
盐胁迫下不同辣椒品种的耐盐性差异及生理生化指标响应
2023-12-11 15:09:16张建鹏
江苏农业科学 2023年16期

摘要:选取15个辣椒品种为试验材料,用150 mmol/L NaCl处理0、5、10、15 d后,通过表型分析筛选耐盐、盐敏感的辣椒品种,并比较分析6个不同盐敏感类型的辣椒品种生理指标的变化。结果表明,15个辣椒品种对盐胁迫表现出不同耐性,高度耐盐的品种为V4(新三樱八号)、V8(南韩天红一号F1)和V17(卡其三樱椒),盐敏感的品种为V3(朝天椒超级808)、V7(柘椒新一代)和V12(抗重茬先锋八号F1),其余品种均为中度耐盐。在150 mmol/L NaCl盐胁迫下,耐盐辣椒品种和盐敏感辣椒品种的叶绿素含量、净光合速率、胞间CO2浓度、气孔导度、蒸腾速率均呈下降趋势,但耐盐辣椒品种相应指标的下降幅度小于盐敏感辣椒品种,在盐胁迫后10、15 d,耐盐辣椒品种的叶绿素含量、净光合速率、胞间CO2浓度、气孔导度、蒸腾速率显著高于盐敏感辣椒品种;耐盐辣椒品种的游离脯氨酸含量呈现上升趋势,盐敏感辣椒品种的脯氨酸含量呈现先上升后下降的趋势,在盐胁迫后10、15 d,耐盐辣椒品种的游离脯氨酸含量显著高于盐敏感辣椒品种。在盐胁迫下,耐鹽辣椒品种和盐敏感辣椒品种的可溶性糖、可溶性蛋白含量均呈现上升趋势,耐盐辣椒品种相应指标的上升幅度大于盐敏感辣椒品种,可溶性糖、可溶性蛋白含量显著高于盐敏感辣椒品种;在盐胁迫下,耐盐辣椒品种和盐敏感辣椒品种的丙二醛含量均呈现上升趋势,耐盐辣椒品种的丙二醛含量较盐敏感辣椒品种上升得较慢,显著低于盐敏感辣椒品种;耐盐辣椒品种和盐敏感辣椒品种的过氧化氢酶(CAT)、过氧化物酶(POD)活性均呈现上升趋势,耐盐辣椒品种的CAT、POD活性上升幅度大于盐敏感辣椒品种,显著高于盐敏感辣椒品种。因此,在盐胁迫下,耐盐辣椒品种的细胞结构受到的损伤较小,叶绿素含量保持在较高水平,各项光合参数下降缓慢;耐盐辣椒品种通过产生脯氨酸、可溶性糖、可溶性蛋白来调节细胞内外的渗透平衡;耐盐辣椒品种的CAT、POD活性保持在较高水平,可以清除盐胁迫产生的活性氧,使膜脂过氧化程度降低,丙二醛含量维持在较低水平。
关键词:盐胁迫;辣椒;光合参数、抗氧化酶活性
中图分类号:S641.301文献标志码:A
文章编号:1002-1302(2023)16-0127-07
收稿日期:2023-01-11
基金项目:河南省科技攻关项目(编号:202102110199);河南省高等学校重点项目(编号:22B210009)。
作者简介:张建鹏(1980—),男,河南开封人,硕士,讲师,主要从事园艺作物生理与栽培方面的研究。E-mail:z1122330414@163.com。
全世界盐渍化的土地面积大约为9.5亿hm2,其中我国盐渍化的土地面积约为760万hm2[1]。我国是世界上盐渍化危害最为严重的国家之一,大量、严重发生盐渍化土壤的地区几乎寸草不生。同时,由于我国近年来土地大量施用化肥及工业污染加剧等原因,造成我国盐渍化土壤的面积不断扩大。因此,在我国开发和利用盐渍化土地对于农业的可持续及优质发展具有重要意义。
