR扩增出不含lacI的线性化pTrc99a载体,再通过重组酶Exnase®II的作用使其自身环化,构建出组成型质粒QI-pTrc99a,该质粒构建示意图见图2。根据目的基因选择适宜的酶切位点,对质粒QI-pTrc99a进行双酶切,利用PCR扩增获得含酶切位点、启动子的目的基因片段,将酶切、PCR产物回收后利用Exnase®II重组,经转化、筛选、鉴定后获得重组质粒。

图2 组成型质粒QI-pTrc99a构建方法Fig.2 Flow chart for the construction of the constitutive plasmid QI-pTrc99a
1.3.2 基因组编辑
本研究利用规律成簇的间隔短回文重复/CRISPR相关蛋白9(clustered regularly interspaced short palindromic repeats/CRISPR-associated protein 9,CRISPR/Cas9)介导的基因编辑系统对菌株基因组进行改造[18]。CRISPR/Cas9基因编辑系统由pREDCas9和pGRB两个质粒构成。温敏型质粒pREDCas9主要由λ噬菌体的RED重组系统、Cas9蛋白表达系统和pGRB质粒消除系统组成,含奇霉素抗性(工作质量浓度100 mg/L,32 ℃)。质粒pGRB以质粒pUC18为骨架,由启动子J23100、gRNA-Cas9蛋白结合区域和终止子组成,含氨苄青霉素抗性(工作质量浓度100 mg/L,37 ℃),该质粒是针对某一靶位点利用gRNA设计工具CRISPR RGEN Tools(http://www.rgenome.net/cas-designer/)选定gRNA序列后设计出的重组质粒。pGRB能够转录出相应的gRNA,与pREDCas9表达出的Cas9蛋白结合后形成复合物并识别到靶位点,从而实现该靶位点的双链DNA断裂;此时,外源提供的根据待编辑基因设计出的重叠DNA片段(整合片段:上游同源臂-待整合基因-下游同源臂;敲除片段:上游同源臂-下游同源臂)可在重组酶作用下,与断裂的双链DNA发生同源重组,进而实现基因组改造。
通过紫外分光光度计(OD600nm)测定菌体生物量。通过SBA生物传感仪测定葡萄糖质量浓度。通过高效液相色谱仪检测β-NMN质量浓度,色谱条件:色谱柱为Hypersil™ ODS-2 C18(4.6 mm×250 mm),流动相为95% 20 mmol/L乙酸铵∶5%乙腈(V/V),柱温30 ℃,检测波长254 nm,流动相总流速1 mL/min。
1.3.3 摇瓶发酵
课堂教学中教师可以利用平台针对某个知识点或者一节课的所有知识点,做一次课堂检测,由于平台可以同步看到学生的答题情况,汇总统计答题的结果,这样教师就能够根据随堂练习的反馈结果及时把握学生的掌握情况,及时对学生进行分层式的个性化指导,也便于后面教学内容的调整。
