
牡丹SHMT基因的鉴定与生物信息学分析*
2023-11-27 03:50周广灿刘夏婷王博杜晶晶
菏泽学院学报 2023年5期
周广灿,刘夏婷,王博,杜晶晶
(菏泽学院农业与生物工程学院(牡丹学院),山东 菏泽274015)
0 引言
丝氨酸羟甲基转移酶(Serine hydroxymethyltransferase,SHMT) 是一种磷酸吡哆醛(PLP)依赖酶,在细胞的一碳代谢途径中通过参与转氨基和脱羧反应,催化丝氨酸和甘氨酸的相互转化以发挥作用[1-3].作为参与基础代谢的重要酶类,SHMT在原核生物中多以二聚体形式存在,而在动物和真菌中则以四聚体形式存在[4].在拟南芥基因组中有7个SHMT基因,分别为AtSHM1~AtSHM7,其中SHM1与SHM2编码的蛋白质产物定位在线粒体基质中,其他5个SHMT基因编码的蛋白质缺乏可识别的定位序列,推测其存在于细胞质中[5].
除为生物合成作用提供一碳单位外,许多研究表明SHMT蛋白在植物抗病中也发挥重要功能[6-9].例如,拟南芥AtSHM1基因发生突变后,突变株易受到不同病原菌的感染[7].栽培大豆(GlycinemaxMerr.)中一个命名为Rhg4的SHMT基因的抗性等位基因具有两个重要的点突变(P130R和N358Y),影响了大豆根细胞内的叶酸稳态,最终限制胞囊线虫的生长,从而具有抵抗大豆胞囊线虫的功能[9].因此,对于植物中SHMT基因的研究可以揭示其功能,并为相关应用提供理论指导.
牡丹是一种重要的观赏植物,在我国多地均有种植.其中,菏泽、洛阳、北京、临夏、彭州、铜陵县等地区牡丹种植面积较大.牡丹不仅可以用于观赏,还具有丰富的食用和药用价值[10].然而,如今牡丹的病虫害问题日益突出,这直接影响了牡丹的生长和品质,同时也增加了种植者的管理难度和经济成本,已经严重影响了牡丹的产业化发展.因此,应加强对牡丹病虫害的监测和防治,以保障牡丹产业的健康发展.在危害牡丹产业化生产的诸多病虫害中,牡丹根结线虫病是主要病害之一,在北京、菏泽的牡丹栽培区均有分布[11-12].该病的病原是北方根结线虫,主要危害牡丹根部.染病植株除出现根结之外,其地上部分也会出现生长滞慢、植株矮小,甚至整株枯死的现象.该病害严重影响了牡丹的开花率和牡丹皮的产量与质量[13].
基于目前已知的大豆中SHMT基因具有抵抗大豆胞囊线虫的功能,根据同源基因的功能相似推测,牡丹中该类基因也可能具有抵抗牡丹根结线虫病的功能,但目前还未见有牡丹SHMT基因的相关研究.因此,本研究旨在借助生物信息学方法鉴定和分析牡丹基因组中该基因家族的数量与理化性质,并进一步分析演化关系,以揭示其功能,从而为防治牡丹根结线虫病等实际应用提供理论指导.
1 材料与方法
1.1 SHMT编码基因的鉴定
首先,在牡丹基因组网站(https://ftp.cngb.org/pub/CNSA/data5/ CNP0003098/ CNS0560369/ CNA 0050666/)下载牡丹基因组数据,在BioEdit软件(Version 7.2.5)中构建本地的牡丹蛋白质数据库.以拟南芥SHMT基因编码的氨基酸序列为查询序列,用BLAST检索获得牡丹基因组中的同源序列.然后使用 Pfam (https://www.ebi.ac.uk/interpro/)网站分析所得候选序列,以确定候选序列中是否存在 SHMT 结构域.最后,利用 NCBI BLASTP (https://blast.ncbi.nlm.nih.gov/Blast.cgi)和CD Search (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)在线网站进一步验证 SHMT结构域的完整性.
1.2 SHMT基因编码蛋白质理化性质及基因结构分析
在ExPasy(https://web.expasy.org/protparam)网站对牡丹SHMT 基因家族成员的氨基酸序列的理化性质进行分析预测,并通过在线网站WoLF PSORT(https://wolfpsort.hgc.jp)对牡丹SHMT 基因进行亚细胞定位预测.利用牡丹基因库gff文件与牡丹SHMT家族的ID号通过TBtools软件对牡丹SHMT基因家族成员进行基因结构分析并进行可视化绘图.