辣椒(Capsicum annuum)是一年生或有限多年生的草本植物,最初从美洲热带地区的墨西哥、秘鲁等地传到中国,目前在世界各国均普遍栽培。辣椒的维生素C含量在蔬菜中位于首位,辣椒的维生素B、胡萝卜素及铁、钙等矿物质含量同样非常丰富。食用辣椒可以大大刺激食欲,改善消化,温胃驱寒。此外,辣椒在临床上还可以用于治疗腰腿痛、冻疮、冻伤、外伤瘀肿、一般外科炎症等。因此,辣椒是我国最具有发展前景的经济作物之一[2]。
目前,关于经济作物的耐盐性研究逐渐增多[3-5],筛选耐盐辣椒品种,对辣椒适应盐胁迫的生理机制进行研究意义重大[6-7]。周静等研究了2个辣椒品种对盐胁迫响应的生理生化机制发现,当NaCl浓度增加时,辣椒幼苗叶片中的丙二醛、脯氨酸含量总体呈现上升趋势,抗氧化酶[超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)]的活性则呈现先上升后下降的趋势[6]。郑佳秋等研究了6个不同辣椒品种在盐胁迫下萌芽期、幼苗期的生理特性,结果表明,在盐胁迫下,不同辣椒品种萌芽期的种子发芽率、发芽势等均表现出不同程度的降低;幼苗期的抗氧化酶[SOD、POD、抗坏血酸过氧化物酶(APX)]活性总体表现为增强[8]。吉雪花等分别用不同浓度的NaCl、NaHCO3、K2SO4对辣椒进行胁迫处理,结果表明,低浓度盐均能够促进叶绿素的生成,相同浓度下K2SO4比NaCl、NaHCO3对叶绿素含量的抑制作用更强,随着盐胁迫时间的延长和盐浓度的增加,辣椒叶片细胞膜中的丙二醛的含量呈现逐渐上升的趋势[9]。
本试验拟对15个辣椒品种进行耐盐性筛选,测定盐胁迫下耐盐、盐敏感辣椒品种幼苗的光合参数、叶绿素含量、脯氨酸含量、可溶性糖含量、可溶性蛋白含量、丙二醛含量和抗氧化酶(CAT、POD)活性等生理指标,研究耐盐辣椒品种、盐敏感辣椒品种不同的耐盐性生理机制,以期解析辣椒响应盐胁迫的生理机制,并为耐盐辣椒品种的筛选及耐盐育种提供一定的理论依据。
1 材料与方法
1.1 试验材料
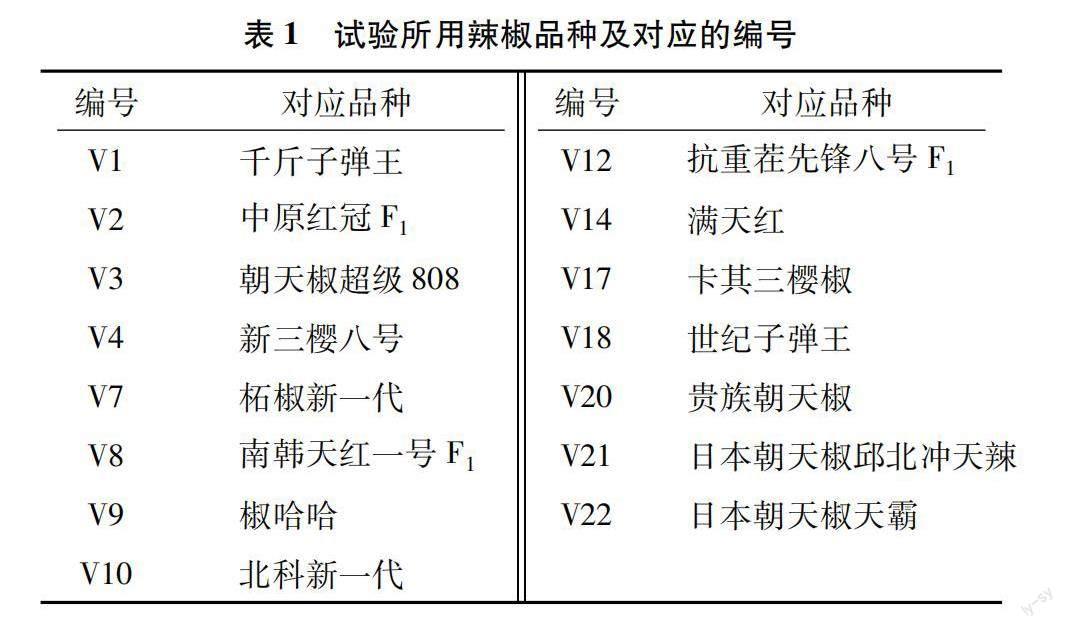
2021年3月于濮阳职业技术学院园艺专业实训基地开展试验。本试验选用的15个辣椒品种编号分别为V1、V2、V3、V4、V7、V8、V9、V10、V12、V14、V17、V18、V20、V21、V22,每个编号对应的辣椒品种见表1。
1.2 试验处理
辣椒幼苗期耐盐胁迫试验:将供试辣椒种子播种至穴盘中,在塑料大棚(昼温20~25 ℃、夜温 10~15 ℃)中进行育苗。当辣椒长至4叶1心期时,取长势一致的辣椒幼苗分别移栽至花盆中进行处理。盐胁迫处理灌施150 mmol/LNaCl溶液,每 3 d 浇灌1次盐溶液,共浇灌5次,每次每株辣椒浇灌100 mL盐溶液,每个处理设40株苗,以浇灌蒸馏水为对照组。分别在处理后0、5、10、15 d调查耐盐表型并进行相关生理指标的测定,测定的生理指标包含光合参数(净光合速率、气孔导度、胞间CO2浓度和蒸腾速率)、叶绿素含量、游离脯氨酸含量、可溶性糖含量、可溶性蛋白含量、丙二醛含量、抗氧化酶(CAT、POD)活性。
1.3 测定方法