重组菌株通过试管斜面培养基传代活化后,接种至30 mL种子培养基中(500 mL圆底瓶),37 ℃、200 r/min培养8~10 h后,按10%接种量转接至30 mL发酵培养基中(500 mL挡板瓶),37 ℃、200 r/min培养24 h。发酵期间根据酸碱指示剂苯酚红的变色情况,适当补加氨水或60 g/100 mL葡萄糖溶液。发酵培养基初始时添加0.1 g/L NAM,此后通过流加的方式保证发酵结束时共加入0.4 g/L NAM。
1.3.4 发酵罐发酵
重组菌株通过茄型瓶斜面培养基传代活化后,接种至2 L种子培养基(5 L发酵罐)中,培养期间通过流加氨水调节发酵液pH 7.0左右,37 ℃培养,维持溶氧在20%~40%之间,通风量在2~4 L/h之间,搅拌转速在200~700 r/min之间。当OD600nm达到10~20时,按15%的接种量接入3 L发酵培养基(5 L发酵罐)中,维持发酵液pH 7.0左右,37 ℃培养,溶氧在10%~35%之间,通风量在2~4 L/h之间,搅拌转速400~900 r/min,发酵周期38 h。当罐内的葡萄糖耗尽时,开始以一定速率流加80 g/100 mL的葡萄糖溶液,维持罐内葡萄糖质量浓度在0.1~5 g/L之间。发酵培养基初始时添加0.5 g/L NAM,此后以一定速率流加NAM,保证在发酵结束时共加入5 g/L NAM。
将超声技术应用于神经阻滞进程中,能促使周围神经可视化,进而促使神经阻滞的有效率相应提升。当下,超声技术在神经阻滞麻醉应用范畴不断拓展,其具有二维分辨率高的优势,多普勒效应能检测出低血流信号,超声技术联合神经刺激仪进行神经阻滞,能够协助麻醉医生全过程均能看到针的移动情况,观察麻醉的散布情况,有助于明显提升阻滞神经定位的精确性,对血管、神经基本不产生影响,术前准备时间较为短暂。对于股神经,闭孔神经等位置相对较浅的神经,若单独使用神经刺激仪,那么神经具体位置确认难度较大、阻滞成功率较低、术前准备时间较长、神经阻滞见效时间较长且并发症发生率较高[2]。
1.3.5 检测与分析
滑坡稳定性计算及滑坡推力计算,其目的是为滑坡在不同工况条件下的稳定性评价及滑坡防治提供设计依据。计算荷载考虑滑体自重、地表荷载、暴雨、动荷载、地震等因素。按《滑坡防治工程设计与施工技术规范》(DZ/T0219-2006),滑坡治理根据其危害对象程度及潜在经济损失,滑坡稳定性计算工况、荷载组合及抗滑稳定安全系数见表3。
在重组菌株构建时,同时将pGRB质粒和重叠DNA片段电转至已导入pREDCas9质粒的感受态细胞中,复苏2 h后涂布于含100 mg/L氨苄青霉素和奇霉素的LB固体培养基上,挑选单菌落进行菌落PCR筛选阳性转化子。将阳性转化子转接至含10 mmol/L阿拉伯糖和奇霉素的LB液体培养基中,诱导pREDCas9中的pGRB质粒消除系统表达,通过菌落PCR筛选已丢失pGRB的阳性菌株转接至LB液体培养基中,42 ℃培养12 h,消除温敏性质粒pREDCas9,最终经过菌落PCR筛选出无质粒的重组菌株。
1.4 数据处理
使用Origin 2019对数据进行处理和作图分析,使用Adobe Photoshop 2020和Visio 2019对图片进行处理组合和绘制。
2 结果与分析
2.1 减少底盘细胞对前体和产物的额外消耗
