
利用CRISPR/Cas9 技术创制小麦耐低肥Taaap3 突变体
2023-12-11 10:03陈艳艳方宇辉王玉民齐学礼
河南农业科学 2023年11期
李 艳,陈艳艳,华 夏,方宇辉,王玉民,巩 晨,齐学礼,胡 琳
(1.河南省作物分子育种研究院/河南省小麦生物学重点实验室/河南省麦类种质资源创新与改良重点实验室,河南郑州 450002;2.神农种业实验室,河南 郑州 450002)
氮素在植物光合作用中起着关键作用,氮素的吸收利用程度与植物生长发育状况密切相关。无机氮主要以硝酸盐和铵的形式被植物吸收,然后直接在根中或转移到叶子后转化为氨基酸[1],氨基酸被输送到根、叶、花和籽粒中[2]。氨基酸转运蛋白(Amino acid transporters,AAT)是位于生物膜上吸收及转运氨基酸的蛋白质,是植物代谢过程中的关键调控因子[3-6],对植物氮素的吸收和转运具有重要贡献[7]。氨基酸透性酶家族(Amino acid permeases,AAPs)是氨基酸转运蛋白超家族(Amino acid transporter family,ATF)中的亚家族,在氮汇和氮供应氨基酸的装载中起着重要作用[8]。
AAPs 基因最早在拟南芥(Arabidopsis thaliana)中被发现,拟南芥中有8 个AAPs 基因(AtAAP1—AtAAP8),AtAAPs 基因优先在维管组织中表达,负责酸性、中性及碱性氨基酸的长距离运输[7]。AtAAP1基因在根尖和根毛的表皮细胞中表达,在土壤中介导谷氨酸和中性氨基酸的吸收转运[9-10]。AtAAP2基因定位在韧皮部,能将叶片中的氨基酸装载到韧皮部进而转运至种子,对氨基酸在源库不同器官中的分配发挥重要作用[11]。AtAAP3和AtAAP5基因主要在根部表达,介导碱性氨基酸的吸收[7]。AtAAP6基因主要在木质部薄壁细胞中表达,调节韧皮部的氨基酸组成[12]。AtAAP8基因主要在未成熟的种子和成熟角果中表达,定位于质膜,负责将氨基酸运输到胚乳中供种胚发育[13-14]。OsAAP6基因显著增强根系对一系列氨基酸的吸收,并影响各种氨基酸的分布[3]。OsAAP4基因过表达可以提高转基因水稻植株中性氨基酸浓度,增加分蘖数,进而提高籽粒产量[15]。OsAAP3基因主要转运碱性氨基酸赖氨酸和精氨酸[16],在水稻中抑制OsAAP3基因表达,可以降低赖氨酸、精氨酸、组氨酸、天冬氨酸、丙氨酸、谷氨酰胺、甘氨酸、苏氨酸和酪氨酸浓度,促进发芽,增加分蘖数,进而提高籽粒产量[1]。