
黑麦草和苕子混合翻压还田对土壤酶活性和养分的短期影响
2023-12-11 21:57:58潘语卓邱泽龙王伟虎康健龙万潮政石蕊陆艺引肖世豪鲁美娟杨文亭
江苏农业科学 2023年16期
潘语卓 邱泽龙 王伟虎 康健龙 万潮政 石蕊 陆艺引 肖世豪 鲁美娟 杨文亭
摘要:种植覆盖作物是优化种植结构、实现化肥减量增效的有效途径之一。深入了解覆盖作物翻压腐解对土壤养分、酶活性的影响,有助于优化作物种植的氮素管理。采用室内培养模拟试验,研究覆盖作物种类[空闲(CK)、黑麦草单播(R)、苕子单播(H)、黑麦草/苕子混播(RH)]和种植模式[种植甜玉米(C)和不种植甜玉米]对覆盖作物翻压腐解前期(0~21 d)土壤养分、土壤酶活性动态变化的影响。结果表明:(1)与对照相比,覆盖作物翻压还田处理显著提高了土壤β-葡萄糖苷酶活性、碱解氮含量和速效钾含量。土壤β-葡萄糖苷酶活性、碱解氮含量在不同覆盖作物处理之间存在显著差异,苕子翻压处理的β-葡萄糖苷酶活性、碱解氮含量在覆盖作物翻压4~14 d时均优于混播、黑麦草处理,且混播处理的碱解氮含量显著高于黑麦草处理。(2)在种植甜玉米的条件下,苕子处理的碱解氮含量在覆盖作物翻压21 d时较覆盖作物翻压4、7 d时均有显著下降,黑麦草/苕子混播处理的碱解氮含量在腐解前期无显著差异。(3)与不种甜玉米相比,种植甜玉米的处理显著降低了苕子和混播处理在覆盖作物翻压7 d时的土壤β-葡萄糖苷酶活性和β-葡萄糖苷酶-脲酶活性比,但显著提高了混播处理在覆盖作物翻压21 d时的土壤 β-葡萄糖苷酶活性和β-葡萄糖苷酶-脲酶活性比。(4)苕子翻压后,土壤脲酶活性与土壤有机质含量、pH值及碱解氮含量存在显著相关性,混播翻压后土壤酶活性与有效磷含量、速效钾含量的相关性显著增加。综上所述,覆盖作物翻压能够在短期内提高土壤速效养分含量和β-葡萄糖苷酶活性,苕子翻压后的土壤碱解氮含量显著高于黑麦草,黑麦草和苕子混合翻压腐解能够保持合理的碱解氮浓度,并延长其有效性时间。
关键词:覆盖作物;土壤β-葡萄糖苷酶;土壤脲酶;土壤养分
中图分类号:S154.2;S158文献标志码:A
文章编号:1002-1302(2023)16-0230-10
收稿日期:2022-11-23
基金项目:江西省研究生创新专项(编号:YC2021-S367);大学生创新创业训练计划(编号:202210410223);国家自然科学基金(编号:31901125)。
作者简介:潘语卓(1997—),女,四川乐山人,硕士研究生,主要研究方向为覆盖作物培肥农田地力机理。E-mail:962863261@qq.com。
通信作者:杨文亭,博士,副研究员,主要研究方向为覆盖作物提高作物产量和培肥地力的机理机制研究。E-mail:wtyang@jxau.edu.cn。
近年來,覆盖作物在农业生态系统中得到了广泛的实践运用[1-3]。大部分覆盖作物是中国传统农业中的绿肥,是一种高效清洁的绿色资源[4]。国内外的研究结果表明,覆盖作物在增加下茬作物产量[5]、提高作物氮肥利用效率、减少化学肥料用量[6]、改善土壤肥力[7]、固碳减排[8]等方面都有正效应,从而有利于维持土壤健康[9]。因此,种植覆盖作物被认为是一种可以实现农业可持续发展的新策略[10]。
覆盖作物翻压腐解释放养分和改善土壤酶活性是促进下茬作物生长和增产的主要原因。对于作物的吸收利用而言,覆盖作物腐解释放的氮与化肥氮肥是等效的,覆盖作物腐解释放氮的过程能够延长生物有效性氮在土壤中的滞留时间[11]。土壤中的胞外酶活性很早就被用于研究覆盖作物腐解过程中的养分循环和生化过程。由于胞外酶在产生过程中会从有机物质中获取营养物质,因此土壤胞外酶活性能够表征微生物的代谢需求[12]。同时,农业生态系统中关于碳氮循环相关酶活性与无机氮之间相互关系的研究已开展[13],但主要集中在某个时期,没有进行连续监测。土壤β-葡萄糖苷酶能够分解纤维素生成葡萄糖,是纤维素分解酶系中的重要组成成分之一,在土壤微生物的糖类代谢方面具有重要生理功能[13]。土壤脲酶主要作用于酰胺的C—N键,能够水解尿素生成氨[14]。由此可见,β-葡萄糖苷酶、脲酶可以分别作为碳多聚体降解以及氨释放的标记[15]。已有文献报道,β-葡萄糖苷酶、脲酶活性与土壤养分具有很高的相关性[16],但这种高相关性在红壤地不同覆盖作物种类翻压还田、甜玉米种植过程中是否存在还不清楚。因此,本研究假设不同覆盖作物种类翻压还田会影响甜玉米地土壤养分和酶活性,通过室内模拟试验,研究覆盖作物种类(黑麦草单播、苕子单播、黑麦草/苕子混播)和种植模式(种植甜玉米和不种植甜玉米)对覆盖作物翻压腐解前期土壤养分含量、酶活性变化的影响,以期验证覆盖作物翻压还田是否能改善土壤氮素有效性、土壤酶活性,从而为进一步优化设计覆盖作物混播组合和种植模式、维持土壤健康提供一定技术支持。