发酵罐发酵培养基:葡萄糖20 g/L,蛋白胨4 g/L,酵母粉6 g/L,无水柠檬酸2 g/L,甲硫氨酸0.3 g/L,KH2PO41 g/L,K2HPO45 g/L,MgSO4·7H2O 2 g/L,FeSO420 mg/L,(NH4)2SO42 g/L,VB1、VB3、VB5、VB12各2 mg/L,微量元素2 mL/L,pH 7.0~7.5。
目标产物β-NMN作为NAD+的直接前体物,在大肠杆菌中的代谢较为活跃,因此需要对其支路代谢途径进行改造。β-NMN的支路代谢主要分为3 种(表3),一是通过pncC编码的烟酰胺核苷酸酰胺水解酶转化为NaMN[20];二是通过nadR编码的烟酰胺核苷酸腺苷酰转移酶转化为NAD+[21];三是通过一系列的核苷酸酶降解为NR。根据文献报道,与β-NMN降解相关核苷酸酶基因包括ushA、aphA、nagD、yjjG、surE[22-26]等。
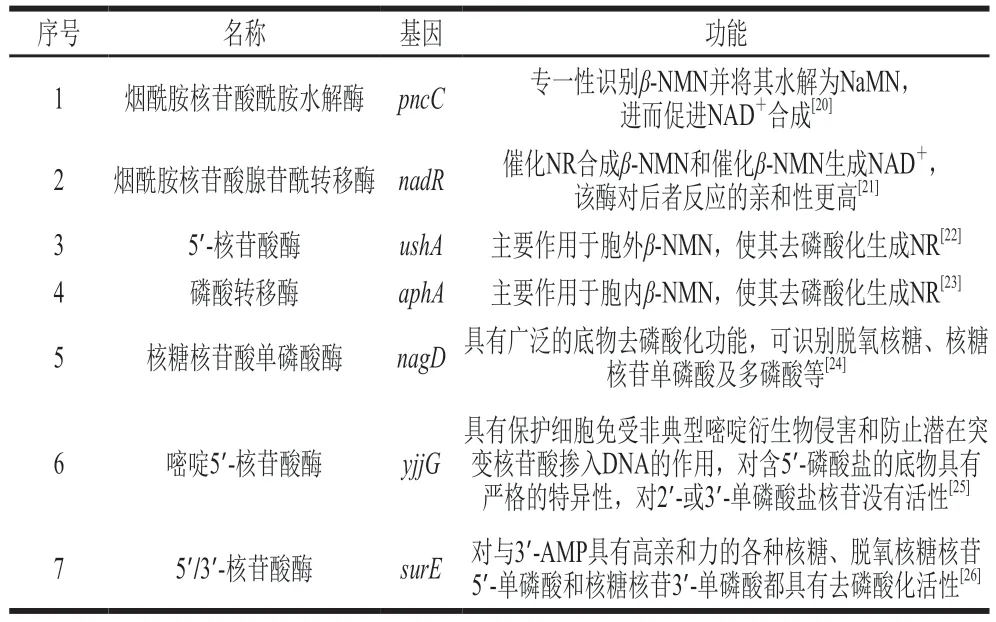
表3 大肠杆菌中β-NMN的主要支路代谢Table 3 Major β-NMN shunt metabolic pathways in E.coli
在工程菌N-1的基础上,对β-NMN支路代谢途径中的酶进行逐一敲除,获得工程菌N-2~N-8。将上述菌株放到含有0.5 g/Lβ-NMN的摇瓶发酵培养基中培养16 h,考察改造后细胞对产物的消耗情况,结果如图3所示。敲除pncC和nadR基因并不会减少细胞对β-NMN的降解,说明β-NMN向NaMN和NAD+的支路代谢并不活跃。敲除ushA后,底盘细胞对β-NMN的消耗量从100%显著降低至25.16%,说明大肠杆菌内β-NMN向NR的转化是β-NMN的主要支路代谢。继续敲除其他核苷酸酶基因aphA、nagD、yjjG和surE后,底盘细胞对β-NMN的消耗量持续降低。最终,工程菌N-8对产物β-NMN的消耗量仅为4.57%,生物量与N-1相比下降了8.84%。通过上述一系列的基因改造,获得了能够用于β-NMN积累的优良底盘。
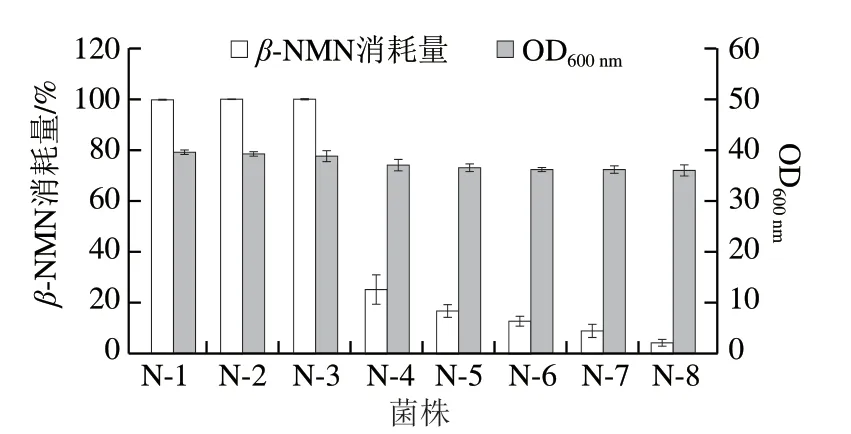
图3 底盘细胞改造对产物β-NMN消耗以及菌株生长的影响Fig.3 Effect of chassis cell modification on β-NMN consumption and cell growth
2.2 构建β-NMN的合成代谢途径
LB培养基:蛋白胨10 g/L,酵母粉5 g/L,NaCl 10 g/L。
弹幕(Barrage)顾名思义是子弹多而形成的幕布,在军事上是指大量密集的炮火攻击。在弹幕视频中指的是大量的评论在屏幕上划过,像是幕布一样。最早也是最著名的弹幕网站是日本的Niconico动画网站,其最大特点是实时弹幕发送,发送后3s就显示在屏幕上,使得观众有一起参与的感觉,取得了空前成功。国内先引进弹幕技术的是Acfun弹幕视频网和Bilibili动漫网站,随后主流视频网站如土豆、爱奇艺和优酷等也相继引入弹幕技术。这种实时交流,产生共鸣的体验感,越来越受到各个领域的广泛关注。