小麦(Triticum aestivumL.)中关于TaAAPs 基因功能的研究报道甚少,仅有研究发现过表达TaAAP3基因可提高转基因小麦的耐盐能力[17],关于其耐低肥方面的研究尚未见报道。为此,以生产上大面积推广的小麦品种郑麦1860、郑麦1342 和郑麦7698 为背景材料,利用CRISPR/Cas9 技术敲除TaAAP3基因,创制小麦Taaap3突变体,为耐低肥小麦新品种的培育奠定基础。
1 材料和方法
1.1 试验材料
试验于2022—2023 年在河南省农业科学院现代农业科技试验示范基地(新乡市原阳县,113°42′4″E、35°0′17″N)进行。供试小麦品种为河南省作物分子育种研究院小麦分子育种团队选育的郑麦1860、郑麦1342 和郑麦7698。基因编辑载体TaU6-sgRNA 和Cas9基因载体pJIT163-ubirCas9由北京吉诺沃公司提供;限制性内切酶BstⅪ和Q5 PCR 扩增酶均购自NEB 有限公司;引物合成及测序由英潍捷基(上海)贸易有限公司完成。
1.2 基因编辑载体的构建
根据水稻OsAAP3基因(LOC_Os06g36180)序列,通过Gramene 网站(https://archive.gramene.org/)BLAST 获得TaAAP3基因A、B、D 亚基因组DNA 和cDNA 序 列(Triticum_aestivum_TRIAE_CS42_7AL_TGACv1_557545_AA1782830、Triticum_aestivum_TRIAE_CS42_7BL_TGACv1_576825_AA1856230、Triticum_aestivum_TRIAE_CS42_U_TGACv1_640866_AA2077760),利 用 网 站http://crispor.tefor.net/在TaAAP3基因编码区设计4 个sgRNA 引物sgRNA1—sgRNA4(图1),引物序列见表1。在sgRNA 正向引物5′端均加上黏性末端CTTG,在反向引物5′端均加上黏性末端AAAC。将4 个sgRNA 引物的sgRNA-F 和sgRNA-R 分别退火形成有黏性末端的双链DNA,将该双链DNA 连接到经BpiⅠ酶切的TaU6-sgRNA 载体上,转化大肠杆菌后,挑取单克隆,利用引物M13F/sgRNA-R 进行菌落PCR 扩增,测序无误后,编辑载体便构建完成。