测定光合参数时,用Ciras-1型光合测定系统在400 μmol/(m2·s)光照度下分別测定辣椒叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(E)等气体交换参数。测定叶绿素含量采用丙酮提取比色法[10],测定游离脯氨酸含量采用茚三酮比色法[10],测定可溶性蛋白含量采用考马斯亮蓝 G-250 染色法[10],测定可溶性糖含量采用蒽酮法[11],测定过氧化物酶(POD)活性采用愈创木酚法[11],测定过氧化氢酶(CAT)活性采用紫外分光光度吸收法[11],测定丙二醛含量采用硫代巴比妥酸法[12]。
1.4 数据统计
用Excel 2003对数据进行处理与作图,用SASS 9.1软件进行方差分析,并对平均值进行Duncans多重比较。
2 结果与分析
2.1 盐胁迫下辣椒幼苗的形态特征
盐胁迫对辣椒形态有明显的影响,在盐胁迫 15 d 后,品种V3、V7和V12叶片的萎蔫程度高,叶片发黄并下垂,并且在处理15 d后叶片脱落严重,为盐敏感辣椒品种(图1);品种V4、V8和V17叶片的萎蔫程度低,叶片挺立,为耐盐辣椒品种(图2);其余品种表现为中度萎蔫。下面只对上述6个品种进行分析。
2.2 盐胁迫对辣椒幼苗生理生化指标的影响
2.2.1 盐胁迫对辣椒幼苗叶片光合参数的影响 由图3可以看出,在盐胁迫下,耐盐辣椒品种V4、V8、V17和盐敏感辣椒品种V3、V7、V12的叶片净光合速率均呈现下降趋势,耐盐辣椒品种的净光合速率较盐敏感辣椒品种下降得缓慢。在盐胁迫后10、15 d,耐盐辣椒品种的净光合速率显著高于盐敏感辣椒品种。在盐胁迫后15 d,耐盐辣椒品种V4、V8、V17的净光合速率与处理前相比分别下降了60.93%、51.11%、66.37%,盐敏感辣椒品种V3、V7、V12的净光合速率与处理前相比分别下降了74.11%、80.73%、78.12%。
由图4可以看出,在盐胁迫下,辣椒叶片气孔导度总体呈现下降趋势,耐盐辣椒品种V4、V8、V17的下降幅度小于盐敏感辣椒品种V3、V7、V12。在盐胁迫后10、15 d,耐盐辣椒品种的气孔导度显著高于盐敏感辣椒品种。在盐胁迫后15 d,耐盐辣椒品种V4、V8、V17的叶片气孔导度相较于处理前分别下降了47.28%、54.06%、56.95%,盐敏感辣椒品种V3、V7、V12的叶片气孔导度相较于处理前分别下降了82.82%、93.40%、87.70%。
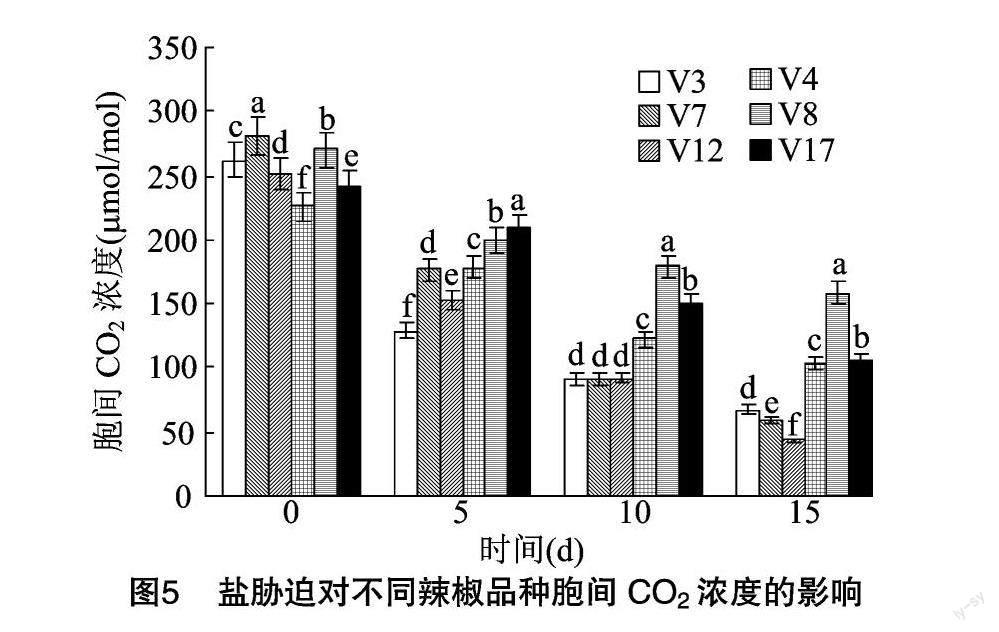
由图5可以看出,在盐胁迫下,辣椒叶片的胞间CO2浓度总体呈现下降趋势。盐胁迫后5、10、15 d,耐盐辣椒品种的胞间CO2浓度显著高于盐敏感辣椒品种。在盐胁迫后15 d,耐盐辣椒品种V4、V8、V17的胞间CO2浓度相较于处理前分别下降了54.43%、41.16%、56.43%;在盐胁迫后15 d,盐敏感辣椒品种V3、V7、V12的胞间CO2浓度相较于处理前分别下降了74.26%、78.92%、82.76%,耐盐辣椒品种的胞间CO2浓度较盐敏感辣椒品种下降得缓慢。
由图6可以看出,在盐胁迫下,辣椒叶片的蒸腾速率呈现下降趋势,其中耐盐辣椒品种的下降幅度小于盐敏感辣椒品种,在盐胁迫后5、10、15 d,耐盐辣椒品种的叶片蒸腾速率显著高于盐敏感辣椒品种。在盐胁迫后15 d,耐盐辣椒品种V4、V8、V17的叶片蒸腾速率相较于处理前分别下降了55.53%、61.56%、70.66%,盐敏感辣椒品种V3、V7、V12的叶片蒸腾速率相较于处理前分别下降了85.28%、84.39%、84.04%,耐盐辣椒品种的蒸腾速率较盐敏感辣椒品种下降得缓慢。