PRPP是β-NMN合成的重要前体,主要是由Prs催化ATP上的焦磷酸基转移到5-磷酸核糖的1-羟基位而产生[28]。为了强化大肠杆菌中PRPP的合成代谢,首先将内源的prsA基因整合至工程菌N-10基因组gapC位点上,并使用其内源Pprs启动子控制其表达,获得工程菌N-11。调节基因purR对大肠杆菌嘌呤生物合成途径所有结构基因的表达起着负调控作用[29]。因此本研究敲除工程菌N-11的purR基因以促进prsA基因的表达,获得工程菌N-12。
在上述代谢改造的基础上,将使用组成型质粒(1.3.1节)表达Chitinphaga pinensis来源的Nampt,保证关键酶的拷贝数。将重组质粒QI-pTrc99a-namptCp分别转化至N-9~N-12菌株,获得工程菌N-9’~N-12’,并对其进行摇瓶发酵测试。发酵过程中添加0.5 g/L前体NAM,发酵周期24 h,结果如图4所示。引入BcNiaP菌株N-9’的发酵液中只检测到0.01 g/L的β-NMN,而引入BmPnuC后,菌株N-10’能够产生0.16 g/L的β-NMN,说明大肠杆菌自身并不存在β-NMN外排系统,必须依赖外源转运蛋白BmPnuC才能实现产物的胞外积累。强化prs基因表达以及敲除purR基因能够使β-NMN的产量提高1.1 倍(0.34 g/L),说明上述基因改造能够有效提高PRPP供应,为β-NMN合成提供更充足的前体。但是,prs基因过表达会导致生物量下降7.8%。
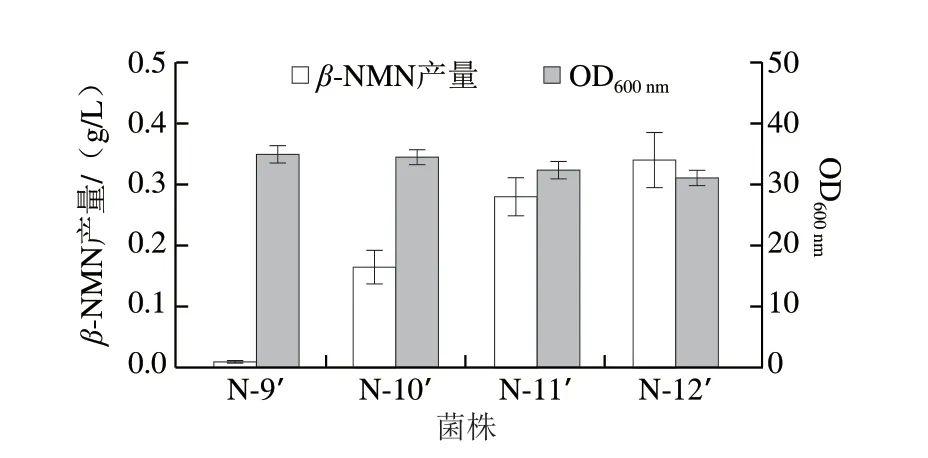
图4 β-NMN的合成代谢途径构建对菌株生长和β-NMN发酵的影响Fig.4 Effect of the construction of β-NMN anabolic pathway on cell growth and β-NMN fermentation
2.3 筛选高活性的Nampt
Nampt是β-NMN合成的限速酶,筛选高活性的Nampt是提升β-NMN发酵产量的关键。根据文献报道,挑选5 个能够在大肠杆菌中表达的Nampt,分别来自于Chitinphaga pinensis、Comamonadaceae bacterium、Meiothermus ruber、Haemophilus ducreyi和Vibriobacteriophage KVP40[30-31]。将不同来源的nampt基因分别与组成型质粒(1.3.1节)QI-pTrc99a连接,构建出重组质粒转化至N-12菌株,获得工程菌N-12′~N-16,并对其进行摇瓶发酵测试。发酵过程中添加0.5 g/L前体 NAM,发酵周期24 h,结果如图5所示。引入VpNampt菌株N-16的β-NMN产量最高(0.90 g/L),但生物量最低,这可能与该酶是噬菌体来源有关。与之相比,引入CbNampt菌株N-13的β-NMN产量达到0.88 g/L,但生物量与N-16相比提高了1.3 倍。廖一波[32]利用分子对接手段,测定了不同来源Nampt与NAM的对接效果,发现CbNampt和MrNampt是细菌来源中评分最高的,但CbNampt的kcat/km值更高,达到9.86×106s-1。研究进一步证实,Comamonadaceae bacterium来源的Nampt在大肠杆菌中的作用效果最好,能够在保证细胞生长的同时提升β-NMN的发酵产量。