1 材料与方法
1.1 试验材料
供试甜玉米品种为赣科甜6号,覆盖作物为黑麦草(Lolium multiflorum Lam.)、紫花光叶苕子(Vicia villosa Roth var.)。黑麦草、紫花光叶苕子分别于2021年3月24日、3月29日收获,于60 ℃烘干至恒质量后,剪成1 cm左右的小段,干燥后保存备用。黑麦草、苕子的氮含量分别为27.15、40.31 g/kg,碳含量分别为365.95、310.89 g/kg。
供试土壤为红壤,采自2017年开始甜玉米间作大豆种植模式的定位试验田,土壤β-葡萄糖苷酶 活性为79.67 μmol/(g·d),脲酶活性为 3 593.21 μg/(g·d),蔗糖酶活性为 18.09 mg/(g·d),基本理化性质如下:有机质含量27.14 g/kg,pH值3.99(土与1 mol/L KCl的质量比为1∶5),碱解氮含量66.5 mg/kg,速效磷含量19.89 mg/kg,速效钾含量68 mg/kg。
1.2 试验设计
本试验于2021年9月6—27日在江西农业大学作物生理生态与遗传育种教育部重点实验室进行。试验设置覆盖作物种类、种植模式2个因素。覆盖作物种类包括空闲(CK)、单播黑麦草翻压(R)、单播苕子翻压(H)、黑麦草/苕子混播(质量比1∶1)翻压(RH),种植模式包括种植甜玉米(C)和不种甜玉米,共8个处理(表1)。
试验用高6.5 cm、上端口直径7.0 cm、下端口直径4.0 cm的塑料杯作为培养盆,每个盆内均放置黑色塑料袋,防止根系见光。每个容器内添加130 g干土,黑麦草、苕子和两者混合物各称取1.3 g直接添加至培养盆中。混合物内黑麦草、苕子的质量比为1∶1,用蒸馏水补足土壤含水量至20%(大田平均含水量),培养期间通过称质量法进行补水,使土壤含水量维持在20%。需要种植甜玉米的处理每盆直播甜玉米种子2粒,放置于人工气候箱中培养,温度设置为25 ℃,光—暗周期为16 h—8 h,光照度为13 000 lx,待甜玉米发芽后每盆保留1株。土壤取样时间为4、7、14、21 d,共取样4次。每个处理每次随机取4盆,为保证试验顺利进行,每个处理均备份4盆,共176盆。
1.3 测定项目与方法
土壤养分的测定参考鲍士旦主编的《土壤农化分析方法》[17],土壤pH值用雷磁酸度计测定(土∶1mol/L KCl为1∶5)。去除覆盖作物后采用浓硫酸-重铬酸钾外加热法测定土壤有机质含量,土壤碱解氮含量采用碱解扩散法测定,速效磷含量采用钼锑抗比色法(普析通用T6新世纪紫外可见分光光度计)测定,速效钾含量采用火焰分光光度法(上海精科火焰光度计——FP6410)测定。土壤酶活性的测定参考关松荫主编的《土壤酶及其研究法》[18],土壤β-葡萄糖苷酶活性采用硝基酚比色法测定,土壤蔗糖酶活性采用3,5-二硝基水杨酸比色法测定,土壤脲酶活性采用奈氏试剂——靛酚蓝比色法测定。
1.4 数据处理
用Excel 2022进行数据处理,用SPSS 26.0进行数据分析,用Duncans法进行显著性检验。土壤酶活性与养分间的相关性用Pearson相关性分析法,用Origin 2022软件进行作图。
2 结果与分析
2.1 覆盖作物翻压后的土壤酶活性