2.2.2 盐胁迫对辣椒幼苗叶绿素含量的影响 由图7可以看出,在盐胁迫下,辣椒中的叶绿素含量总体呈缓慢下降的趋势,耐盐辣椒品种V4、V8、V17的叶绿素含量显著高于盐敏感辣椒品种V3、V7、V12。在盐胁迫后15 d,耐盐辣椒品种V4、V8、V17的叶绿素含量相较于处理前分别下降了6.96%、7.38%、14.46%,盐敏感辣椒品种V3、V7、V12的叶绿素含量相较于处理前分别下降了6.07%、21.73%、6.11%。
2.2.3 盐胁迫对辣椒幼苗游离脯氨酸含量、可溶性糖含量、可溶性蛋白含量的影响 由图8可以看出,在盐胁迫下,耐盐辣椒品种V4、V8、V17的脯氨酸含量呈现上升趋势,盐敏感辣椒品种V3、V7、V12的脯氨酸含量呈现先上升后下降的趋势。在盐胁迫处理后10、15 d,耐盐辣椒品种的脯氨酸含量显著高于盐敏感辣椒品种。在盐胁迫处理后5 d,耐盐辣椒品种V4、V8、V17的脯氨酸含量相比于处理前分别上升了84.00%、106.62%、57.50%,在盐胁迫处理后10 d分别上升了119.75%、168.96%、103.72%,在盐胁迫处理后15 d分别上升了160.64%、211.73%、160.06%。在盐胁迫后5 d,盐敏感辣椒品种V3、V7、V12的脯氨酸含量相比于处理前分别上升了233.89%、239.96%、193.64%,在盐胁迫后10 d分别上升了128.72%、119.07%、95.69%,在盐胁迫后15 d分别上升了38.75%、85.44%、62.52%。
由图9可以看出,在盐胁迫下,耐盐辣椒品种V4、V8、V17和盐敏感辣椒品种V3、V7、V12的可溶性糖含量随时间呈现上升趋势。耐盐辣椒品种的可溶性糖含量上升幅度大于盐敏感辣椒品种,显著高于盐敏感辣椒品种。在盐胁迫后5 d,耐盐辣椒品种V4、V8、V17的可溶性糖含量相比处理前分别上升了82.63%、19.61%、202.65%,在盐胁迫处理后10 d分别上升了190.72%、93.10%、361.40%,在盐胁迫处理后15 d分别上升了358.14%、195.65%、615.90%。在盐胁迫后5 d,盐敏感辣椒品种V3、V7、V12的可溶性糖含量分别上升了77.42%、188.04%、23.11%,在盐胁迫后10 d分别上升了109.11%、380.62%、65.68%,在盐胁迫后15 d分别上升了258.68%、652.79%、122.79%。
由图10可以看出,在盐胁迫下,辣椒中的可溶性蛋白含量总体呈上升趋势,耐盐辣椒品种的可溶性蛋白含量显著高于盐敏感辣椒品种。在盐胁迫后5 d,耐盐辣椒品种V4、V8、V17的可溶性蛋白含量相比处理前分别上升了1215.02%、286.53%、912.08%,在盐胁迫后10 d时分别上升了 1 338.77%、267.77%、1 271.79%,在盐胁迫后15 d时分别上升了1 791.72%、386.24%、1 515.16%。在盐胁迫后5 d,盐敏感辣椒品种V3、V7、V12的可溶性蛋白含量分别上升了355.95%、94.21%、190.08%,在盐胁迫后10 d分别上升了459.58%、199.73%、322.44%,在盐胁迫后15 d分别上升了733.71%、275.45%、380.66%。
