
长白山红松和臭冷杉树轮密度变化特征及其气候响应
2023-11-24 17:05如先古丽阿不都热合曼张同文喻树龙袁玉江张瑞波王兆鹏王勇辉
地球环境学报 2023年5期
如先古丽·阿不都热合曼 ,张同文,喻树龙,袁玉江,张瑞波,王兆鹏,郭 冬,王勇辉
1.新疆师范大学 地理科学与旅游学院,干旱区湖泊环境与资源重点实验室,乌鲁木齐 830054
2.中国气象局树木年轮理化研究重点实验室,新疆树木年轮生态实验室,乌鲁木齐 830002
3.哈尔滨师范大学 地理科学学院,哈尔滨 150025
IPCC第六次评估报告指出,较工业化(1850 —1900 年)前水平,2010 — 2019 年人类活动引起的全球平均温度升高约1.07℃(0.8 — 1.3℃)(樊星等,2021;IPCC,2021)。气候变暖的加剧对全球降雨的分布也产生了影响,其中季风降水发生着复杂的变化(周天军,2021-08-16)。在气候变暖的大背景下,了解过去气候变化对于认识气候变化规律和预测未来气候变化趋势,从而对人类更好地应对由全球变化引起的各种问题具有重要的意义。
记录过去气候变化的信息载体较多,其中,树木年轮因其具有易获取、定年准确、分辨率高等优势成为国际上获取过去气候变化的重要代用资料之一(Yuan et al.,2007;Cook et al.,2015)。树木年轮密度分析使树木年轮学的研究有了重大的突破(魏本勇和方修琦,2008)。有研究表明:冷湿地区的针叶树种树轮密度指标较宽度指标可能表达出更为丰富的气候信息,有助于研究树木生长对气候的响应以及过去气候的重建(Hughes et al.,1984;Cook and Kairiukstis,1990)。国内树轮密度研究相对较晚(魏本勇和方修琦,2008;张同文等,2011a)。20 世纪90 年代,吴祥定和邵雪梅(1994)首次在秦岭进行树轮密度研究,分析结果表明密度变化较宽度对气候的响应可能更加显著,之后相关研究逐渐开展。王丽丽等(2005)发现漠河的落叶松(Larix gmelinii)与樟子松(Pinus sylnestrisvar.mongolica)的树轮最大密度均与生长季后期的温度显著相关。在横断山脉中部,麦吊云杉(Picea brachytyla)的最大晚材密度主要受夏季温度的影响(Fan et al.,2009)。袁玉江等(2008)对天山西部上树线3个采样点云杉(Picea asperata)的树轮最大密度研究表明,在年降水量越多的山区,云杉树轮最大密度年表与温度的相关性越好。我国树轮密度研究大多集中在干旱半干旱地区,在季风区的相关研究则较为有限。在位于亚热带季风区的福建省开展树轮密度研究发现,相对湿度是影响当地杉木(Cunninghamia lanceolata)树轮密度变化的主导因素(张辉等,2021)。