2.1.1 土壤β-葡萄糖苷酶活性 从图1可以看出,无论是否种植甜玉米,在覆盖作物翻压后4~14 d,土壤β-葡萄糖苷酶活性均显著高于对照。在覆盖作物翻压后21 d,在种植甜玉米处理下,不同覆盖作物翻压后土壤的β-葡萄糖苷酶活性显著高于对照;在未种植甜玉米处理下,黑麦草、苕子翻压处理的β-葡萄糖苷酶活性均显著高于对照,混播翻压处理的β-葡萄糖苷酶活性与对照间无显著差异。与未种甜玉米的处理相比,苕子翻压后种植甜玉米的土壤β-葡萄糖苷酶活性在覆盖作物翻压后7、14 d分别显著下降了66.06%、30.94%,混播翻压后仅在7 d时显著下降了33.57%;但在覆盖作物翻压后21 d时,混播翻压后种植甜玉米的土壤 β-葡萄糖苷酶活性显著增加了102.90%。
与覆盖作物翻压后4 d时相比,在未种植甜玉米处理下,苕子、混播翻压后7 d的土壤β-葡萄糖苷酶活性均显著增加,分别增加了53.82%、42.05%;在种植甜玉米处理下,苕子翻压后14 d的土壤β-葡萄糖苷酶活性显著增加了46.79%,混播翻压后均无显著差异。与覆盖作物翻压后14 d相比,在未种植甜玉米处理下,苕子、混播翻压后21 d的土壤β-葡萄糖苷酶活性分别显著降低了59.39%、46.02%;在种植甜玉米处理下,黑麦草、苕子翻压后的土壤β-葡萄糖苷酶活性在覆盖作物翻压后21 d时均显著降低,分别降低了35.10%、27.77%,混播翻压后无显著差异,表现出较好的酶活稳定性。
2.1.2 土壤脲酶活性 从图2可以看出,覆盖作物翻压后7 d时,无论是否种植甜玉米,黑麦草翻压的土壤脲酶活性均显著高于苕子翻压处理,分别增加了14.65%、12.18%。在种植甜玉米处理下,与对照相比,混播翻压后14 d的土壤脲酶活性显著增加了16.73%,其他时期均无显著差异;与苕子翻压处理相比,混播翻压后7 d的土壤脲酶活性有显著增加,但在覆盖作物翻压后21 d时有显著下降。
与覆盖作物翻压后4 d相比,苕子翻压处理后,无论是否种植甜玉米,土壤脲酶活性均在覆盖作物翻压后21 d时显著增加,分别增加了15.13%、37.52%;混播翻压后,种植甜玉米的土壤脲酶活性在覆盖作物翻压后14 d显著增加了31.21%;黑麦草翻压后,随着时间的变化,土壤脲酶活性无显著差异。
2.1.3 土壤蔗糖酶活性 從图3可以看出,与对照相比,在不同覆盖作物翻压后21 d,种植甜玉米的土壤蔗糖酶活性均显著增加,分别增加了69.87%、73.41%和53.07%;未种植甜玉米的土壤蔗糖酶活性仅在混播翻压后显著增加了26.29%;在其余时期,不同种覆盖作物翻压后对土壤蔗糖酶活性均无显著影响。覆盖作物翻压后的土壤蔗糖酶活性在4个取样时期无显著变化。
2.1.4 土壤β-葡萄糖苷酶:脲酶活性比率 从图4可以看出,苕子翻压7 d后,未种植甜玉米的土壤β-葡萄糖苷酶-脲酶活性比等于1.0,其他处理均小于1.0。与对照相比,无论是否种植甜玉米,翻压覆盖作物后4~14 d内,土壤β-葡萄糖苷酶-脲酶活性比均显著升高。在覆盖作物翻压后21 d,相比于对照,未种植甜玉米条件下,仅黑麦草翻压后的土壤β-葡萄糖苷酶-脲酶活性比显著升高了59.55%,混播处理还有下降的趋势;在种植甜玉米的条件下,覆盖作物翻压后的土壤β-葡萄糖苷酶-脲酶活性比均显著升高,黑麦草、苕子和混播翻压处理的土壤β-葡萄糖苷酶-脲酶活性比分别升高了124.33%、157.28%和186.40%。在未种植甜玉米的条件下,覆盖作物翻压后4~14 d,苕子翻压后的土壤β-葡萄糖苷酶-脲酶活性比均显著高于黑麦草,分别升高了36.21%、168.75%和114.20%;混播处理的土壤β-葡萄糖苷酶-脲酶活性比在覆盖作物翻压后7 d时也显著高于黑麦草处理,提高了94.02%。
与覆盖作物翻压后4 d相比,在未种植甜玉米处理下,苕子翻压后的土壤β-葡萄糖苷酶-脲酶活性比在覆盖作物翻压后7、14 d均显著升高,分别升高了59.09%、51.48%;混播翻压后的土壤β-葡萄糖苷酶-脲酶活性比在覆盖作物翻压7 d时显著升高了40.74%;黑麦草翻压后无显著变化。在种植甜玉米处理下,黑麦草翻压后21 d的土壤β-葡萄糖苷酶-脲酶活性比显著下降了19.66%;苕子翻压后的土壤β-葡萄糖苷酶-脲酶活性比在覆盖作物翻压后7 d显著下降了22.99%;混播翻压后无显著变化。
2.2 覆盖作物翻压后的土壤养分