2.2.4 盐胁迫对辣椒幼苗过氧化氢酶活性、过氧化物酶活性的影响 由图11可以看出,在盐胁迫下,辣椒幼苗叶片过氧化氢酶活性呈上升趋势,耐盐辣椒品种过氧化氢酶活性显著高于盐敏感辣椒品种。在盐胁迫后5 d,耐盐辣椒品种V4、V8、V17过氧化氢酶活性相较于处理前分别上升了257.23%、194.13%、145.93%,在处理后10 d分别上升了584.44%、545.63%、361.67%,在处理后 15 d 分别上升了902.10%、815.79%、632.63%。在鹽胁迫处理5 d后,盐敏感辣椒品种V3、V7、V12过氧化氢酶活性相较于处理前分别上升了131.52%、120.59%、109.95%,在盐胁迫处理10 d时分别上升了217.99%、266.53%、225.78%,在盐胁迫处理15 d时分别上升了574.25%、665.55%、558.52%。
由图12可以看出,盐胁迫下的辣椒幼苗叶片POD活性总体呈现上升趋势,耐盐辣椒品种的POD活性显著高于盐敏感辣椒品种。在盐胁迫5 d时,耐盐辣椒品种V4、V8、V17的过氧化物酶活性相较于处理前分别上升了47.57%、71.32%、77.20%,在盐胁迫10 d时分别上升了128.48%、136.81%、129.09%,在盐胁迫15 d时分别上升了176.70%、186.19%、147.98%。在盐胁迫5 d时,盐敏感辣椒品种V3、V7、V12的过氧化物酶活性相较于处理前分别上升了24.14%、46.49%、56.39%,在盐胁迫处理10 d时分别上升了41.90%、88.09%、89.23%,在盐胁迫处理15 d时分别上升了79.91%、123.44%、125.91%。
2.2.5 盐胁迫对辣椒幼苗丙二醛含量的影响 丙二醛是膜脂过氧化过程中最重要的代表性产物之一,它的产生能够加剧膜的损伤,因此丙二醛含量的变化是反映细胞膜脂过氧化作用的一个重要指标[13]。由图13可以看出,在盐胁迫下,辣椒幼苗叶片的丙二醛含量呈上升趋势,其中耐盐辣椒品种的丙二醛含量较盐敏感辣椒品种上升得慢,显著低于盐敏感辣椒品种。在盐胁迫处理5 d时,耐盐辣椒品种V4、V8、V17的丙二醛含量相比处理前分别上升了44.12%、49.39%、59.18%,在盐胁迫后10 d时分别上升了71.30%、80.46%、83.86%,在盐胁迫后15 d时分别上升了72.35%、94.67%、107.05%。在盐胁迫后5 d,盐敏感辣椒品种V3、V7、V12的丙二醛含量相比处理前分别上升了70.82%、51.95%、68.47%,在处理后10 d分别上升了124.65%、106.67%、111.06%,在盐胁迫后 15 d 分别上升了173.98%、180.14%、199.53%。
3 讨论
植物的非生物胁迫反应受到许多因素控制,不同植物的耐受性和耐受机制不同。本试验结果表明,在盐胁迫下,耐盐辣椒品种的形态特征与盐敏感辣椒品种有明显差异,其生理指标在耐盐辣椒品种和盐敏感辣椒品种之间也有显著差异。
3.1 辣椒幼苗叶片叶绿素含量和光合参数对盐胁迫的响应