将构建好的4个TaU6-sgRNA 载体分别与pJIT163-ubi-rCas9载体共转化小麦原生质体,转化方法按照小麦原生质体制备及转化试剂盒(北京酷来搏科技有限公司)说明书进行,23 ℃黑暗条件下培养2 d后,提取原生质体DNA,sgRNA 活性检测采用突变位点检测通用引物(TaAAP3-T)进行PCR 扩增,对PCR 扩增出的700 bp 目的片段进行BstⅪ酶切,若未被BstⅪ限制性内切酶切开,则证明sgRNA 有活性,可用于小麦遗传转化。

表1 引物信息Tab.1 Information of primers
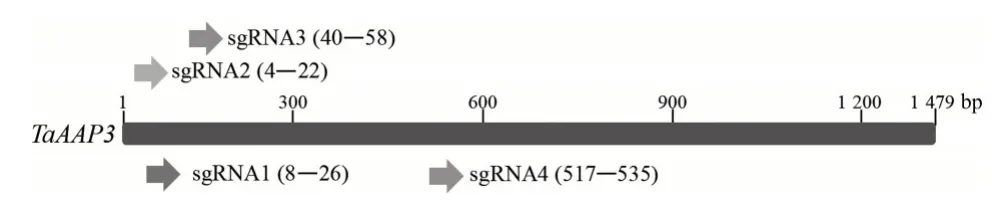
图1 设计的sgRNA靶向TaAAP3基因编码区的位置Fig.1 The location of the designed sgRNAs targeting the coding sequence of TaAAP3 gene
1.3 遗传转化
在小麦开花后12~14 d,取穗中部、大小一致的未成熟种子,用70%乙醇表面消毒1 min,无菌水冲洗3 次,然后用0.1%HgCl2消毒6 min,无菌水冲洗3~4 次。在超净工作台上剥出幼胚,接种于诱导培养基(MS+2 mg/L 2,4-D+4 g/L 琼脂+30 g/L 蔗糖,pH值6.2)上,培养1 d 后用于遗传转化。利用基因枪介导法将构建好的TaU6-sgRNA 与pJIT163-ubirCas9共转化小麦愈伤组织,具体转化方法参照HU等[18]的方法并稍作改进。经基因枪轰击后的愈伤组织在高渗培养基(MS+2 mg/L 2,4-D+7 g/L 琼脂+0.4 mol/L 甘 露 醇+30 g/L 蔗 糖,pH 值5.8)上 培 养16 h,转至诱导培养基,经过诱导、分化和生根,获得再生植株。待再生苗根系较健壮时,移载入花盆中,放置于温室中培养,获得T0植株。提取T0植株的基因组DNA,用突变位点检测通用引物(TaAAP3-T)进 行PCR 扩 增,对PCR 扩 增 出 的700 bp 目的片段进行BstⅪ酶切,目的片段未被BstⅪ限制性内切酶切开的植株即为T0突变植株。T0突变植株自交获得T1种子。
1.4 田间肥力试验设计