2.2.1 土壤有机质含量 从图5可以看出,在未种植甜玉米条件下,相比于对照(CK),翻压覆盖作物(黑麦草、苕子、黑麦草/苕子混播)的土壤有机质含量在覆盖作物翻压后4、7、14 d均显著增加。相同时期翻压不同种类覆盖作物对土壤有机质含量均无显著影响。在种植甜玉米条件下,相比于对照(CKC),苕子翻压后的土壤有机质含量在覆盖作物翻压后4、7、21 d均有显著增加,分别增加了5.24%、6.94%、14.61%,黑麦草、混播翻压后的土壤有机质含量分别在覆盖作物翻压4、21 d时显著增加,分别增加了7.31%、17.39%。
与覆盖作物翻压4 d相比,黑麦草翻压后,未种植甜玉米的土壤有机质含量在覆盖作物翻压14、21 d 时均显著增加,分别增加了13.24%、22.98%。苕子翻压后种植甜玉米,土壤有机质含量在覆盖作物翻压14、21 d均显著增加,分别增加了15.72%、9.41%。混播翻压后,未种植、种植甜玉米的土壤有机质含量均在覆盖作物翻压后14、21 d显著增加,分别增加了11.93%、24.48%和25.53%、16.11%。
2.2.2 土壤pH值 从图6可以看出,无论是否种植甜玉米,不同种类覆盖作物翻压后4 d的土壤pH值均显著高于对照。在未种植甜玉米处理下,相比于对照,黑麦草翻压后14、21 d的土壤pH值显著下降,分别降低了0.23、0.25,混播翻压后21 d的土壤pH值也有显著降低,降低了0.26。在种植甜玉米处理下,相比于对照,黑麦草翻压后21 d的土壤pH值显著降低,但在苕子处理下有显著升高。
覆盖作物翻压后,土壤pH值整体呈波动上升的趋势。与覆盖作物翻压4 d相比,未种植甜玉米时,不同种类覆盖作物翻压后,其他时期的土壤pH值均无显著变化;但种植甜玉米时,黑麦草、苕子翻压后的土壤pH值在覆盖作物翻压14 d时显著增加,分别增加了0.22、0.18。
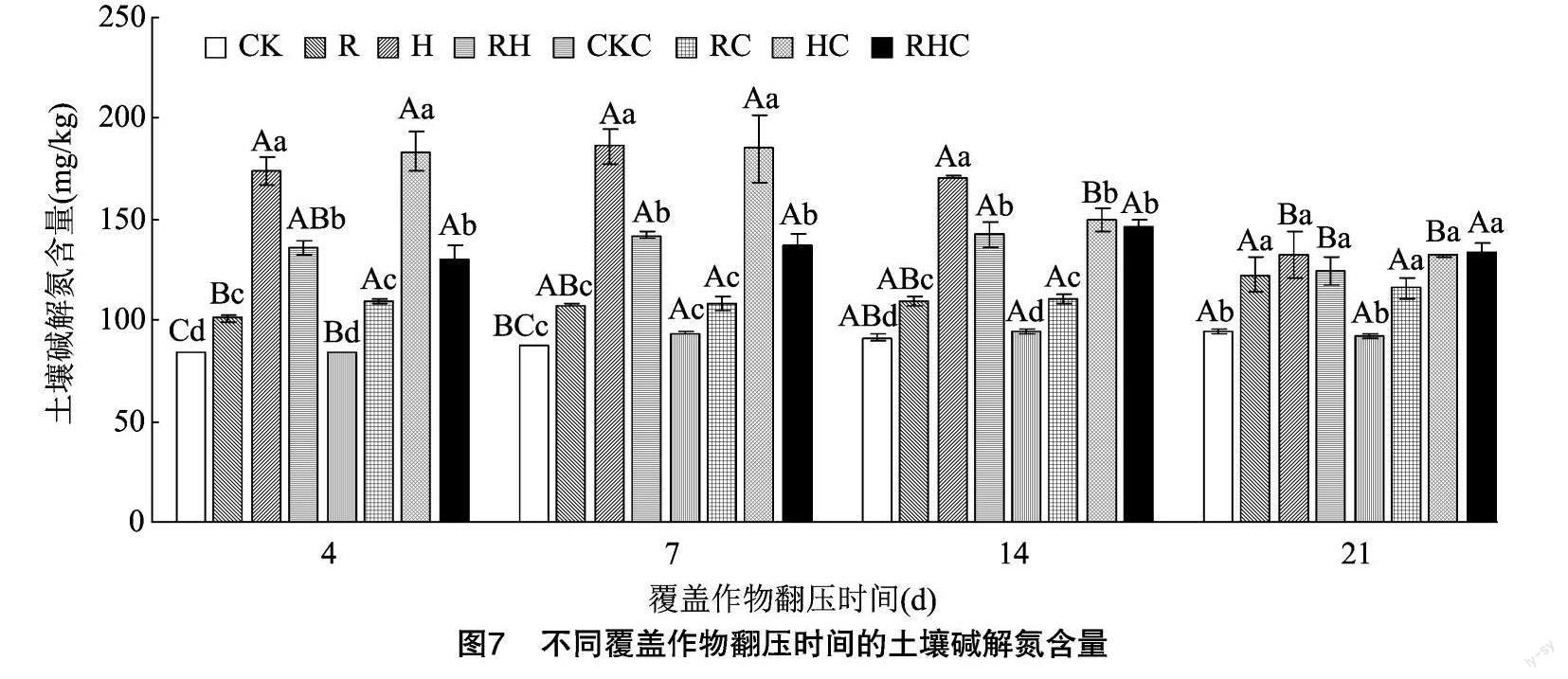
2.2.3 土壤碱解氮 从图7可以看出,无论是否种植甜玉米,苕子翻压后的土壤碱解氮含量均呈显著下降的趋势,而黑麦草和混播翻压处理后,土壤碱解氮含量均维持稳定,且无显著差异。相比于对照,除了种植甜玉米覆盖作物翻压7 d的黑麦草,其他覆盖作物翻压后的土壤碱解氮含量均显著升高。相比于黑麦草,苕子和混播翻压后的土壤碱解氮含量在覆盖作物翻压4~14 d均显著增加。苕子翻压后的土壤碱解氮含量在覆盖作物翻压4、7 d时均显著高于其他处理。
无论是否种植甜玉米,黑麦草翻压后21 d的土壤碱解氮含量较覆盖作物翻压后4 d时均显著增加,分别增加了21.16%、6.20%,但苕子翻压后 21 d 较覆盖作物翻压后4 d显著降低,分别降低了23.56%、28.10%。混播翻压后,无论是否种植甜玉米,土壤碱解氮含量均在覆盖作物翻压后14 d达到最大值,但在未种植甜玉米处理下,土壤碱解氮含量在覆盖作物翻压后21 d较覆盖作物翻压后14 d显著下降,降低了21.54%,种植甜玉米的土壤碱解氮含量在4个取样时期均无显著差异。
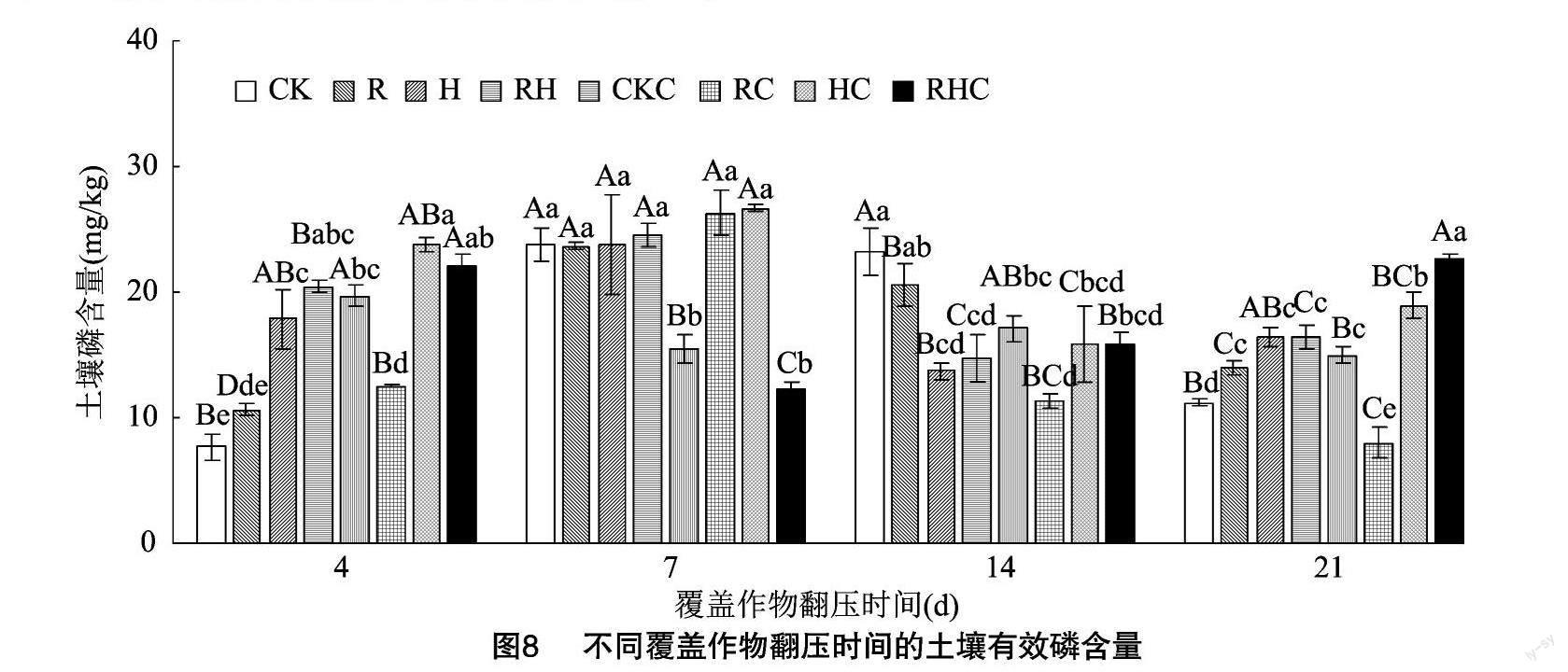
2.2.4 土壤有效磷、速效钾含量 从图8可以看出,在未种植甜玉米处理下,相比于对照,苕子和混播翻压后4 d的土壤有效磷含量显著增加,但覆盖作物翻压14 d时均有显著下降;覆盖作物翻压21 d时,不同覆盖作物翻压后的土壤有效磷含量均显著增加,分别增加了24.90%、46.97%、46.56%。在种植甜玉米处理下,相比于对照,黑麦草翻压后,土壤有效磷含量在覆盖作物翻压后7 d显著增加了70.14%,但在覆盖作物翻压4、14、21 d时显著下降。苕子翻压后,土壤有效磷含量分别在覆盖作物翻压后4、7、21 d显著增加,分别增加了20.40%、72.77%、26.61%。混播翻压后,土壤有效磷含量在覆盖作物翻压后21 d显著增加了51.43%。与覆盖作物翻压后4 d相比,黑麦草翻压后,未种植甜玉米处理的土壤有效磷含量在覆盖作物翻压后7~21 d均显著增加,种植甜玉米的土壤有效磷含量均在覆盖作物翻压后7 d显著增加。在混播翻压处理下,未种植甜玉米的土壤有效磷含量在覆盖作物翻压后7 d显著增加了20.43%,但在覆盖作物翻压后14、21 d顯著下降。