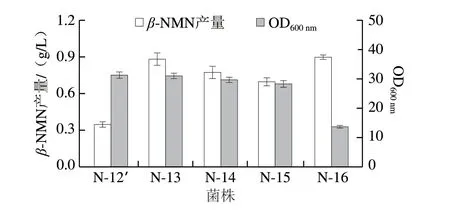
图5 不同来源Nampt对菌株生长和β-NMN发酵的影响Fig.5 Effects of Nampts from different sources on cell growth and β-NMN fermentation
2.4 加强PRPP供应及产物外排对β-NMN的影响
根据化学计量方程,在摇瓶发酵中添加0.5 g/L前体NAM,理论上能够获得1.37 g/L的β-NMN。但N-13菌株摇瓶发酵只产生了0.88 g/L的β-NMN,为理论转化率的64.2%。菌株N-13中关键酶Nampt是以质粒形式表达的,而Prs和BmPnuC是以基因组整合方式表达,因此推测可能是前体PRPP供应或产物外排不足限制了β-NMN的合成。在质粒Z-2的基础上,构建了namptCb-prsA双基因串联质粒Z-6,以及namptCb-prsA-pnuCBm三基因串联质粒Z-7,并将其分别转化至N-12菌株,获得工程菌N-17和N-18。对上述菌株进行摇瓶发酵测试,发酵过程中添加0.5 g/L前体 NAM,发酵周期24 h,结果如图6所示。强化prsA基因表达后,菌株N-17的β-NMN产量比N-13提高了44.3%(1.27 g/L),但生物量下降了55.1%。强化pnuCBm基因表达后,菌株N-18的β-NMN产量进一步提高,达到1.36 g/L,接近理论转化率,但生物量比N-13下降了48.7%。发酵结果表明,加强PRPP供应及产物外排能够有效提高β-NMN的发酵产量,但相关基因的表达强度需要严格控制,减少其对细胞生长的负面影响。

图6 加强PRPP供应及产物外排对菌株生长和β-NMN发酵的影响Fig.6 Effect of strengthening of PRPP supply and product efflux on cell growth and β-NMN fermentation
2.5 工程菌N-18分批补料发酵生产β-NMN
为了测试菌株N-18的发酵性能,在5 L发酵罐中进行分批补料发酵实验。发酵过程中添加5 g/L前体NAM,结果如图7所示。4~26 h,随着菌体生长,β-NMN产量不断增加,并保持较高的合成速率。当菌体生长进入稳定期后,随着发酵时间延长,菌体的摄糖速率变慢,β-NMN的合成速率有所降低。发酵36 h,β-NMN产量达到10.2 g/L,NAM到β-NMN的摩尔转化率为74.5%。此外,发酵液中存在1.2 g/L的副产物烟酸,说明除了PncA以外,大肠杆菌内可能存在未知的烟酰胺酶。
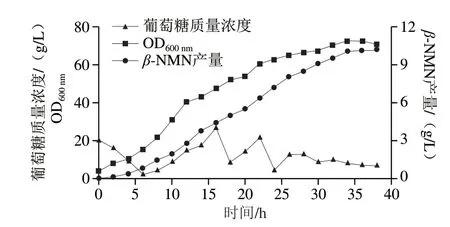
图7 工程菌N-18在5 L发酵罐分批补料发酵曲线Fig.7 Time course of fed-batch fermentation of the engineered strain N-18 in a 5 L bioreactor