采用T3Tappp3突变体和野生型进行田间试验,设置正常和低肥2种处理,其中,正常处理基础肥力为有机质11.32 g/kg、全氮0.76 g/kg、碱解氮71.84 mg/kg、有效磷53.51 mg/kg、速效钾212.43 mg/kg,施肥量为尿素(含N 46%)493.65 kg/hm2、磷酸二铵(含N 17%、P2O540%)89.25 kg/hm2;低肥处理基础肥力为有机质11.98 g/kg、全氮0.64 g/kg、碱解氮65.52 mg/kg、有效磷22.39 mg/kg、速效钾149.60 mg/kg,施肥量为尿素257.55 kg/hm2、磷酸二铵115.5 kg/hm2。每个处理设置3次重复,每个重复0.3 hm2,正常田间管理。成熟时,收获整个小区小麦用于测产。
1.5 TaAAP3基因靶点序列突变检测
提取T2幼苗叶片DNA,先用TaAAP3基因靶点序列的通用引物(TaAAP3-T)进行PCR 扩增,然后用限制性内切酶BstⅪ对PCR 扩增产物进行酶切,筛选突变植株。分别采用含有TaAAP3基因靶点序列的A、B、D 亚基因组特异引物对获得的突变体进行PCR 扩增、测序,利用BioXM 2.6 软件比对分析突变位点。所用引物序列见表1。
1.6 TaAAP3基因表达量分析
采用植物RNA 提取试剂盒(天根生物科技有限公司)提取T2Taaap3突变体和野生型叶片总RNA。采用反转录试剂盒(Promega公司)将RNA 反转录为cDNA,使 用ChamQ Universal SYBR qPCR Master Mix Q711(诺唯赞生物有限公司)在Bio-Rad iQ5 上进行实时荧光定量PCR(qRT-PCR)扩增。用TaAAP3基因靶点序列的通用引物扩增TaAAP3基因,以小麦Actin基因为内参基因,引物序列见表1。扩增程序:95 ℃预变性30 s;95 ℃变性10 s,60 ℃复性30 s,40 个循环。所有试验样品均3 次重复。采用2-ΔΔCt方法分析基因的相对表达量。
1.7 氨基酸序列比对分析
利用Geneious 软件进行蛋白质氨基酸序列比对分析。
1.8 数据分析
使用Excel 2010 进行数据整理,采用SPSS 22.0进行差异显著性分析。
2 结果与分析
2.1 TaAAP3基因编辑载体的构建
将4 个靶标TaAAP3基因编码区的sgRNA 引物sgRNA-F 和sgRNA-R 分别退火形成双链DNA,将该双链DNA 连接到经BpiⅠ酶切的TaU6-sgRNA 载体上,转化大肠杆菌,进行菌落PCR 扩增,电泳结果显示在400 bp 处出现目的条带(图2)。对菌落PCR扩增阳性的菌斑进行测序,结果显示4 个sgRNA 引物均连入TaU6-sgRNA 载体(图3),经原生质体活性检测后确定sgRNA4(517—535 bp)为靶点序列,536—538 bp 作 为PAM(Protospacer adjacent motif)序列,将其构建好的载体命名为TaU6-AAP3-sgRNA,用于小麦遗传转化。
