
生殖生长期遮光处理对大豆叶片光合生理和衰老的影响
2023-12-01 06:30姚兴东王小凡檀卓芮何德鑫孙贺祥谢甫绨
沈阳农业大学学报 2023年5期
姚兴东,王小凡,檀卓芮,何德鑫,孙贺祥,谢甫绨
(沈阳农业大学农学院,沈阳 110161)
大豆是我国重要的粮食作物和油料作物,在我国粮食安全中占有重要地位。2022年我国消费大豆11 529万t,其中自产仅为2 028万t,严重依赖进口。但我国耕地面积有限,直接改种大豆会影响玉米等其他作物的供应。大豆玉米带状复合种植改单一作物种植为高低作物搭配间作、改等行种植为大小垄种植,充分发挥边行优势,提高作物群体光能利用率,实现玉米产量基本不减、增收一季大豆[1],该技术是缓解我国大豆自己能力不足的有效手段。但该模式中高秆玉米的遮光会改变矮秆大豆冠层的光环境,导致大豆产量下降[2]。因此,研究遮光对大豆生长发育及产量形成的影响对于提高大豆玉米带状复合种植下大豆产量具有重要意义。
遮光会引起大豆植株的形态性状和生理生化过程产生一定的变化,最终导致产量的下降。遮光会引起大豆株高增高,茎粗变细,各个器官干物质积累量明显减少,进而导致产量下降,且在大豆生长期内遮光时间越长,大豆单株产量越低[3]。遮光会引起大豆叶片光合能力下降,导致干物质积累的下降[4]和产量的降低[5]。玉米大豆带状间作种植时,营养生长阶段玉米对大豆的遮光效果不明显;随着玉米的生长,玉米对大豆的遮光胁迫逐渐加重,在大豆开花期玉米对大豆产生明显的荫蔽效应。大豆开花后进入生殖生长阶段,该时期是大豆产量形成最重要的时期,因此,研究大豆生殖生长阶段对遮光的响应对于深入理解大豆对遮光的响应规律具有重要意义。
叶片衰老的启动时间和衰老速度会直接影响作物的光合时间和光合有效面积,进而影响光合产物的合成与积累,最终决定了作物的产量[6]。叶片衰老过程中,细胞内活性氧的积累导致膜脂过氧化程度增加,进而破坏了生物膜结构和功能,引起衰老。而细胞内活性氧的产生和清除与抗氧化酶活性密切相关[7]。叶片的衰老存在基因型差异,同时还受遮光等环境变化的影响。本试验采取盆栽试验,设置人工遮光处理,探讨生殖生长期遮光对大豆光合生理和叶片衰老的影响,为提高大豆玉米带状复合种植条件下大豆的产量和选育耐荫品种提供理论依据。
1 材料与方法
1.1 材料