1.3 SHMT基因的染色体定位分析
根据牡丹基因组的物理位置,利用牡丹gff文件和SHMT基因家族的ID通过TBtools软件的Gene location visualize from GTF/GFF绘制牡丹SHMT基因的染色体位置示意图.
1.4 SHMT基因启动子顺式元件分析
为预测和揭示牡丹SHMT基因的功能,通过Plant CARE(https://bioinformatics.psb.ugent.be/webtools/plantcare/html/)网站预测分析牡丹SHMT基因的启动子顺式作用原件.首先,采用 TBtools 软件的GXF Sequences Extract提取牡丹基因组中起始密码子ATG上游2 000 bp基因序列作为启动子区域;进一步通过TBtools软件的Fasta Extract提取牡丹SHMT基因的启动子区域.将提取到的启动子区域提交至Plant CARE网站进行预测分析,并将结果用 TBtools 软件的Basic Biosequence View进行可视化分析.
1.5 SHMT基因的系统发育分析
为了分析牡丹SHMT基因的演化特征,以拟南芥SHMT基因作为参考,使用MEGA5.1软件,用ML法(最大似然法)构建进化树.首先,利用整合在MEGA5.1里的Clustal W软件对牡丹SHMT基因和拟南芥SHMT基因编码的氨基酸序列进行align排序,手动校正,建树时Bootstrap method数值设定为1 000,其余参数为默认值,建立演化关系树.通过建立的进化树分析以揭示牡丹SHMT基因在系统发育进化过程中的演化特征.
2 结果与分析
2.1 SHMT基因家族特征分析