从图9可以看出,在覆盖作物翻压后7~21 d,不同种类覆盖作物翻压后的土壤速效钾含量之间均无显著差异。在未种植甜玉米处理下,相比于对照,翻压覆盖作物后土壤钾含量均显著增加,增加幅度分别为118.98%~131.89%、124.32%~133.90%、108.06%~128.65%。在种植甜玉米处理下,相比于对照,翻压覆盖作物后土壤钾含量均显著增加,增加幅度分别为102.40%~123.73%、103.45%~129.51%、91.63%~123.33%。
2.3 土壤酶活性与土壤养分的相关性分析
由图10可知,不同覆盖作物翻压后,土壤酶活性与土壤养分之间的相关性存在显著差异。黑麦草翻压后,土壤β-葡萄糖苷酶活性与土壤pH值呈极显著正相关,与蔗糖酶、脲酶活性间无显著相关性。苕子翻压后,土壤脲酶活性与土壤有机质含量和pH值均呈显著正相关,与土壤碱解氮呈极显著负相关;土壤蔗糖酶活性与土壤速效钾含量呈显著负相关;土壤β-葡萄糖苷酶活性与土壤养分均未有显著相关性。混播翻压后,土壤β-葡萄糖苷酶活性与土壤碱解氮、有效磷含量均呈显著正相关;土壤脲酶活性与土壤有机质含量呈极显著正相关,与土壤速效钾含量呈显著正相关,与土壤有效磷含量呈显著负相关;土壤蔗糖酶活性仅与速效钾含量呈显著正相关。
3 讨论
3.1 土壤酶活性
在本试验中,黑麦草、苕子及二者混合翻压均能显著提高腐解前期(覆盖作物翻压后21 d)的土壤β-葡萄糖苷酶活性,这与Nevins等的研究结果[19]相似。覆盖作物翻压还田后,一方面,其残体能够释放出多种酶类;另一方面,还田的作物向土壤输入了大量碳水化合物(包括纤维素、蔗糖)和氮素,促进了土壤微生物生长繁殖与代谢及甜玉米根系生长,进而增加了相关水解酶的浓度。其中 β-葡萄糖苷酶在纤维素降解中起到关键作用[20]。此外,覆盖作物处理与对照相比能够减少水分蒸发损失,保持较高的土壤含水量,有利于微生物的代谢活动[21]。
土壤酶活性受到不同覆蓋作物腐解释放碳、氮的影响[22]。在本试验中,相比于不种甜玉米处理,种植甜玉米的处理显著降低了苕子和混播处理7 d时土壤β-葡萄糖苷酶活性和β-葡萄糖苷酶-脲酶活性比,但显著增加了混播处理21 d时土壤 β-葡萄糖苷酶活性和β-葡萄糖苷酶-脲酶活性比。可能主要由于苕子氮含量较高(40.31 g/kg),碳氮比低于黑麦草,前期腐解释放的易降解碳氮较多,使其在前期腐解较快[15],初期(7 d)腐解释放的氮素较多,β-葡萄糖苷酶-脲酶活性比较好,土壤 β-葡萄糖苷酶活性较高;另一方面,由于温度适宜(25 ℃),覆盖作物翻压7 d时甜玉米已有1叶1心,甜玉米吸收利用了部分氮素,抑制了微生物生长[23],从而导致种植甜玉米处理中的土壤β-葡萄糖苷酶活性降低。覆盖作物翻压后21 d时,种植甜玉米的土壤β-葡萄糖苷酶活性显著提高,可能由于甜玉米根系分泌物增加为微生物生长提供了养分[24];另一方面,随着翻压腐解时间的推进,易降解有机碳减少,土壤β-葡萄糖苷酶酶活性均有较大幅度降低,但黑麦草的腐解释放较苕子慢[25],减缓了土壤酶活性的降低,从而有利于混播处理保持较高的土壤β-葡萄糖苷酶活性。短期内覆盖作物腐解速率较快,但在腐解中后期覆盖作物腐解速率变缓,关于土壤酶活性在覆盖作物腐解中后期会有何响应还需要进一步研究。
本研究结果表明,β-葡萄糖苷酶-脲酶的比值均小于1.0(H处理除外),说明脲酶的活性较高,土壤中微生物比较活跃,相对于氮素,碳是主要限制因素,需要补充更多碳源[26]。相比于对照,翻压黑麦草、苕子和黑麦草/苕子混播均显著增加了酶活性比,在一定程度上说明覆盖作物腐解增加了土壤碳源,促进了微生物的生长,提高了β-葡萄糖苷酶活性。相比于苕子,混播翻压后的酶活性比较低,可能由于苕子氮素含量高,腐解释放的氮素多,在一定程度上抑制了土壤脲酶活性[27]。另一方面,苕子翻压腐解过程中有较高的β-葡萄糖苷酶活性,但是随着试验时间的延长,混播翻压后种植甜玉米的β-葡萄糖苷酶-脲酶比维持稳定,未有显著变化,而苕子翻压后无论是否种植甜玉米,β-葡萄糖苷酶-脲酶比值均逐渐降低,且在覆盖作物翻压后21 d时显著低于混播翻压,表明混播处理更有利于甜玉米的生长。在之后的研究中为了更好地平衡土壤碳氮相关的酶活性,需要考虑引进更多额外的碳源或者更高碳含量的覆盖作物。
3.2 土壤养分