试验于2017年5~10月在沈阳农业大学教学科研基地进行。供试大豆品种为耐荫品种辽豆32和不耐荫品种辽豆29,供试大豆品种特性如表1。采用遮阳网进行人工遮光处理,不同遮阳网处理下光通量密度如图1。
1.2 方法
采用裂区试验设计,主区为3个遮光处理(自然光、人工遮光50%和人工遮光70%处理,使光照强度分别为自然光照、自然光照的50%和自然光照的30%;分别记为S0、S50 和S70),在始花期(7 月11日)通过覆盖透光率不同的遮阳网进行遮光(不同遮光处理下光通量密度如图1)。副区为不同大豆品种。
采用盆栽试验,盆钵直径32 cm,盆高40 cm,试验用土在装盆前晒干并过筛。每次处理设15 次重复。播种前对种子进行精选,每盆播种4粒,播种深度3 cm,每盆留苗2株,常规管理。
1.2.1 光合特性测定 于始荚期、鼓粒期、鼓粒末期,使用便携式叶面积仪(LI-3100C,LI-COR,USA)测定单株叶面积。
于始荚期、鼓粒期、鼓粒末期取植株主茎顶端倒数第3 片叶,将叶片切片处理,称取0.1 g 叶片放入5 mL 95%乙醇溶液中充分浸泡,黑暗条件下浸提48 h。取浸提液后以95%乙醇溶液为对照,分别测量在663 nm 和645 nm波长下的光密度。通过公式计算溶液中叶绿素a、叶绿素b、总叶绿素的含量,及叶绿素a/b的值。
选择晴天9∶00-1∶00,利用LI-6400 便携式光合测定系统(美国,LI-COR 公司)测定植株光合速率,固定红蓝光源叶室,光合有效辐射(PAR)设定为1 000 μmol·m-2·s-1,于始荚期、鼓粒期、鼓粒末期测定大豆主茎顶端倒数第3片复叶片中间小叶(避开叶脉)的净光合速率,每盆测2株,每处理测3盆(3次重复)。
使用FMS便携脉冲调制式荧光仪于始荚期、鼓粒期、鼓粒末期对大豆主茎顶端倒数第3片复叶片中间小叶(避开叶脉)进行测定,重复3 次。将植株置于暗室暗适应30 min, 分别测定叶片的初始荧光(F0)、最大荧光(Fm);在光处理30 min 下测定光下稳定荧光(Fs)、最大荧光最大光(Fm')和最小荧光(F0')。通过公式计算最大化学效率Fv/Fm=(Fm-F0)/Fm,PSⅡ实际光化学效率ΦPSⅡ=(Fm'-Fs)/Fm'和光化学淬灭系数qP=(Fm'-Fs)/(Fm'-F0')。