长白山位于我国东北季风区,是气候敏感区和生态环境脆弱地带(王晓东,2011),也是我国东北地区保存最为完整、面积最大的温带原始森林分布区。该区域拥有广阔的原始森林和丰富的物种,为树轮学研究提供了大量的资源,因此受到树轮研究者的广泛关注(韩艳刚等,2019)。前人在长白山的树轮学研究已证明,同一生境、不同树种生长受气候变化的影响存在差异(王晓春,2004),对气候因子的响应关系也呈现不稳定(高琳琳等,2011;高露双等,2011;王守乐等,2019)。苏金娟和王晓春(2017)指出张广岭同一地区不同阔叶树种的生长具有物种特异性。也有研究人员发现,长白山过渡带红松(Pinus koraiensis)的径向生长受生长季温度和降水共同限制,而鱼鳞云杉(Picea jezoensis)径向生长受生长季初期降水的影响更大,全球变暖可能有利于高海拔红松生长,而鱼鳞云杉对气候变化不敏感(高露双等,2011)。目前,对长白山的树轮气候学研究成果颇丰,然而大量研究均基于树轮宽度,气候变化对同一生境、不同树种的树轮密度变化的影响还有待揭示。
本文采用树轮密度分析方法,以长白山建群树种红松(Pinus koraiensis)和臭冷杉(Abies nephrolepis)为研究对象,分析不同树种树轮密度参数与逐月、不同生长期气候因子相关关系的差异性并检测相关关系的稳定性。本研究有助于进一步丰富长白山地区同一生境、不同树种的树轮密度变化特征及其气候响应的研究,并为今后这一地区的气候重建提供基础信息。
1 材料与方法
1.1 研究区概况
采样区位于我国长白山东北部汪清金沟岭林场低海拔区域的针阔混交林带(43°21′ N、130°10′ E,海拔为634 — 696 m),地形属山地丘陵地带。该区属于中温带季风气候,具有夏季多雨、冬季寒冷的特点。全年均温为3.9℃,年降水量为600 — 700 mm(方伟东等,2011)。研究区域具有明显的植被垂直带,其中林场的顶级群落主要为臭冷杉针叶林及针阔混交红松林,并且主要分布在海拔400 m 以上地带。土壤类型为山地暗棕色森林土(张悦等,2015;韩金等,2019)。
红松在长白山林区主要分布在海拔500 —1200 m,属半阳性、浅根性树种,主根不发达,耐寒性强却不耐湿、不耐干旱,对土壤水分要求较高(康雨昌等,2019)。臭冷杉在长白山主要分布在海拔500 — 1800 m,属阴性、耐荫性树种,生长在冷湿环境及酸性土壤,适应性强(郑万钧,1983)。红松和臭冷杉作为长白山的建群树种,对气候变化敏感,在维持森林生态系统的稳定性方面具有重要作用(朱良军等,2015;覃鑫浩等,2016)。
1.2 样本采集与树轮年表建立