叶绿素是一类重要的含脂色素家族,存在于类囊体膜上,是植物进行光合作用的必备条件,在植物光合作用中的光反应阶段中起着核心作用,在植物对光能的吸收、传递和转化等方面具有重要的生理功能和作用[14]。盐胁迫加速了植物中叶绿体的降解,使得类囊体膜的稳定性降低,导致叶绿体对光能的吸收显著降低,最终使光合速率下降[15-16]。刘强等研究发现,NaCl处理后曼陀罗幼苗叶内叶绿素含量降低,叶片气孔导度显著下降,CO2进入叶肉的阻力随之增加,进而使得光合速率下降[17]。卢艳等研究发现,盐胁迫通过降低叶绿素含量导致叶片光能利用率降低,同时盐胁迫产生的渗透胁迫、离子毒害作用导致叶绿体的超微结构发生改变,进而导致光合能力降低[18]。
在本试验中,在盐胁迫下,耐盐辣椒品种V4、V8、V17和盐敏感辣椒品种V3、V7、V12的叶绿素含量、净光合速率、胞间CO2浓度、气孔导度、蒸腾速率均呈现下降趋势,但耐盐辣椒品种的下降幅度小于盐敏感辣椒品种。在盐胁迫10、15 d时,耐盐辣椒品种的叶绿素含量、净光合速率、胞间CO2浓度、气孔导度、蒸腾速率显著高于盐敏感辣椒品种。上述研究结果表明,在盐胁迫下,耐盐辣椒品种通过比盐敏感辣椒品种含有较高的叶绿素含量来维持一定的光能捕获能力,并通过较高的胞间CO2浓度、气孔导度和蒸腾速率来维持相对较高的净光合速率。
3.2 辣椒幼苗叶片游离脯氨酸、可溶性糖和可溶性蛋白含量对盐胁迫的响应
植物可以通过渗透物质调节来维持渗透平衡和适应胁迫环境。游离脯氨酸是植物中一种重要的有机渗透调节物质,同时也是一种抗氧化物质,它具有稳定细胞蛋白质的结构、平衡液泡中的高浓度的盐分、防止酶的变性失活、避免细胞质的脱水等生理功能和作用[19-20]。可溶性糖在许多植物中是非常有效的渗透调节剂,通过调节细胞渗透模式以维持细胞的水分平衡。可溶性蛋白质也参与植物的渗透调节,它可以维持细胞的渗透势和功能性蛋白质的数量,以此来减轻植物在胁迫下的遭受的伤害程度[21-22]。
植物在盐胁迫时可以合成大量如糖类、氨基酸类(脯氨酸、苹果酸)、蛋白质类等渗透调节物质来维持细胞的渗透平衡[23]。李海燕等研究发现,蜡梅可以通过提高游离脯氨酸含量、可溶性糖含量、可溶性蛋白含量来主动响应低盐胁迫[24]。郭茜茜等研究发现,不同低、中浓度下的盐胁迫可以提高高辣度辣椒叶片中的可溶性糖、可溶性蛋白等渗透调节物质含量[25]。
在本试验中,盐胁迫下的耐盐辣椒品种V4、V8、V17叶片中的游离脯氨酸含量呈上升趋势,盐敏感辣椒品种V3、V7、V12中的脯氨酸含量呈现出先升后降的趋势。在盐胁迫10、15 d,耐鹽辣椒品种的游离脯氨酸含量显著高于盐敏感辣椒品种。在盐胁迫下,耐盐辣椒品种、盐敏感辣椒品种的可溶性糖、可溶性蛋白含量均呈上升趋势,其中耐盐辣椒品种的上升幅度大于盐敏感辣椒品种,可溶性糖、可溶性蛋白含量也显著高于盐敏感辣椒品种。因此可见,在盐胁迫下耐盐辣椒品种通过积累相关的渗透调剂物质来缓解盐胁迫对植物细胞的毒害,维持细胞内外渗透平衡。
3.3 辣椒幼苗叶片中丙二醛含量、过氧化氢酶活性、过氧化物酶活性对盐胁迫的响应