1.2.2 叶片抗氧化酶活性测定 分别于大豆始荚期、鼓粒期、鼓粒末期取大豆主茎顶端倒数第3片叶,在低温环境下称取0.2 g叶片,并将其用液态氮于研钵中研磨,放入离心管中加入磷酸二氢钾溶液,低温下以10 000 r·min-1离心5 min,吸取上清液低温保存。
过氧化氢酶(CAT)采用紫外吸收法测定[9],过氧化物酶(POD)采用愈创木酚法测定[8],超氧化物歧化酶(SOD)使用硝基蓝四氮唑(NBT)光化学还原法测定[9]。
1.2.3 叶片丙二醛(MDA)含量测定 分别于大豆始荚期、鼓粒期、鼓粒末期取大豆主茎顶端倒数第3片叶,在低温环境下称取0.2 g叶片,液氮中研磨后加入磷酸二氢钾溶液,用通过硫代巴比妥酸计时法测定MDA含量[8],
1.3 数据处理方法
试验所得数据结果采用Microsoft Excel 2013 和DPS 7.05 进行统计分析,处理间差异用LSD 法比较,用Origin 2021作图。
2 结果与分析
2.1 生殖生长期遮光处理对大豆光合特性的影响
2.1.1 单株叶面积 在生殖生长阶段,大豆的单株叶面积均随着生育进程的推进逐渐减少(图2A)。遮光显著的降低了大豆的单株叶面积,且降低幅度随着遮光程度的增加而变大。随着遮光时间的增加,遮光对单株叶面积的影响变大。在结荚期,自然光处理下辽豆32的单株叶面积与50%遮光处理下差异不显著,但显著的高于70%遮光处理;而不同处理间辽豆29的单株叶面积均表现显著差异。在鼓粒末期,两大豆品种的单株叶面积在各处理间均表现为差异显著,且在相同遮光处理下,辽豆29的单株叶面积下降幅度均大于辽豆32的。在生殖生长阶段,大豆的光合势均随着生育进程的推进逐渐减少(图2B)。生殖生长阶段遮光显著降低了大豆的光合势,且随着遮光程度的增加下降幅度变大。相同遮光处理下,辽豆32的光合势均显著高于辽豆29的。
2.1.2 叶绿素含量 在生殖生长阶段,大豆叶片的叶绿素a、叶绿素b和总叶绿素含量均随着生育进程的推进呈下降的趋势,遮光会增加大豆叶片的叶绿素含量,且随着遮光程度的增加大豆叶绿素含量增加幅度变大(图3)。在同一生育时期,辽豆32叶片中叶绿素a、叶绿素b和总叶绿素含量均大于辽豆29。大豆叶片叶绿素a/b比值随着生育期进程的推进呈先上升后下降的趋势,在鼓粒期达到最大值。遮光降低了大豆叶片的叶绿素a/b比值。