两个树种的树芯样本采集于2016 年5 月,且采样区域无明显人类活动影响。因当年生长季尚未结束,所以所用的树轮密度数据截至2015 年。采样树种分别为红松(JGL1,43°21′ N、130°10′ E,海拔为696 m)和臭冷杉(JGL2,43°20′ N、130°10′ E,海拔为634 m)。由于两个采样点空间位置接近且海拔差异不大,所以将其视为同一生长环境。
树轮密度分析之前,先依据树木年代学基本原理进行树轮宽度研究工作。随后对已测宽样本进行筛选,再对其脱糖脱脂,并且根据样本木纤维走向的角度将样本切成梯形断块固定在板上,再利用DendroCut 仪器将样芯切成1 mm 的薄片,最后用X 射线透射成像制片(Holmes,1983)。利用Dendro-2003 树轮密度分析系统将光学强度转化为树轮密度数据,获得全轮密度(tree-ring wood density,TRD)、最大密度(max wood density,MXD)、最小密度(min wood density,MND)、早材平均密度(earlywood density,EWD)和晚材平均密度(latewood density,LWD)5 种树轮密度参数(王丽丽等,2005)。结合已完成的树轮宽度定年结果,通过树轮密度分析仪和COFECHA程序对树轮密度参数进行交叉定年和质量控制。最终利用ARSTAN 程序的负指数函数去趋势方法,完成两树种年表质量较好的全轮、早材和晚材密度的标准化年表(standard chronology,STD)(Fritts,1976;Cook,1985)。
1.3 气象资料
距离研究区较近的气象站点有两个:延吉气象站(42°53′ N、129°28′ E,海拔179 m,资料长度:1953 — 2015 年)和汪清气象站(43°20′ N、129°46′ E,海拔461 m,资料长度:1989 — 2015年)。其中,汪清气象站从位置和海拔上更接近采样点,但其记载数据长度较短。相关分析结果显示:两个气象站月均气温和降水量数据呈极显著相关关系(p<0.01)。其中,两气象站间月均气温的相关系数为0.999,月降水量的相关系数为0.997。因此,选择了记载数据较长的延吉气象站。本研究所选气候要素有平均气温(T)、平均最高气温(Tmax)、平均最低气温(Tmin)和降水量(P),时段为1953 — 2015 年。图1 为研究区域多年平均气候状况。该区域雨热同期,呈单峰型分布,秋冬季低温少雨,夏季高温多雨。1 月平均气温可达-13.7℃,降水量仅4.5 mm,8 月平均气温为21.4℃,降水量117.9 mm。通过对延吉气象站1953 — 2015 年年均气温、年均最高气温、年均最低气温和年降水量的变化趋势分析发现,年均气温和年均最低气温呈显著上升趋势(p<0.001),年均最高气温和年降水量变化趋势则不显著。
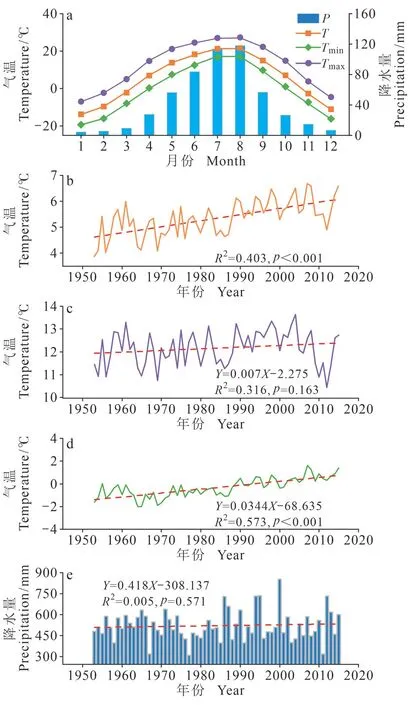