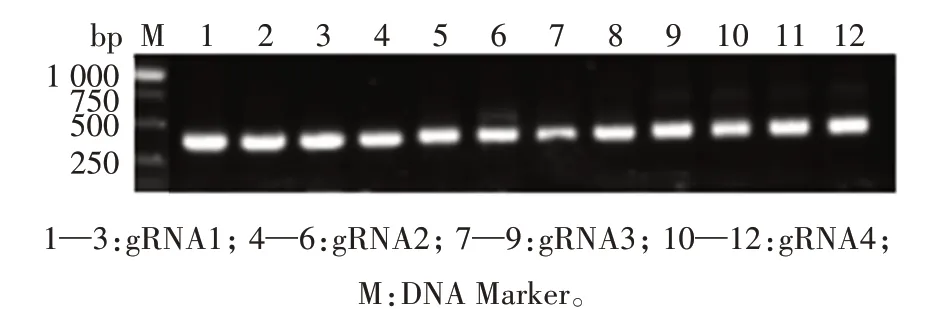
图2 4个基因编辑载体的单克隆菌落PCR检测结果Fig.2 Monoclonal PCR detection results of four gene editing vectors

图3 4个基因编辑载体的阳性单克隆测序结果Fig.3 Positive monoclonal sequencing results of four gene editing vectors
2.2 不同遗传背景下Taaap3 突变体的获得及TaAAP3基因靶点序列突变类型分析
以郑麦1860、郑麦1342 和郑麦7698 为受体材料,利用基因枪介导法将TaU6-AAP3-sgRNA 与pJIT163-ubi-rCas9载体共转化小麦愈伤组织,获得再生植株。经过PCR 扩增和酶切验证共筛选获得3个突变体,郑麦1860、郑麦1342 和郑麦7698 各获得1 个 突 变 体,分 别 为Taaap3-1、Taaap3-2 和Taaap3-3(图4—图5)。分别采用A、B、D 亚基因组靶位点检测引物通过PCR 扩增和测序分析T2突变体的突变类型。测序结果显示(图6),突变体Taaap3-1 为三突变类型,A、B、D 3 个亚基因组均为纯合突变,A 亚基因组在靶点处有26 bp 缺失,B 亚基因组插入1 bp,D 亚基因组插入51 bp;突变体Taaap3-2也为三突变类型,A、B、D亚基因组亦均为纯合突变,A 亚基因组在靶点处插入1 bp,B 亚基因组缺失23 bp,D亚基因组插入1 bp;突变体Taaap3-3为单突变类型,仅A 亚基因组发生突变,插入2 bp碱基,B和D亚基因组均未突变。
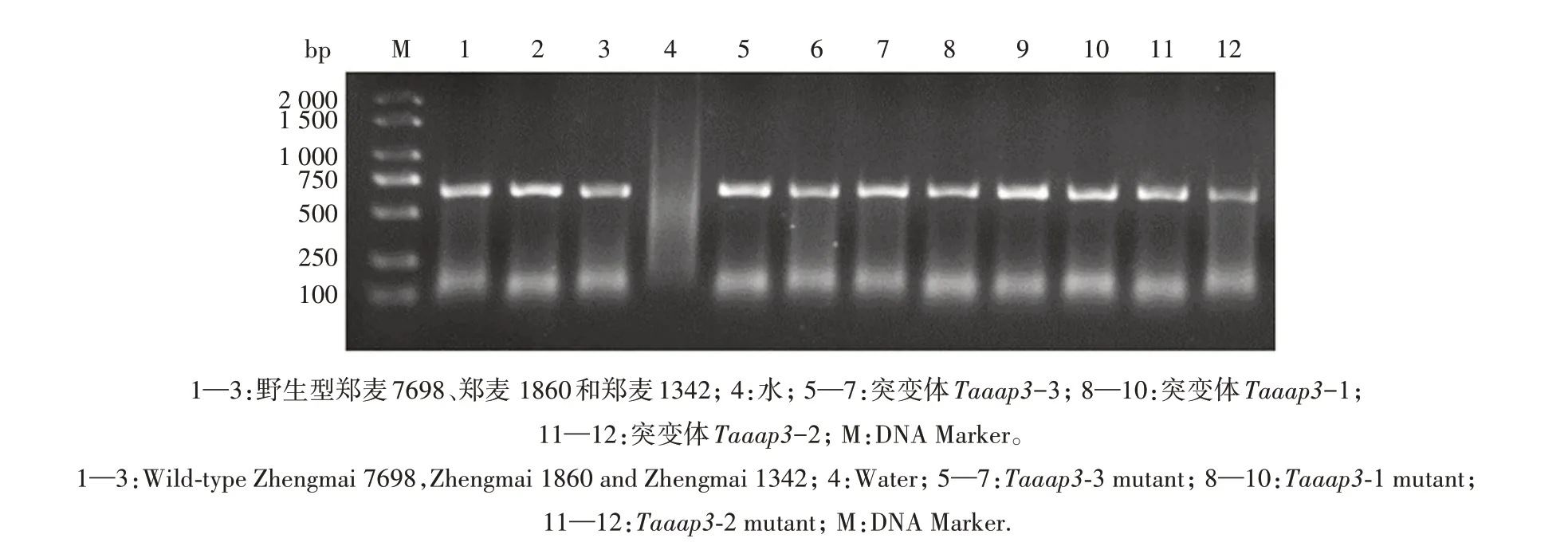
图4 Taaap3突变体的PCR检测结果Fig.4 PCR detection results of Taaap3 mutants
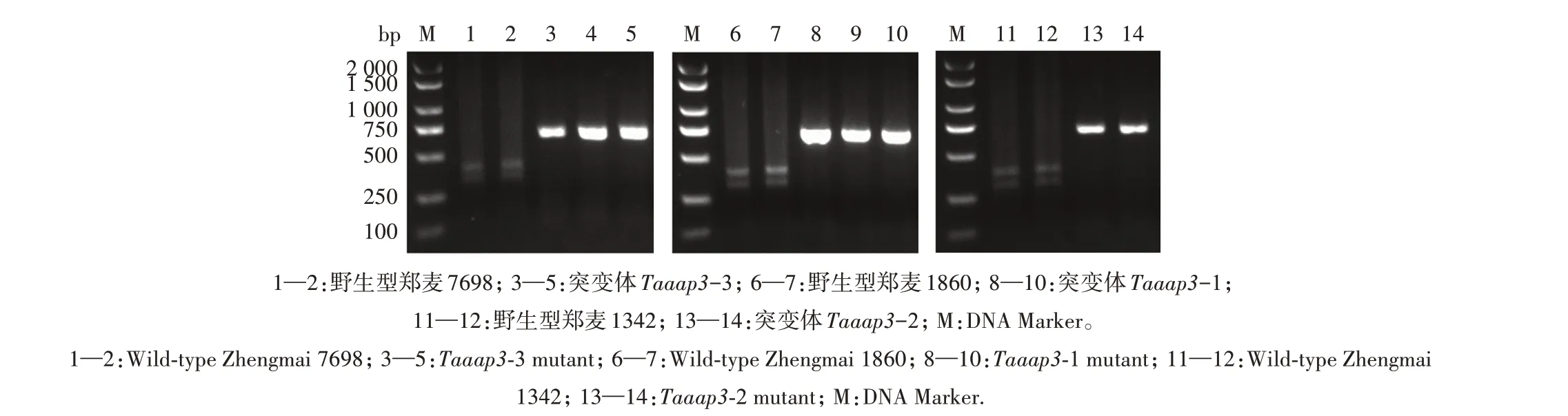
图5 突变体Taaap3的Bst Ⅺ酶切结果Fig.5 Bst Ⅺdigestion results of Taaap3 mutants
2.3 Taaap3突变体编码蛋白质的氨基酸序列分析