在牡丹基因组中共鉴定到33条SHMT基因,这些基因编码的蛋白质在氨基酸数量、分子量大小、等电点、不稳定系数等理化性质方面均存在较大差异(见表1).该家族成员编码的蛋白质含有69~603个不等的氨基酸,分子量大小为7.52~66.90 kD,理论等电点处于4.90~10.45之间,大多数pI值为碱性.该家族中含有20个稳定性蛋白(不稳定系数<40),13个不稳定性蛋白.亚细胞定位预测结果显示,近半数(15个)成员都定位在细胞质中,其余成员分布在叶绿体、线粒体、细胞质膜等位置,不具有明显规律性.
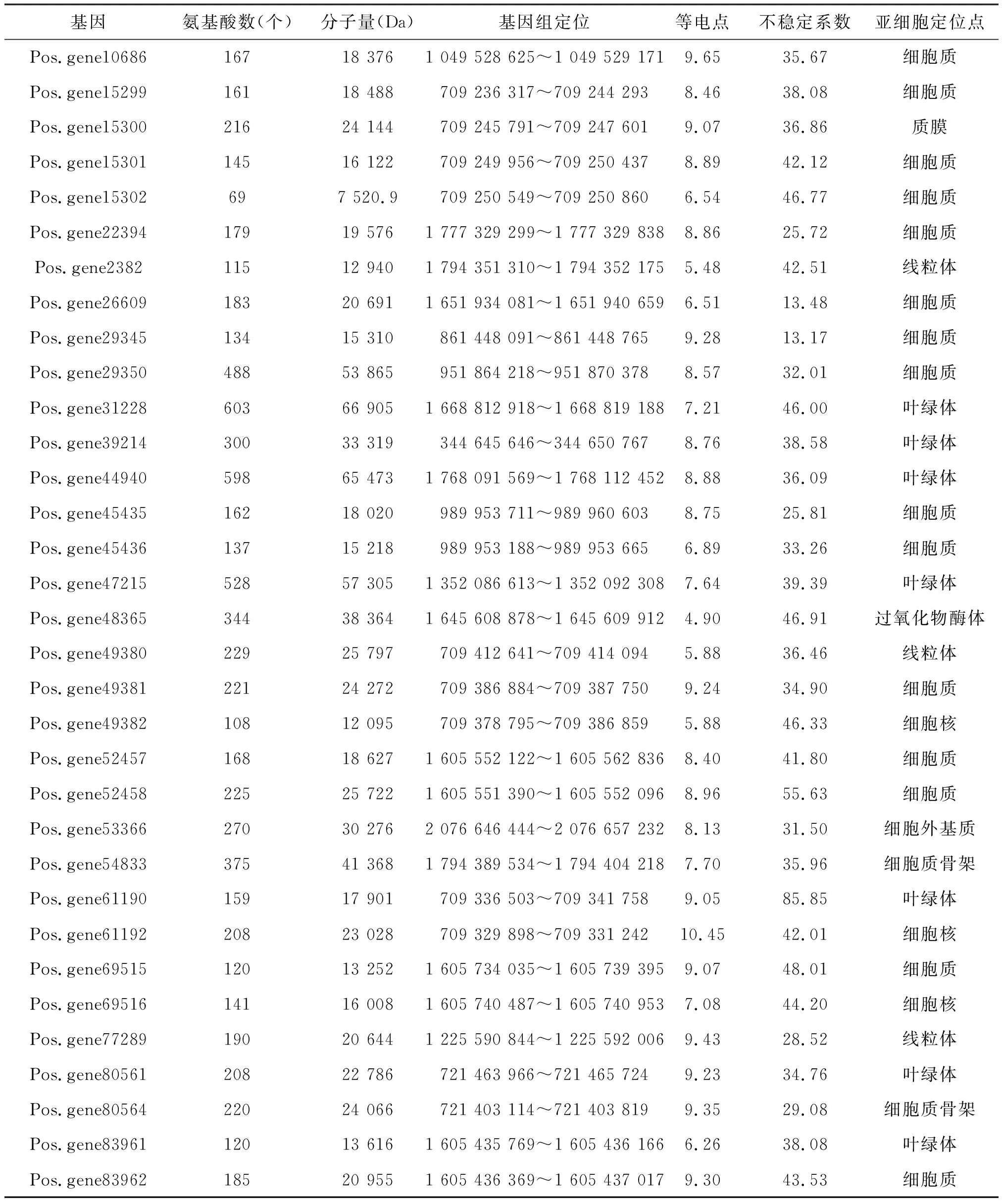
表1 牡丹SHMT基因理化性质分析
2.2 SHMT基因结构及编码蛋白质保守氨基酸基序
通过 MEME 在线网站预测了20个保守氨基酸基序,将预测结果与使用牡丹SHMT蛋白的氨基酸序列构建的系统发育树整合后绘制保守基序分析图.结果如图1所示,可根据系统发育树将牡丹SHMT蛋白分成3个亚群.第Ⅰ亚群中包含19个成员,其中有7个成员特异性含有motif15,3个成员特异性含有motif18.第Ⅱ亚群仅有1个成员(Pos.gene52457),它仅含有motif2和motif6两个保守基序.第Ⅲ亚群中有13位成员,其中Pos.gene44940与Pos.gene47215的motif组成较为类似.
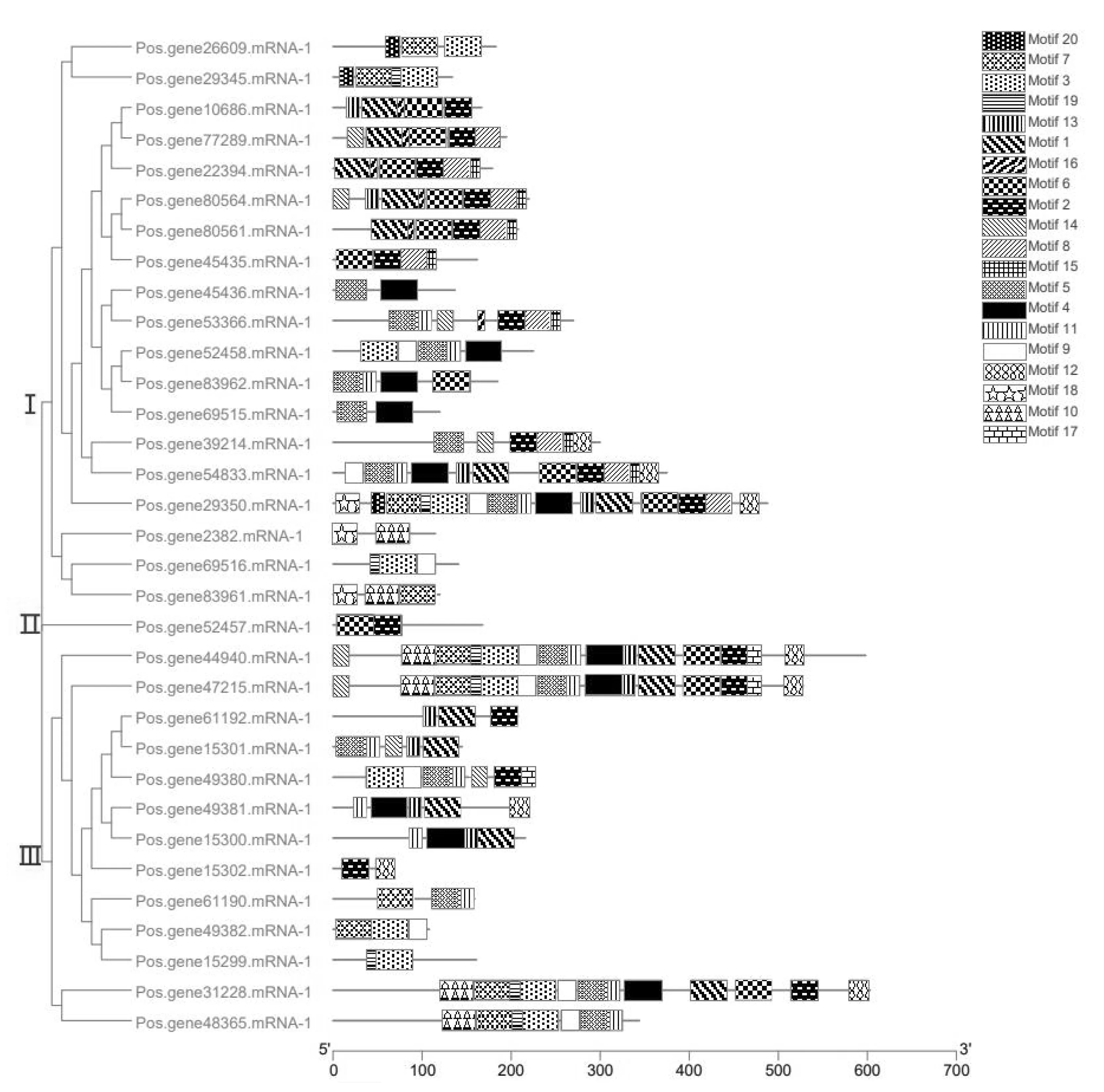
图1 牡丹SHMT基因保守氨基酸基序可视化图
此外,牡丹SHMT基因编码的蛋白质所含保守氨基酸基序种类较多,其中同一亚群的成员具有相似的保守氨基酸基序,而不同亚群中保守氨基酸基序种类差异较大.这些数据提示同一亚群成员功能的相似性和不同亚群间的功能差异性,即同一亚群的成员具有相似的生物学功能,而不同亚群的成员可能有不同的生物学功能.该结果对于通过牡丹SHMT基因家族保守基序来研究这些基因的功能及其在进化中的作用具有重要意义.
基因结构可视化结果如图2所示,除第Ⅰ亚群的Pos.gene22394与第Ⅲ亚群的Pos.gene48365基因只含有1个外显子,其余成员均含两个或多个外显子.其中,Ⅰ亚群的的Pos.gene29350基因含有14个外显子,Ⅲ亚群的Pos.gene44940基因含有13个外显子.