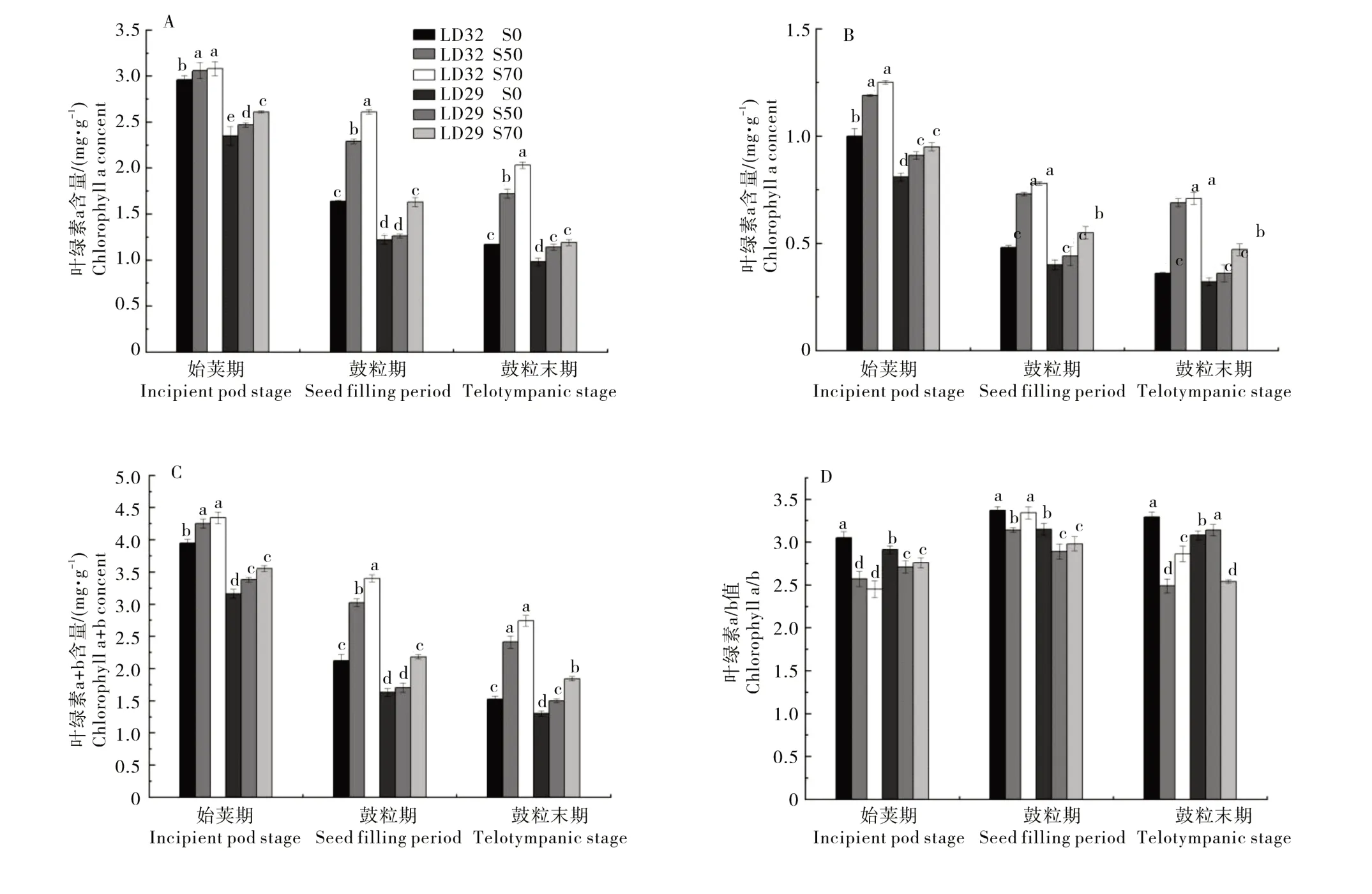
图3 不同遮光处理下大豆品种生殖生长期的叶绿素a(A)、叶绿素b(B)和叶绿素a+b(C)含量,及叶绿素a/b(D)Figure 3 Chlorophyll a (A), chlorophyll b (B), chlorophyll a+b (C) contents, and chlorophyll a/b (D) of different soybean cultivars at reproductive growth stages under different shading treatments
2.1.3 光合速率 在生殖生长阶段,大豆叶片的光合速率随着生育进程的推进呈下降的趋势,遮光降低了大豆叶片的光合速率,且随着遮光程度的增加大豆叶片的光合速率的下降幅度变大。同一遮光处理下,辽豆32叶片的光合速率高于辽豆29(图4)。

图4 不同遮光处理下不同品种大豆生殖生长期的光合速率Figure 4 Photosynthetic rate of different soybean cultivars at reproductive growth stages under different shading treatments
2.1.4 荧光参数 在生殖生长阶段,大豆叶片的初始荧光强度(F0)、最大光化学效率(Fv/Fm)、PSⅡ实际光化学效率(ΦPSⅡ)和叶绿素荧光光化学淬灭系数(qP)值均随着生育进程的推进呈下降的趋势(图5)。遮光降低了大豆叶片的F0、ΦPSⅡ和qP值,且随着遮光程度的增加下降幅度变大,而Fv/Fm呈相反的趋势。遮光条件下,辽豆32的F0、ΦPSⅡ和qP值均要高于辽豆29。Fv/Fm呈现相反的趋势,即遮光提高了叶片的Fv/Fm值,其随着遮光程度的增加升高幅度变大。
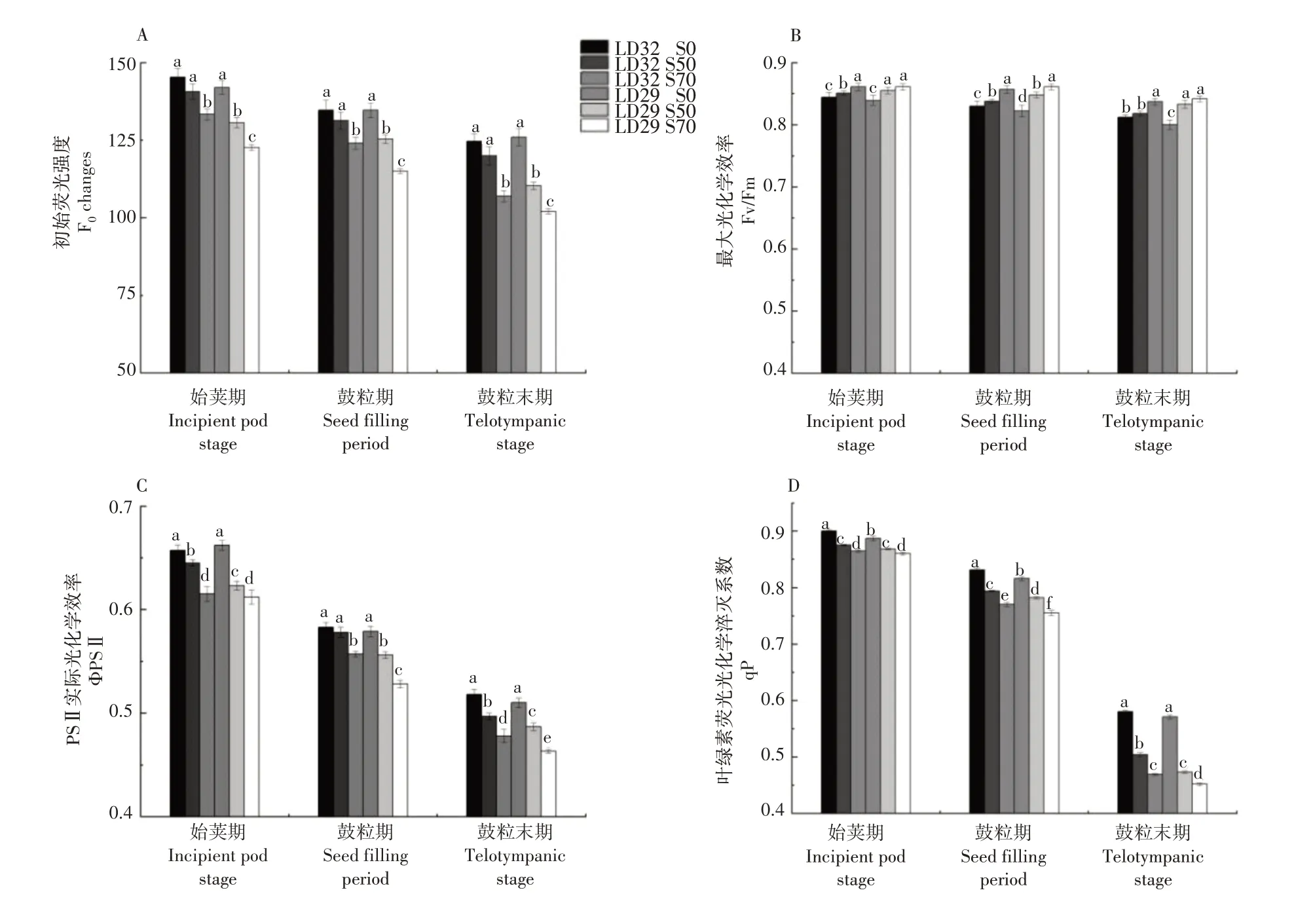
图5 不同遮光处理下不同品种大豆生殖生长期的初始荧光强度(A)、最大光化学效率(B)、PSⅡ实际光化学效率(C)和叶绿素荧光光化学淬灭系数(D)Figure 5 F0 changes (A),Fv/Fm(B),ΦPSⅡ(C),qP (D)of different soybean cultivars at reproductive growth stages under different shading treatments