图1 延吉气象站(1953 — 2015 年)月均温、月均最高气温、月均最低气温、月降水变化特征(a);年均气温(b)、年均最高气温(c)、年均最低气温(d)、年均降水量(e)变化Fig. 1 Monthly and annual climatic data for the Yanji during 1953 — 2015; monthly average temperature, average maximum temperature, average minimum temperature, and precipitation (a);trend of annual average temperature (b), annual average maximum temperature (c), annual average minimum temperature (d), annual precipitation (e) change
1.4 数据分析
由于树木年轮对气候因子的响应具有“滞后效应”(Fritts,1976;吴祥定,1990),故选取上一年9 月至当年10 月的平均气温、平均最高气温、平均最低气温和降水量与两个树种的全轮、早材和晚材密度的标准化年表做逐月相关分析。树木年轮的形成与变化不仅受单月气候要素的影响,还受多个月份气候因子的综合影响(孙毓等,2012)。因此,结合长白山气候状况和研究树种的树木生理学意义,将红松和臭冷杉的生长季节划分为上年生长季晚期PEG(P9 — P10)、冬季WI(P11 — P12,C1 — C3)、当年生长季早期CB(C4 — C5)、当年生长季中期CG(C6 — C8)、当年生长季晚期CEG(C9 — C10),用以研究树轮密度在不同生长期对气候因子的响应(鲍安,2019)。此外,为进一步了解不同树种与气候因子间的相互关系,采用响应面函数进行分析。响应面函数以树轮年表为因变量,气候因子为自变量,通过描述年表与气候因子间的线性关系,气候因子的幂次和因子间的乘积关系,更能体现出年表与气候因子间的相互联系(张志华等,1996;喻树龙等,2012)。其数学模型表达式为:
式中:Y表示树轮指数;T、P分别代表与树轮指数相关的某一时段的气候因子指标;a、b分别代表随机误差和气候变量。
最后,为了阐明不同树种树轮密度变化与气候因子动态关系的差异,以30 a 为滑动窗口,1 a为滑动步长,进行滑动相关分析。
2 结果与分析
2.1 年表的基本统计量