种植覆盖作物能活化富集土壤磷、钾等养分[28],豆科覆盖作物还能增加生物固氮量[22],而这种生物固氮量在覆盖作物翻压还田后通过矿化释放出来,既能提高土壤有机质含量,也能为后茬作物提供速效养分[29]。本研究结果表明,3种覆盖作物均能提高土壤碱解氮、速效磷和有效钾含量,土壤pH值和有机质含量也有增加趋势。与黑麦草和苕子翻压相比,混播翻压后,无论是否种植甜玉米,土壤养分均维持稳定,且与覆盖作物翻压后4 d相比,有增加的趋势。额外氮源的摄入可提高土壤碱解氮含量[23]。苕子的含氮量高于黑麦草,故苕子和混播翻压后的土壤碱解氮含量均高于黑麦草,且在覆盖作物翻压后21 d时苕子和混播处理无显著差异,但后续混播是否能持续保持优势,还有待进一步研究。
在本研究中,不同覆盖作物翻压后,无论是否种植甜玉米,土壤速效钾含量均显著提高。钾在土壤中主要以离子态存在,易溶于水,故释放较快[30]。苕子和混播翻压后,土壤有效磷含量整体呈波动增加的趋势,而黑麦草翻压后,土壤有效磷含量较对照显著降低。土壤对磷的吸附能力与有机酸有关,而不同覆盖作物在腐解过程中通过溶解、还原、酸溶等作用,使土壤中难溶的有机态磷转化为无机态磷[31]。同时,覆盖作物腐解过程中产生的有机酸种类和含量也能直接影响土壤有效磷含量[32]。有研究发现,土壤有效磷含量在前期低,有利于在玉米生育后期为其提供充足的氮素养分,且随着玉米后期利用氮素的能力提高,有利于玉米对于土壤速效钾及有效磷的吸收[33],表明与苕子和混播相比,黑麦草翻压后种植甜玉米可能更有利于增加二者之间氮素需求的协同性。
3.3 土壤养分与酶活性相关性
本研究結果表明,黑麦草翻压后仅土壤β-葡萄糖苷酶与土壤pH值呈极显著正相关,混播翻压后土壤β-葡萄糖苷酶活性与土壤碱解氮、有效磷含量均呈显著正相关,而苕子翻压后土壤β-葡萄糖苷酶与土壤养分未有显著相关性。土壤β-葡萄糖苷酶作为专一的纤维素水解酶,主要参与土壤中的碳循环过程[26]。本研究结果显示,无论是否种植甜玉米,苕子翻压后的土壤β-葡萄糖苷酶活性整体高于黑麦草、混播翻压,表明苕子翻压前期的外源碳可能多于黑麦草和混播翻压。碳源的摄入能显著提高土壤pH值[34],而pH值增加加快了碱性磷酸酶水解速率并抑制了土壤碳氮养分循环,从而间接抑制苕子翻压后的土壤β-葡萄糖苷酶活性[35]。也有可能是由于土壤pH值对土壤基质中固定的胞外酶活性有直接影响[36]。
脲酶作为专一的尿素水解酶,能催化尿素在土壤中的分解,促进植株吸收,可反映土壤的供氮能力[26]。单播苕子翻压后土壤脲酶活性与土壤pH值和有机质含量均呈显著正相关,与碱解氮含量呈显著负相关。苕子作为豆科植物,自身含氮量较高,前期腐解速率较快[37],翻压后改变了土壤微生物群落,增加了土壤脲酶活性。混播翻压后增强了土壤脲酶活性与土壤速效钾含量呈显著正相关,减少了由苕子翻压后土壤脲酶活性与土壤碱解氮的显著负相关影响。与苕子翻压相比,混播中有黑麦草提供的额外碳源,提高了土壤碳氮比,更有利于覆盖作物的腐解。在含有苕子的处理中,土壤脲酶活性与土壤有机质含量表现为显著正相关,这与和文祥等的研究结果[38]一致。土壤有机质通过影响全氮、速效氮等因素对脲酶活性存在较大的间接正效应[39],而脲酶在促进土壤氮素循环的同时,也间接地对土壤有机质造成一定的影响。
4 结论
覆盖作物翻压还田前期(覆盖作物翻压后 21 d)能够提高土壤β-葡萄糖苷酶活性。等量苕子较黑麦草翻压还田能够释放更多的无机氮,维持更高的土壤β-葡萄糖苷酶活性,但存在增加氮损失、氮淋溶的风险,而苕子和黑麦草混合种植翻压还田腐解有利于保持土壤中合理的碱解氮浓度和有效性时间,具有促进甜玉米氮素吸收、减少土壤氮素损失的潜力。从覆盖作物腐解养分释放与甜玉米养分吸收的协同性考虑,种植甜玉米后较苕子和黑麦草翻压相比,混播处理下的土壤β-葡萄糖苷酶活性、碱解氮和有效磷均维持稳定,表明混播翻压还田更有利于维持甜玉米对养分的需求。
参考文献:
[1]孙明雪,张玉霞,丛百明,等. 苜蓿抗寒性对秋季施用钾肥种类及用量的生理响应[J]. 干旱地区农业研究,2022,40(1):247-254.
[2]李青梅,王华玲,张玲玲,等. 白三叶草和鼠茅草对果园土壤微生物和线虫群落的影响差异[J]. 植物营养与肥料学报,2021,27(6):1055-1067.