图2 牡丹SHMT基因结构比较图
2.3 SHMT基因的染色体定位分析
SHMT基因染色体定位结果显示,所有成员均能定位到牡丹的5条染色体上,但在染色体上的分布不均匀(见图3).其中,1号染色体和3号染色体仅分布1个成员,2号染色体和5号染色体上各有5个成员,4号染色体上的成员数最多,为21个.

图3 牡丹SHMT基因的染色体定位图
2.4 SHMT基因启动子顺式元件分析
为推测牡丹SHMT基因的功能,对其启动子顺式作用元件进行功能预测分析.结果如图4所示,SHMT基因启动子中存在众多参与植物应激反应、生长发育的作用元件.在参与激素应答方面,SHMT基因中有10个成员可以对生长素作出反应,15个成员可以对赤霉素作出反应,16个成员对水杨酸作出反应,21个成员可以对茉莉酸甲酯作出反应,还有28个成员对脱落酸作出反应.由于各成员所含激素应答的元件种类和数量均有差异,表明SHMT基因家族在牡丹的不同激素响应过程中可能有着重要作用.此外,牡丹SHMT基因启动子中还包含多种与非生物胁迫因素相关的元件.如:有21个成员含有厌氧诱导元件,11个成员含有防御胁迫元件,14个成员含有低温响应元件,19个成员含有干旱诱导元件等,表明该家族成员在胁迫应答中也发挥着重要作用.在植物生长发育方面,SHMT基因家族中有6个成员含有细胞周期调控元件,2个成员含有胚乳表达元件,24个成员含有分生组织表达相关元件,2个成员含有细胞分化调节元件等.值得注意的是,SHMT基因家族中有3个成员还含有与黄酮类生物合成相关的基因调控元件,推测这些基因成员在黄酮类生物合成方面发挥着重要作用.在该家族中的诸多作用元件中,光反应元件占比最多,除Pos.gene49382基因以外,其余32个成员均具有一个或多个光反应作用元件.上述结果表明,调控SHMT基因家族成员的顺式作用元件的种类和数量较多,说明SHMT基因家族可能具有丰富的潜在功能.