2.2 生殖生长期遮光处理对大豆叶片抗氧化酶活性的影响
在生殖生长阶段,2个品种大豆叶片的POD、SOD 和CAT酶活性均随着生育进程的推进呈先增加后减少的趋势(图6)。遮光增加了大豆叶片的POD、SOD 和CAT 酶活性。在始荚期,大豆叶片的POD、SOD 和CAT 酶活性均随着遮光程度的增加而增加。在鼓粒期和鼓粒末期,50%遮光处理下大豆叶片的POD、SOD 和CAT 酶活性高于70%和未遮光处理。同一处理下,辽豆32叶片的POD、SOD和CAT酶活性要低于辽豆29。

图6 同遮光处理下不同品种大豆生殖生长期的叶片过氧化氢酶(A)、过氧化物酶(B)和超氧化物歧化酶(C)活性Figure 6 CAT activity (A), POD activity (B) and SOD activity (C) in leaves of different soybean cultivars at reproductive growth stages under different shading treatments
2.3 生殖生长期遮光处理对大豆叶片MDA含量的影响
在生殖生长阶段,大豆叶片的MDA 含量均随着生育进程的推进逐渐增加(图7)。遮光显著的增加了大豆叶片的MDA含量,且增加幅度随着遮光程度的增加而变大。随着遮光时间的增加,遮光对叶片MDA含量的影响逐渐变大。不同品种叶片的MDA 含量对遮光处理的响应不同。在鼓粒末期,50%遮光处理下辽豆32 的显著低于辽豆29,但在75%遮光处理下2个品种叶片的MDA含量差异不显著。
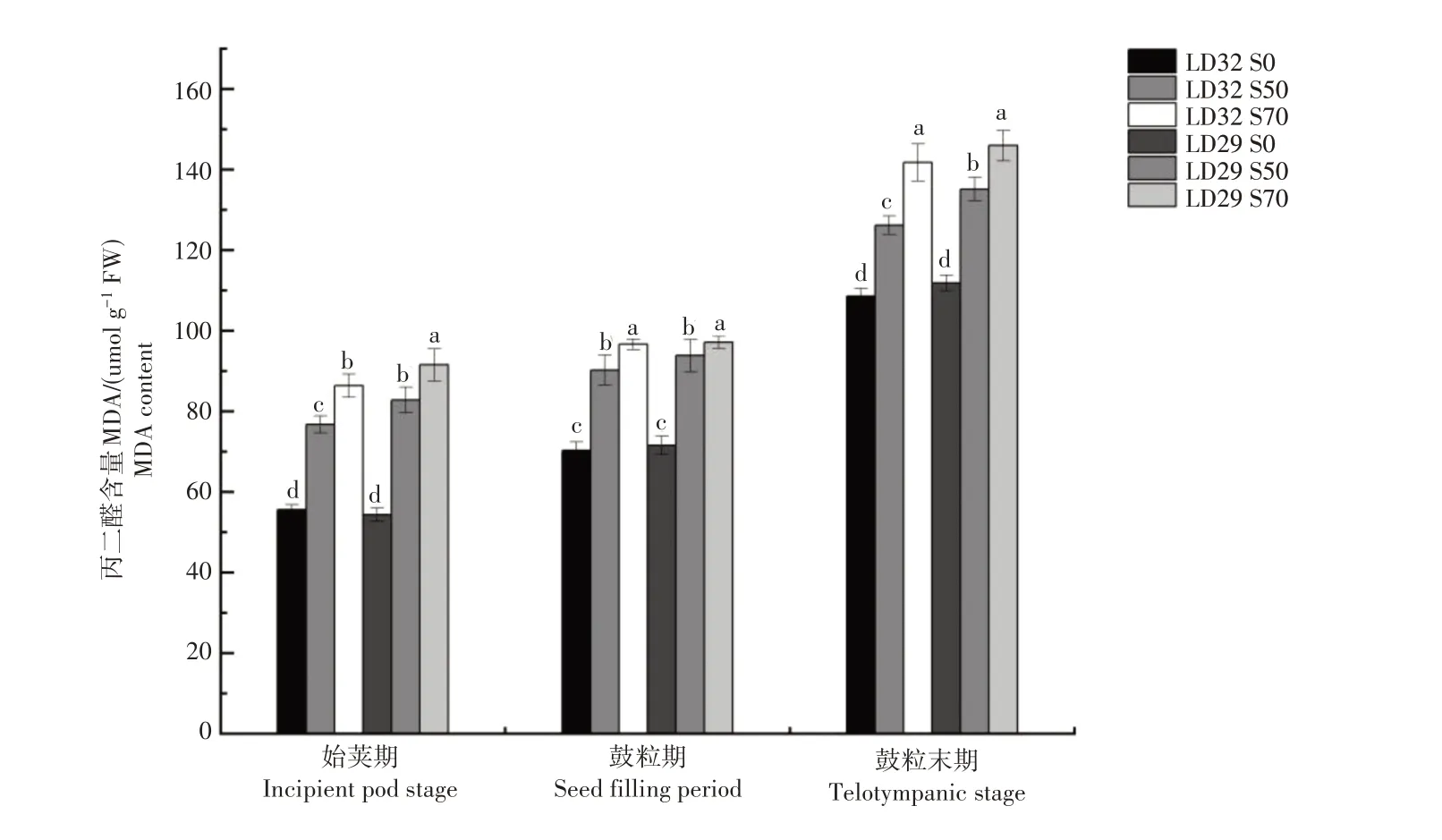
图7 不同遮光处理下不同品种大豆生殖生长期的叶片丙二醛含量Figure 7 MAD content in leaves of soybean varieties at different growth stages under different shading treatments
3 讨论与结论
3.1 遮光对大豆单株叶面积和叶片光合生理的影响