丙二醛含量是反映植物细胞膜系统状况的重要生理指标,在盐胁迫下,植物细胞受损后会产生膜脂过氧化,而丙二醛是膜脂过氧化的最终降解产物,因此可用来反映植物遭受逆境伤害的程度[26]。CAT、POD是抗氧化防御系统的重要组成物质,CAT是将过氧化氢催化分解成氧和水的酶,POD可以以过氧化氢作为电子受体来催化底物氧化的酶,二者在盐胁迫下能维持较高的活性,协同作用可有效地抵抗耐盐胁迫诱导的氧化伤害[8,27]。
雷新慧等研究发现,盐胁迫处理增加了荞麦种子、幼苗叶片中的丙二醛含量,提高抗氧化酶(SOD、POD、CAT)的活性[28]。董建梅等研究发现,随着盐浓度的逐渐增加,石榴苗叶片中的丙二醛含量、CAT活性、POD活性随之升高[29]。马丽清等研究发现,耐盐苹果品种CAT、POD的活性随着盐胁迫的加大而增高,而盐敏感品种的CAT、POD 活性随着盐胁迫的加大而降低[30],说明膜保护酶系活性强弱与耐盐性呈正相关。
本试验中,盐胁迫下的耐盐辣椒品种V4、V8、V17和盐敏感辣椒品种V3、V7、V12的丙二醛含量均呈现上升趋势,其中耐盐辣椒品种丙二醛含量较盐敏感辣椒品种上升得慢,显著低于盐敏感辣椒品种。在盐胁迫下,耐盐辣椒品种、盐敏感辣椒品种的CAT、POD活性均呈现上升趋势,耐盐辣椒品种CAT、POD活性的上升幅度大于盐敏感辣椒品种,盐胁迫下耐盐辣椒品种的CAT、POD活性显著高于盐敏感辣椒品种。因此可见,耐盐辣椒品种能够通过较高的CAT、POD活性,有效清除掉在盐胁迫处理下产生的大量活性氧,使活性氧维持在较低的水平,保护细胞的膜系统,使盐胁迫下耐盐辣椒品种的丙二醛含量显著低于盐敏感辣椒品种,这与张景云等研究植物耐盐生理生化机制得出的结果[31]一致。
参考文献:
[1]李建国,濮励杰,朱 明,等. 土壤盐渍化研究现状及未来研究热点[J]. 地理学报,2012,67(9):1233-1245.
[2]徐珊珊,张广臣,叶景学. 盐碱胁迫对辣椒幼苗生长影响的研究[J]. 北方园艺,2011(8):5-9.
[3]巩志勇,辛建华,商小雨,等. 盐碱胁迫下香椿幼苗光合及抗逆生理特性[J]. 西北植物学报,2021,41(7):1199-1209.
[4]杨惠栋. 枳和资阳香橙等4种柑橘砧木的耐盐耐硼性差异及其生理研究[D]. 重庆:西南大学,2017:1-13.
[5]豆昕桐,王英杰,王华忠,等. 耐盐和盐敏感型小麦品种对NaCl胁迫的生理响应及耐盐性差异[J]. 生态学报,2021,41(12):4976-4992.
[6]周 静,徐 强,张 婷. NaCl胁迫对不同品种辣椒幼苗生理生化特性的影响[J]. 西北农林科技大学学报,2015,43(2):120-125.
[7]周 静. NaCl胁迫对辣椒种子、幼苗生长发育和叶绿素荧光特性的影响[D]. 合肥:安徽农业学,2014:2-9.
[8]郑佳秋,郭 军,梅 燚,等. 辣椒种子萌发和幼苗生理特性对盐胁迫的响应[J]. 江苏农业科学,2016,44(11):182-186.
[9]吉雪花,钱 渝. 三种盐胁迫对线辣椒生理特性的影响[J]. 北方园艺,2017(20):1-6.
[10]李合生,孙 群,赵世杰,等. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:130-137.
[11]孔祥生,易现峰. 植物生理学实验技术[M]. 北京:中国农业出版社,2008:267-258.