图4 牡丹SHMT基因启动子的顺式作用元件图
2.5 SHMT基因的系统发育分析
利用牡丹和拟南芥的SHMT基因构建系统发育树如图5所示,结果显示牡丹SHMT基因分为三个亚类.其中Ⅰ亚类含有11个牡丹SHMT基因和拟南芥SHMT3基因,Ⅱ亚类含有20个牡丹SHMT基因和2个拟南芥SHMT (SHM1、SHM2)基因,Ⅲ亚类仅包含2个牡丹SHMT基因和4个拟南芥SHMT基因(SHM4、SHM5、SHM6和SHM7).
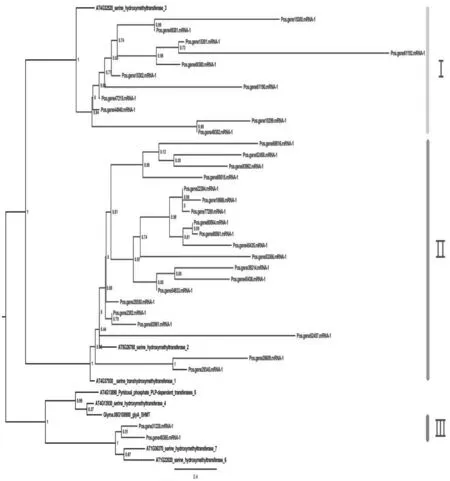
图5 牡丹SHMT基因系统发育树
3 讨论与结论
3.1 不同物种间SHMT基因数量的差异
随着基因克隆技术的进步和众多植物基因组测序的完成,很多植物的SHMT基因被鉴定,并对它们的结构和功能及表达调控进行了研究[14].例如:拟南芥有7个SHMT基因,毛果杨(Populustrichocarpa)有9个SHMT基因[15],水稻(Oryzasativa)有5个SHMT基因[16],坛紫菜(Pyropiahaitanensis)有2个SHMT基因[17],大豆有14个SHMT基因[18],而本研究在牡丹基因组中鉴定出33个SHMT基因.由此可见,不同物种间SHMT基因的数量具有差异性.
3.2 不同物种基因组大小与SHMT基因数量的关系
坛紫菜基因组大小为50.25 Mb,含有2条SHMT基因;拟南芥基因组大小为125 Mb,含有7条SHMT基因;水稻基因组大小为307.2 Mb,含有5条SHMT基因;毛果杨的基因组大小为392.2 Mb,含有9条SHMT基因;大豆基因组大小为978.4 Mb,含有14条SHMT基因;牡丹的基因组大小为11.5 G,有33条SHMT基因,由此可见,SHMT基因数量和基因组的大小呈一定的正相关关系.
3.3 牡丹SHMT基因功能的预测
启动子顺式作用元件预测表明,在牡丹SHMT基因启动子中存在大量参与植物生长发育、应激反应的顺式元件,表明其潜在的功能多样.在该家族中的诸多作用元件中,光反应元件占比最多,除Pos.gene49382基因以外,其余32个成员均具有一个或多个光反应作用元件,可以推测该家族成员与光反应有着密切的关系.值得注意的是,牡丹SHMT基因家族中有3个成员含有与黄酮类物质生物合成相关的调控元件,而黄酮类化合物又有着抗肿瘤[19]、降血糖[20]、抗氧化性[21]等功能.可以推测这些基因在黄酮类生物合成方面发挥着重要作用,可能使牡丹具有以上功能.
目前已有许多研究表明SHMT蛋白在植物抗病中发挥着重要功能.例如,拟南芥AtSHM1基因与大豆SHMT基因发生突变后,植株的抗病虫害功能均发生变化.本研究构建的系统发育树结果显示Ⅱ亚类的牡丹SHMT基因数量较多,且与拟南芥SHM1基因的亲缘性较近,可以为推测该亚类中的牡丹SHMT基因的抗病虫害功能提供一定的理论支撑.
猜你喜欢
新医学(2023年10期)2023-12-09
现代临床医学(2023年1期)2023-03-24
中学生天地(A版)(2023年1期)2023-02-17
中国医药科学(2022年5期)2022-05-05
南方医科大学学报(2022年3期)2022-04-13
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
山东农业工程学院学报(2016年6期)2016-12-01
中国卫生标准管理(2015年3期)2016-01-14