叶片是植物进行光能截获、吸收和转化的主要器官,叶片的大小和形状直接影响植物对光能的吸收[10]。单株叶面积是决定产量的重要性状,叶面积减少会限制作物对光能的截获,减少干物质的积累[11]。延长生殖生长期大豆叶面积的持续时间可增加干物质生产[12],提高产量[13]。任万军等[14]研究表明,弱光条件下大豆叶片会变长、变宽进而获得更多的光能,但叶片会变薄。龚万灼等[15]研究表明玉米大豆带状套作遮光会显著降低大豆的单株叶面积。汪直华等[16]研究了苗期人工遮光对大豆的影响,结果表明人工遮光会显著降低大豆的单株叶面积,且受影响程度随着遮光强度的增加而增加。本试验的研究表明,生殖生长期遮光显著的降低了大豆的单株叶面积和光合势,且降低幅度随着遮光程度的增加而增加。在相同的遮光处理下,辽豆32 的单株叶面积和光合势均要高于辽豆29,这有利于其截获更多的光能。生殖生长阶段,大豆植株光合积累的有机物和营养物质主要运输到生殖器官,同时遮光限制了大豆叶片的光合作用,造成库大源小,进一步促进叶片中的干物质和养分转运到营养器官,导致叶片衰老。
作物叶片内叶绿素可以起到传递和转化光能的作用,其组成和含量会直接影响叶片的光合作用。叶绿素a和叶绿素b是叶绿素中两个重要的组成部分,叶绿素参与光合作用中光能的吸收、传递和转化,是植物进行光合作用重要的物质[17]。叶绿素b 又被称为荫生叶绿素[18],它能够良好地利用漫反射中的蓝紫光,在遮光条件下蓝光会增加,叶绿素b 的增加更有利于植株对光能的捕获,促进植株的生长。在遮光处理下,大豆叶片内的叶绿体结构及色素的形成,含量和分布状况会随之改变,叶绿素a、叶绿素b 和叶绿素a+b 的含量及叶绿素a/b 的比值会受到改变[19]。本试验结果表明,遮光显著的增加了叶片中叶绿素a、叶绿素b 和叶绿素a+b 的含量,显著降低了叶片中叶绿素a/b的比值。说明遮光下,大豆通过提高叶绿素的含量获取更多的光能[20]。植株受到弱光胁迫后叶绿素b 含量的增加幅度大于叶绿素a 含量的上升幅度,因此叶绿素a/b 的比值呈现下降趋势[21]。在同一生育时期,辽豆32叶片中叶绿素a、叶绿素b和总叶绿素含量均大于辽豆29。说明耐荫品种对弱光的获取能力更强。