目前,已有一些发酵法生产β-NMN的报道,发酵方式主要为全细胞催化或者流加NAM发酵,但都需要在产酶或者发酵阶段添加诱导剂(表4)。Huang Zhongshi等[31]证明噬菌体来源的VpNampt效果最好,强化PRPP供给和产物外排后,工程菌能够发酵产生16.2 g/L的β-NMN,NAM到β-NMN的摩尔转化率为97%,均为已有报道的最高水平。对比了不同来源的Nampt,发现细菌来源的CbNampt效果与VpNampt相差不大,且对菌体的生长负担较小。在5 L发酵罐补料发酵过程中,无需添加诱导剂即可实现β-NMN发酵生产,菌体生物量较高。但是,由于副产物烟酸的产生,限制了底物向β-NMN的转化。值得注意的是,在摇瓶发酵中并没有检测到烟酸积累,烟酰胺几乎完全转化为β-NMN。但在分批补料发酵过程中,当菌体浓度较高时,烟酸开始形成,β-NMN生产速率显著下降。根据上述现象,后续可以从代谢工程和发酵工程角度提升工程菌的发酵水平:一是通过启动子和RBS序列优化Nampt、Prs和BmPnuC的表达水平,并利用生物信息学筛选未知的烟酰胺酶;二是调整底物流加方式和流加时间,防止过量补加NAM;三是调整补糖速率和溶氧水平,防止菌体的过度生长,减少副产物的产生。
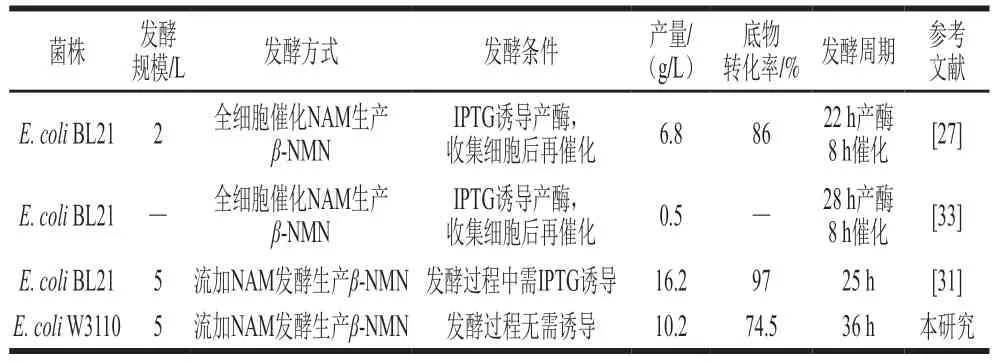
表4 不同β-NMN发酵方法对比Table 4 Comparison of β-NMN production using different fermentation methods
3 结论
研究以野生型大肠杆菌为底盘细胞,通过系统代谢工程改造,实现了β-NMN的发酵生产。主要策略包括:1)对NAM和β-NMN的支路代谢涉及的8 个酶进行失活,减少底盘细胞对前体和产物的额外消耗;2)引入外源NAM和β-NMN的转运蛋白以及Nampt,增强胞内PRPP供给,实现了β-NMN的初步积累(0.34 g/L);3)比较筛选了酶活力较高且对底盘细胞生长负担较小的Nampt,通过进一步优化PRPP供应和产物外排,β-NMN的摇瓶发酵产量提高至1.36 g/L。工程菌N-18于5 L发酵罐中发酵38 h,β-NMN产量可达10.2 g/L,NAM到β-NMN的摩尔转化率为74.5%。研究构建的β-NMN发酵菌株具有遗传背景清晰、无营养缺陷、无需诱导等优势,可为β-NMN的工业化发酵生产及菌株改良提供理论依据。
猜你喜欢
中国环境科学(2023年8期)2023-08-29
天津医科大学学报(2021年1期)2021-12-05
湖北农业科学(2017年8期)2017-05-26
广东饲料(2016年1期)2016-12-01
现代检验医学杂志(2016年5期)2016-08-20
大陆桥视野·下(2016年6期)2016-08-06
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
药学与临床研究(2014年3期)2014-03-06
无机化学学报(2014年6期)2014-02-28
茶叶通讯(2014年2期)2014-02-27