[12]赵世杰,李德全. 现代植物生理学实验指南[M]. 北京:科学出版社,1999:305-306.
[13]余海英,李廷轩,周健民. 典型设施栽培土壤盐分变化规律及潜在的环境效应研究[J]. 土壤学报,2006,43(4):571-576.
[14]王邦锡,何军贤,黄久常. 水分胁迫导致小麦叶片光合作用下降的非气孔因素[J]. 植物生理学报,1992,18(1):77-84.
[15]李海波,陈温福,李全英. 盐胁迫下水稻叶片光合参数对光强的响应[J]. 应用生态学报,2006(9):1588-1592.
[16]Hoai N T T,Shim I S,Kobayashi K,et al. Accumulation of some nitrogen compound in response to salt stress and their relationships with salt tolerance in rice (Oryza sativa L.) seedlings[J]. Plant Growth Regulation,2003,41(2):159-164.
[17]刘 强,周晓梅,王占武. NaCl处理对曼陀罗幼苗生长、光合、离子积累及抗氧化系统的影响[J]. 东北林业大学学报,2021,49(1):33-37.
[18]卢 艳,王 飞,韩明玉,等. NaCl 胁迫对4种砧穗组合苹果的生长及光合特性的影响[J]. 西北农业学报,2011,20(8):106-110.
[19]裘丽珍,黄有军,黄坚钦,等. 不同耐盐性植物在盐胁迫下的生长与生理特性比较研究[J]. 浙江大学学报,2006,32(4):420-427.
[20]徐鲜钧,沈宝川,祁建民. 植物耐盐性及其生理生化指标的研究进展[J]. 亚热带农业研究,2007,3(4):275-280.
[21]岳健敏,任 琼,张金池. 植物盐耐机理研究进展[J]. 林业科技开发,2015,29(5):9-13.
[22]Améglio T,Cochard H,Lacointe A,et al. Adaptation to cold temperature and response to freezing in walnut tree[J]. Acta Horticulturae,1999,544:247-254.
[23]孙聪聪,赵海燕,郑彩霞. NaCl胁迫对银杏幼树渗透调节物质及脯氨酸代谢的影响[J]. 植物生理学报,2017,53(3):470-476.
[24]李海燕,邵金彩,王 静,等. NaCl胁迫对5年生蜡梅生长及生理特性的影响[J]. 东北林业大学学报,2021,49(3):31-38.
[25]郭茜茜,吴 鹏,陈柏杰,等. 钠盐胁迫对不同辣度辣椒的耐受性影响研究初探[J]. 东北农业科学,2021,46(4):69-74,87.
[26]郭爱华. 外源褪黑素对盐胁迫下苦菜幼苗生长的影响[J]. 江苏农业科学,2022,50(13):153-157.
[27]Meloni D A,Oliva M A,Martinez C A,et al. Photosynthesis and activity of superoxide dismutase,peroxidase and glutathione reductase in cotton under salt stress[J]. Environ Exp Bot,2003,49(1):69-76.
[28]雷新慧,万晨茜,陶金才,等. 褪黑素与2,4-表油菜素内酯浸种对盐胁迫下荞麦发芽与幼苗生长的促进效应[J]. 作物学报,2022,48(5):1210-1221.
[29]董建梅,劉翠玉,赵玉洁,等. 盐胁迫对石榴生长与生理生化特性的影响[J]. 西南林业大学学报,2021,41(1):47-54.
[30]马丽清,韩振海,周二峰,等. 盐胁迫对珠美海棠和山定子膜保护酶系统的影响[J]. 果树学报,2006,23(4):495-499.
[31]张景云. 黄瓜不同抗盐品种生理生化特性的研究[D]. 哈尔滨:东北农业大学,2007:28-37.
猜你喜欢
少儿科学周刊·少年版(2022年18期)2022-05-30 21:40:32
少儿科学周刊·少年版(2022年18期)2022-05-30 10:48:04
少儿科学周刊·少年版(2022年18期)2022-05-30 10:48:04
小哥白尼(神奇星球)(2021年6期)2021-07-28 06:31:36
中外文摘(2020年9期)2020-06-01 13:47:56
江苏农业科学(2016年8期)2017-02-15 20:17:01
江苏农业科学(2016年8期)2017-02-15 19:41:25
江苏农业科学(2016年8期)2017-02-15 19:41:06
山东农业科学(2016年12期)2017-01-21 14:20:16
中国科技博览(2016年1期)2016-04-25 07:47:28