光合速率的快慢代表植株制造碳水化合物能力的强弱,是植株产量高低的决定性因素。汪扬媚等[22]研究表明,苗期人工遮光会导致大豆叶片光合速率的显著下降,且下降幅度随着遮光程度的增加而增加。陈吉玉等[23]研究了2~4叶期遮光对大豆光合生理的影响,结果表明,轻度遮光(遮光20%)和中度遮光(遮光40%)下大豆叶片的光合速率高于对照,重度遮光(遮光75%)下的光合速率显著低于对照。本研究结果表明,在生殖生长阶段,遮光降低了大豆叶片的光合速率,且随着遮光程度的增加大豆叶片的光合速率的下降幅度变大。同一遮光处理下,辽豆32 叶片的光合速率高于辽豆29。说明遮光抑制了大豆叶片的光合作用,耐荫品种在遮光下的光合作用能力更强。
叶绿素荧光动力学参数能够快速和准确地反映出植株内部光合作用中对光的吸收、传递、分配和耗散的情况。遮光会导致小麦旗叶的初始荧光F0和最大光化学效率Fv/Fm增加,但PSⅡ光化学反应中心的实际光化学效率ΦPSⅡ和光化学荧光猝灭系数qP 下降[24]。本试验研究结果表明,遮光降低了大豆叶片的F0、ΦPSⅡ和qP值,且随着遮光程度的增加下降幅度变大,而Fv/Fm呈相反的趋势。初始荧光F0的降低表明在光反应中非光化学能量耗散增加,PSⅡ反应中心受到损伤。最大光化学效率Fv/Fm的升高,加大了PSⅡ原初光能转化效率,但遮光降低了ΦPSⅡ和qP,表明PSⅡ光化学反应中心活性受到了遮光的影响,遮光条件下大豆叶片用于光化学反应的光能捕获效率下降,非光化学能量耗散升高,因此为暗反应提供的同化物能量减少,尽管遮光处理的大豆叶片叶绿素含量增加,但最终还是导致了大豆叶片光合速率的明显下降。耐荫品种辽豆32的荧光参数受遮光影响的变化幅度低于不耐荫品种辽豆29,这表明不耐荫品种受光环境改变表现更敏感。
3.2 遮光对大豆叶片衰老的影响
叶片衰老的启动时间和衰老速度会直接影响作物的光合时间和光合有效面积,进而影响光合产物的合成与积累,最终决定了作物的产量[25]。在衰老过程中,叶片会发生光合能力减弱,叶绿素逐渐分解,绿叶面积减小,细胞膜结构遭到破坏等生理生化现象[26]。植物体内的抗氧化系统可以防御由活性氧积累带来的损伤并维持细胞内氧化还原的平衡,进而维持正常的呼吸、光合等代谢功能,对抵抗作物的衰老具有重要的作用[27]。植物进入衰老阶段,植物体内的活性氧(ROS)和丙二醛(MDA)含量增加,植物通过提高SOD、POD和CAT等抗氧化酶的活性,保持ROS产生与清除机制的平衡,降低MDA的含量[28]。