对Taaap3突变体编码蛋白质的氨基酸序列进行分析发现(图7),与野生型TaAAP3蛋白氨基酸序列相比,突变体Taaap3-1 A 亚基因组自第174 个氨基酸开始发生突变,之后的氨基酸序列完全被改变;B 亚基因组和D 亚基因组均在第178 个氨基酸处发生突变,并产生移码突变,导致翻译提前终止,编码氨基酸数由野生型的490 个分别缩短为177 个和182 个。突变体Taaap3-2 A、D 亚基因组均从第178 个氨基酸开始发生突变,B 亚基因组自第175 个氨基酸开始发生突变,之后的氨基酸序列完全被改变。突变体Taaap3-1和Taaap3-2 A、B、D亚基因组突变均导致蛋白质结构域发生改变,可能使TaAAP3 蛋白丧失了跨膜转运的活性。突变体Taaap3-3 A亚基因组自第178个氨基酸开始发生突变,之后的氨基酸序列亦完全被改变;B亚基因组和D亚基因组均未发生突变。
2.4 Taaap3突变体中TaAAP3基因的表达分析
TaAAP3基因核苷酸序列和表达水平在郑麦1860、郑麦1342 和郑麦7698 中无明显差异。因此,以郑麦1860 为对照,利用qRT-PCR 方法分析突变体Taaap3-1、Taaap3-2 和Taaap3-3 幼 苗 叶 片 中TaAAP3基因表达情况。结果显示(图8),3 个突变体幼苗叶片中TaAAP3基因表达水平均极显著低于野生型;突变体Taaap3-1 和Taaap3-2 中TaAAP3基因表达水平极显著低于Taaap3-3。说明TaAAP3基因A、B、D 亚基因组碱基的缺失和插入极显著降低了TaAAP3基因的表达量,且三突变类型突变体的TaAAP3基因表达水平极显著低于单突变类型。因此,通过CRISPR/Cas9 技术已成功对郑麦1860、郑麦1342 和郑麦7698 中TaAAP3基因进行了编辑,碱基的插入、缺失导致了该基因移码突变,从而影响了其正常转录和表达水平。
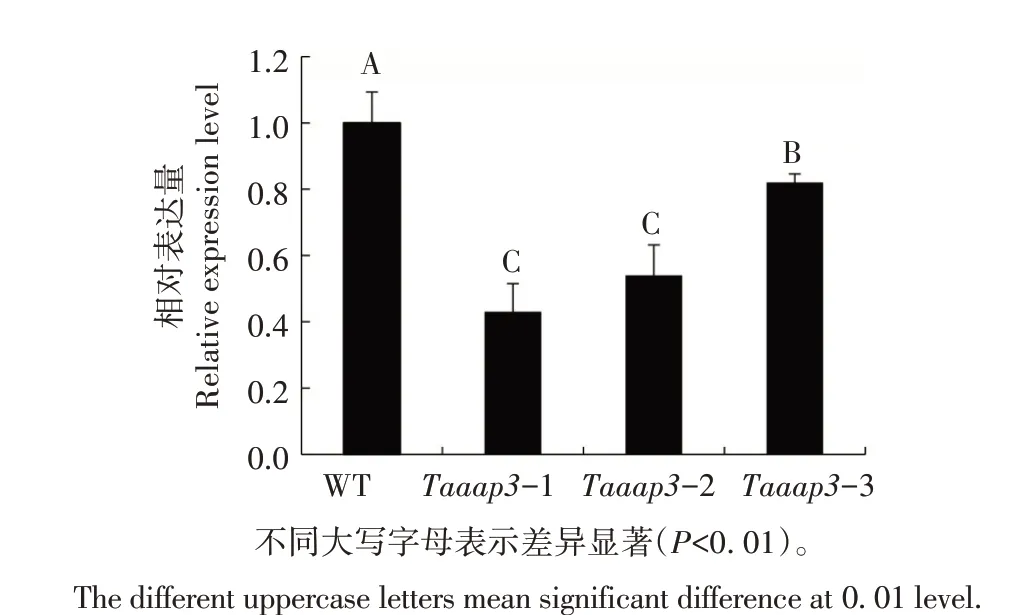
图8 Taaap3突变体叶片中TaAAP3基因的表达分析Fig.8 Expression analysis of TaAAP3 gene in leaves of Taaap3 mutants
2.5 Taaap3突变体的籽粒产量分析