[3]王晓维,徐健程,龙昌智,等. 施氮量和土壤含水量对黑麦草还田红壤氮素矿化的影响[J]. 植物营养与肥料学报,2018,24(2):365-374.
[4]曹卫东,包兴国,徐昌旭,等. 中国绿肥科研60年回顾与未来展望[J]. 植物营养与肥料学报,2017,23(6):1450-1461.
[5]Hunter M C,Kemanian A R,Mortensen D A. Cover crop effects on maize drought stress and yield[J]. Agriculture,Ecosystems & Environment,2021,311:107294.
[6]Langelier M,Chantigny M H,Pageau D,et al. Nitrogen-15 labelling and tracing techniques reveal cover crops transfer more fertilizer N to the soil reserve than to the subsequent crop[J]. Agriculture,Ecosystems & Environment,2021,313:107359.
[7]Adetunji A T,Ncube B,Mulidzi R,et al. Management impact and benefit of cover crops on soil quality:a review[J]. Soil and Tillage Research,2020,204:104717.
[8]Wen Y,Zang H D,Ma Q X,et al. Impact of water table levels and winter cover crops on greenhouse gas emissions from cultivated peat soils[J]. The Science of the Total Environment,2020,719:135130.
[9]Griffiths M,Delory B M,Jawahir V,et al. Optimisation of root traits to provide enhanced ecosystem services in agricultural systems:a focus on cover crops[J]. Plant,Cell & Environment,2022,45(3):751-770.
[10]蹇述蓮,李书鑫,刘胜群,等. 覆盖作物及其作用的研究进展[J]. 作物学报,2022,48(1):1-14.
[11]Glasener K M,Wagger M G,MacKown C T,et al. Contributions of shoot and root nitrogen-15 labeled legume nitrogen sources to a sequence of three cereal crops[J]. Soil Science Society of America Journal,2002,66(2):523-530.
[12]Luo L,Meng H,Gu J. Microbial extracellular enzymes in biogeochemical cycling of ecosystems[J]. Journal of Environmental Management,2017,197:539-549.
[13]Bowles T M,Acosta-Martínez V,Calderón F,et al. Soil enzyme activities,microbial communities,and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape[J]. Soil Biology and Biochemistry,2014,68:252-262.