SOD催化超氧化物的歧化反应,产生H2O2和氧气。CAT 可以将H2O2分解为H2O 和氧气,而POD 在植物衰老前期可以清除H2O2,防止衰老[29]。遮光显著提高了大豆叶片内MDA、H2O2的含量及O2-的产生速率,加剧了细胞膜的过氧化程度。本试验中,随着生育进程的推进,大豆叶片的MDA含量逐渐增加,同时光合势逐渐下降,表明大豆叶片逐渐衰老。遮光显著的增加了大豆叶片的MDA 含量,且增加幅度随着遮光程度的增加而变大。MDA 含量的增加表明大豆叶片膜脂过氧化程度加剧,叶片衰老加剧。遮光会提高大豆叶片中的SOD、CAT 和POD 活性,进而缓解叶片的衰老。本试验中,遮光提高了大豆叶片的SOD、CAT 和POD 活性。短期遮光(始荚期)处理,SOD、CAT 和POD 活性随着遮光程度的增加而增加,长期遮光(鼓粒期)处理,70%遮光处理的SOD、CAT 和POD 活性低于50%遮光处理的。说明随着遮光时间的增加,70%遮光处理的抗氧化酶系统已不能缓解遮光胁迫对叶片的影响,进而导致叶片的衰老。在鼓粒末期,50%遮光处理下耐荫品种辽豆32的MDA含量低于不耐荫品种辽豆29,但CAT和SOD酶活性均显著高于不耐荫品种辽豆29,表明耐荫品种的叶片衰老要慢于不耐荫品种,延长了其光合时间,有利于获得更高的产量。
综上所述,生殖生长期遮光显著增加大豆叶片的丙二醛含量和抗氧化酶活性,显著降低了光合势和光合能力,表明生殖生长期遮光促进了大豆叶片的衰老。耐荫品种具有更强的光合能力和较低的叶片衰老速度,这有利于其在遮光条件下获得更高的产量。
猜你喜欢
云南化工(2021年9期)2021-12-21
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
中国生殖健康(2020年8期)2021-01-18
中国生殖健康(2020年2期)2021-01-18
新疆农业科学(2020年1期)2020-02-14
中国生殖健康(2018年1期)2018-11-06
中国生殖健康(2018年2期)2018-11-06
中国麻业科学(2015年5期)2015-12-28
影像科学与光化学(2014年5期)2014-03-11
无机化学学报(2014年3期)2014-02-28