在大田正常和低肥条件下,对突变体Taaap3的籽粒产量进行分析发现(图9),正常条件下,突变体Taaap3-1、Taaap3-2 和Taaap3-3 籽粒产量均高于其野生型,分别较野生型增产2.29%、3.40% 和0.93%,但均未达到显著水平;低肥条件下,3个突变体籽粒产量均显著或极显著高于其野生型,Taaap3-1、Taaap3-2 和Taaap3-3 分别较野生型增产6.89%、5.90% 和4.59%。综上,低肥条件下TaAAP3基因表达水平降低可提高小麦籽粒产量。
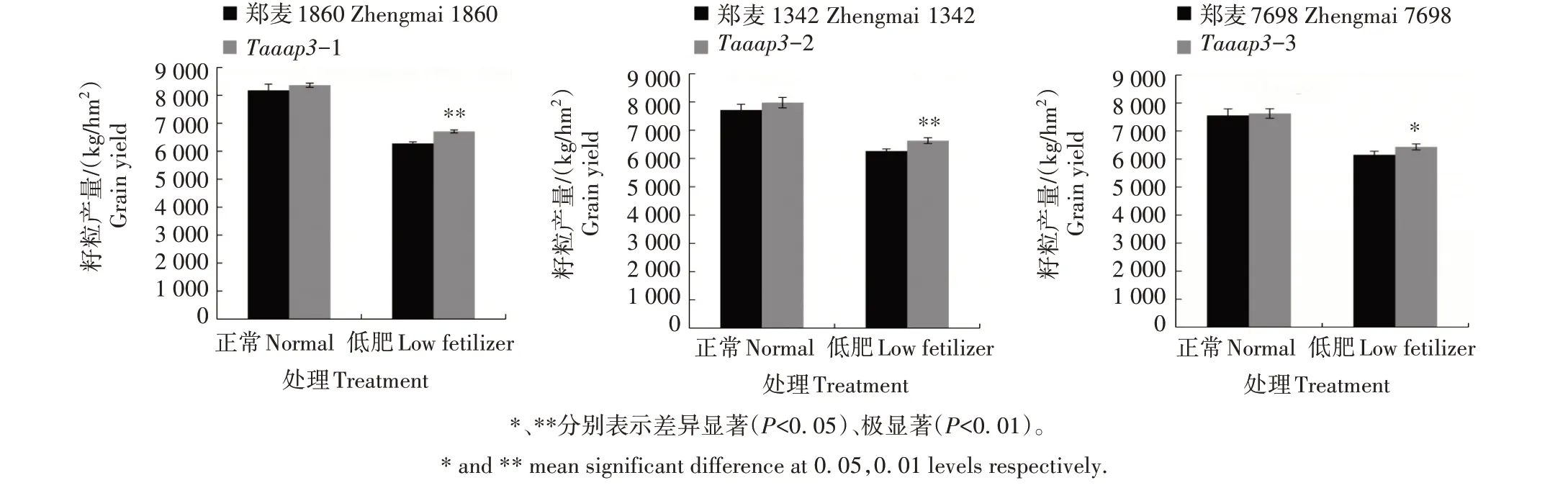
图9 Taaap3突变体田间籽粒产量分析Fig.9 Analysis of grain yields of Taaap3 mutants in field
3 结论与讨论
氨基酸转运蛋白在氨基酸跨膜转运中起着重要的作用,并直接或间接参与对植物生长发育至关重要的氮代谢过程。目前,有关植物氨基酸转运蛋白功能的研究主要集中在ATF(Amino acid transporter family)和APC(Amino acid polyamine and choline transporters)2 个家族。AAPs 是ATF 家族中的亚家族,已在植物中进行了广泛的功能研究。水稻中有19个AAPs基因,其中OsAAP6基因影响水稻中各种氨基酸的分布,并正向调控稻米蛋白质含量和品质[3]。有些研究发现,OsAAP3或OsAAP5基因被抑制表达后,通过调控赖氨酸和精氨酸浓度来提高籽粒产量[1,19]。OsAAP1基因通过调控中性氨基酸的吸收和再分配来影响植株生长和产量[20]。还有些研究发现,AtAAP3基因的T-DNA 插入突变体和AtAAP1基因的RNAi突变体的表型与野生型均没有明显差异,推测其原因是AtAAP2、AtAAP5和AtAAP6基因可能在某种程度上与AtAAP3基因功能互补[5,21-22],且AtAAP5基因可能还与AtAAP1基因功能互补。本研究结果表明,在小麦中敲除TaAAP3基因后,正常条件下,突变体Taaap3较野生型增产幅度较小,未达到显著水平,这与前人研究结果相似[5,21-22];低肥条件下,突变体Taaap3较野生型显著增产,且TaAAP3基因表达水平越低,增产幅度越大,即A、B、D 3 个亚基因组均发生突变比单个亚基因组发生突变增产明显。推测抑制TaAAP3基因表达可以提高小麦养分利用效率。今后将进一步深入研究TaAAP3基因在小麦中的功能。