[14]Yang K,Zhu J J,Yan Q L,et al. Soil enzyme activities as potential indicators of soluble organic nitrogen pools in forest ecosystems of Northeast China[J]. Annals of Forest Science,2012,69(7):795-803.
[15]Dilly O,Blume H P,Munch J C. Soil microbial activities in Luvisols and Anthrosols during 9 years of region-typical tillage and fertilisation practices in northern Germany[J]. Biogeochemistry,2003,65(3):319-339.
[16]Adetunji A T,Lewu F B,Mulidzi A R,et al. The biological activities of β-glucosidase,phosphatase and urease as soil quality indicators:a review[J]. Journal of Soil Science and Plant Nutrition,2017,17(3):794-807.
[17]鲍士旦. 土壤农化分析方法[M]. 北京:中国农业出版社,2000:25-108.
[18]关松荫. 土壤酶及其研究法[M]. 北京:北京农业出版社,1986:30-125.
[19]Nevins C J,Lacey C,Armstrong S D. The synchrony of cover crop decomposition,enzyme activity,and nitrogen availability in a corn agroecosystem in the Midwest United States[J]. Soil and Tillage Research,2020,197:104518.
[20]Hobbie J E,Hobbie E A. Microbes in nature are limited by carbon and energy:the starving-survival lifestyle in soil and consequences for estimating microbial rates[J]. Frontiers in Microbiology,2013,11(4):324-334.
[21]王理德,王方琳,郭春秀,等. 土壤酶学硏究进展[J]. 土壤,2016,48(1):12-21.
[22]de Notaris C,Mortensen E ?,S?rensen P,et al. Cover crop mixtures including legumes can self-regulate to optimize N2fixation while reducing nitrate leaching[J]. Agriculture,Ecosystems & Environment,2021,309:107287.
[23]徐 欣,王晓军,谢洪宝,等. 秸秆腐解对不同氮肥水平土壤脲酶活性的影响[J]. 中国农学通报,2018,34(34):99-102.
[24]蔡银美,张成富,赵庆霞,等. 模拟根系分泌物输入对森林土壤氮转化的影响研究综述[J]. 浙江农林大学学报,2021,38(5):916-925.
[25]薄晶晶,王 俊,付 鑫. 两种绿肥腐解及其碳氮养分释放动态特征[J]. 生态科学,2019,38(6):37-45.
[26]Nevins C J,Lacey C,Armstrong S. Cover crop enzyme activities and resultant soil ammonium concentrations under different tillage systems[J]. European Journal of Agronomy,2021,126:126277.
[27]Laidler B P H K. The molecular kinetics of the urea-urease system.Ⅱ. The inhibition by products[J]. Journal of the American Chemical Society,1950,72(6):2487-2489
[28]高菊生,黄 晶,杨志长,等. 绿肥和稻草联合还田提高土壤有机质含量并稳定氮素供应[J]. 植物营养与肥料学报,2020,26(3):472-480.
[29]高嵩涓,周国朋,曹卫东. 南方稻田紫云英作冬绿肥的增产节肥效应与机制[J]. 植物营养与肥料学报,2020,26(12):2115-2126.
[30]徐健程,王晓维,朱晓芳,等. 不同绿肥种植模式下玉米秸秆腐解特征研究[J]. 植物营养与肥料学报,2016,22(1):48-58.
[31]趙冬雪,王盼盼,常春丽,等. 绿肥套作对植烟土壤微生物群落功能多样性的影响[J]. 华北农学报,2019,34(5):201-207.
[32]顾炽明,李 越,李银水,等. 绿肥腐解液中有机酸组成对铝磷和铁磷活化能力的影响[J]. 植物营养与肥料学报,2021,27(9):1627-1635.
[33]刘诗璇,陈松岭,蒋一飞,等. 控释氮肥与普通氮肥配施对东北春玉米氮素利用及土壤养分有效性的影响[J]. 生态环境学报,2019,28(5):939-947.
[34]郑慧芬,吴红慧,翁伯琦,等. 施用生物炭提高酸性红壤茶园土壤的微生物特征及酶活性[J]. 中国土壤与肥料,2019(2):68-74.
[35]秦嘉海,张 勇,赵芸晨,等. 祁连山黑河上游不同退化草地土壤理化性质及养分和酶活性的变化规律[J]. 冰川冻土,2014,36(2):335-346.
[36]Sinsabaugh R L,Lauber C L,Weintraub M N,et al. Stoichiometry of soil enzyme activity at global scale[J]. Ecology Letters,2008,11(11):1252-1264.
[37]单明娟,秦 华,陈俊辉,等. 两种间作体系对丛枝菌根真菌侵染及多氯联苯去除的影响[J]. 应用与环境生物学报,2018,24(3):470-477.
[38]和文祥,谭向平,王旭东,等. 土壤总体酶活性指标的初步研究[J]. 土壤学报,2010,47(6):1232-1236.
[39]黄书涛. 黄河三角洲盐碱地土壤脲酶活性与土壤性质的通径分析[J]. 山东农业科学,2007(6):89-91.
猜你喜欢
红领巾·探索(2021年2期)2021-08-26 15:02:02
学生天地(2020年34期)2020-06-09 05:50:36
军事文摘·科学少年(2020年2期)2020-03-19 08:48:18
当代陕西(2019年11期)2019-06-24 03:40:26
江苏农业科学(2016年8期)2017-02-15 20:30:50
农民致富之友(2016年22期)2017-02-07 01:46:19
作文通讯·高中版(2017年12期)2017-02-06 05:56:42
河北渔业(2015年5期)2016-03-01 16:21:29
天津农业科学(2015年12期)2015-12-03 00:02:50
湖北农业科学(2015年16期)2015-10-28 21:29:11