目前,CRISPR/Cas9 技术已被广泛应用于水稻[23-25]、玉米[26-27]和小麦[28-29]等多种农作物性状改良研究。由于普通小麦是由A、B、D 3 个亚基因组组成的六倍体作物,具有庞大的基因组和功能冗余的基因。因此,同时突变位于多个亚基因组位点的多个等位基因是比较困难的。但随着分子生物学技术的迅猛发展,利用CRISPR/Cas9 技术已创制了抗病、优质专用和高产等小麦新种质。研究人员利用CRISPR/Cas9 技 术 定 点 敲 除TaEDR1(Enhanced disease resistance 1)和TaMLO(Mildew resistance locus)基因后,均获得了抗白粉病小麦新种质[30-31]。TaNFXL1(Nuclear transcription factor X box-binding-like 1)基因和TaSBEIIa(Starch branching enzyme Ⅱa)基因被定点敲除后,分别获得了抗赤霉病和高抗性淀粉小麦新种质[28,32]。本研究以生产上大面积推广的小麦品种郑麦7698、郑麦1860 和郑麦1342 为背景材料,利用CRISPR/Cas9 技术敲除了TaAAP3基因,分别获得了A 亚基因组纯合单突变类型和A、B、D 3个亚基因组纯合突变的三突变类型的突变体。TaAAP3基因在突变体中的表达量显著低于野生型,在三突变类型突变体Taaap3-1 和Taaap3-2 中的表达量均显著低于单突变类型突变体Taaap3-3。另外,低肥条件下,突变体Taaap3-1 和Taaap3-2 较野生型的增产幅度大于突变体Taaap3-3,突变体Taaap3-1增产幅度最大。说明小麦中TaAAP3基因突变类型不同,突变体产量表现亦有差异。在小麦中精准高效编辑该基因,可创制出高产高效突破性小麦新种质。CRISPR/Cas9技术虽可定点敲除目标基因,但无法高效地诱导点突变等编辑类型。引导编辑技术相比CRISPR/Cas9技术具有更高的特异性和安全性,将在未来农作物遗传改良中发挥重要作用。
猜你喜欢
麦类作物学报(2022年1期)2022-03-01
现代畜牧科技(2021年4期)2021-12-05
现代畜牧科技(2021年10期)2021-11-19
粮食加工(2020年4期)2020-02-28
农民致富之友(2019年33期)2019-12-20
河北农业科学(2018年2期)2018-07-26
商洛学院学报(2017年2期)2017-05-17
安徽医科大学学报(2016年12期)2017-01-15
山东农业工程学院学报(2016年6期)2016-12-01