由表1 可知,在两树种各密度年表的公共区间(1949 — 2015 年),红松和臭冷杉的全轮密度年表的树间平均相关系数、信噪比和样本代表性大于其各自的其他密度年表。结果表明:该采样点的全轮密度年表中包含的气候变化信息可能较多,具有区域代表性。平均敏感度表示年轮时间序列对气候信息的敏感程度(吴普等,2006;杨银科等,2006),其中红松和臭冷杉密度年表均符合树木年轮密度研究标准(包光等,2015),红松树轮密度年表的敏感度较臭冷杉大。红松的全轮和早材密度年表的一阶自相关大于臭冷杉年表,说明上年气候特征变化对当年红松的树轮密度影响比臭冷杉更大(张同文等,2015)。结合表1 的变异系数值和两树种年轮密度标准年表的序列变化趋势图(图2),可知在公共区间,红松和臭冷杉的树轮密度年表变化趋势具有一定的差异性,臭冷杉密度变化较红松稳定。
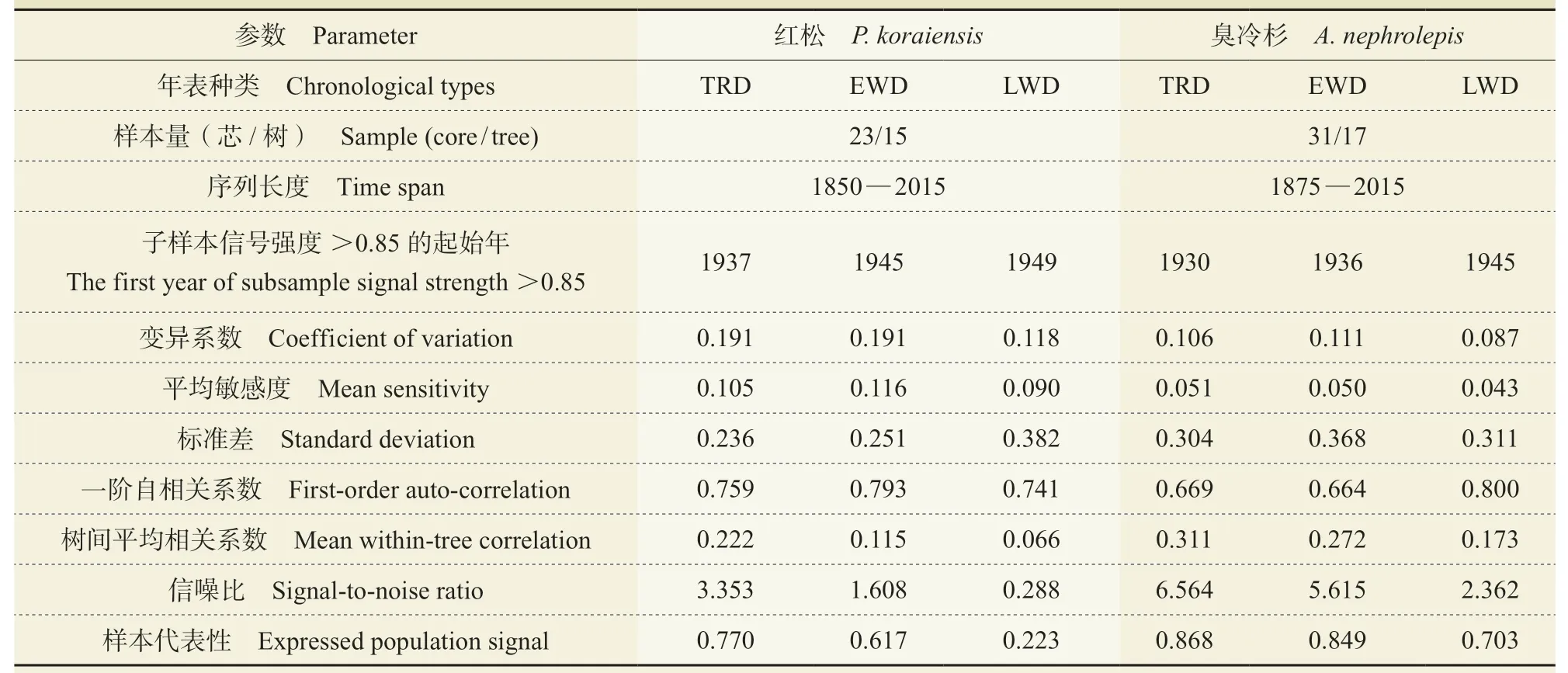
表1 长白山红松和臭冷杉树轮密度标准化年表公共区间(1949 — 2015 年)特征参数及分析Tab. 1 Statistical characteristics of density chronologies and the 1949 — 2015 common period analysis
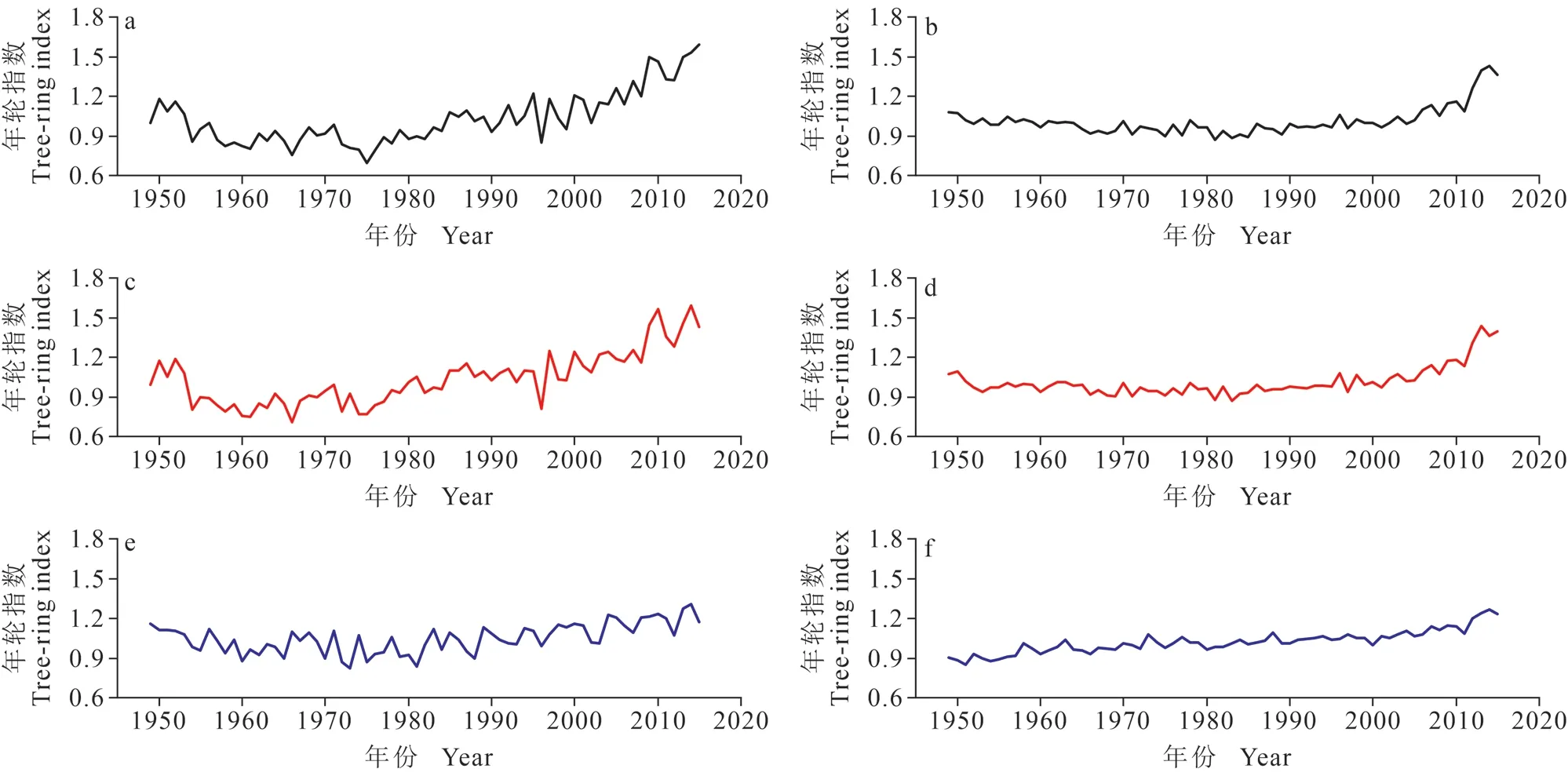
图2 长白山红松和臭冷杉标准化密度年表(公共区间:1949 — 2015 年);红松的全轮密度年表(a);臭冷杉的全轮密度年表(b);红松的早材密度年表(c);臭冷杉的早材密度年表(d);红松的晚材密度年表(e);臭冷杉的晚材密度(f)Fig. 2 Tree-ring wood density chronologies for the 1949 — 2015 common period, chronology of P. koraiensis tree ring annual wood density (a); chronology of A. nephrolepis tree ring annual wood density (b); chronology of P. koraiensis earlywood density (c);chronology of A. nephrolepis earlywood density (d); chronology of P. koraiensis latewood density (e); chronology of A. nephrolepis latewood density (f )
为了描述红松和臭冷杉密度和宽度标准化年表相关关系,计算各树轮密度、宽度年表间的互相关系数。由表2 可知:红松的密度变化与其径向生长呈负相关,而臭冷杉的径向生长与全轮、早材密度年表呈负相关,与晚材密度的相关性不显著。说明臭冷杉早材密度对整轮宽度的影响较大。两树种各树轮密度标准化年表间互相关关系显著,均通过了99%的显著性检验(n= 67,p<0.01),全轮密度年表均与其本身的早材密度年表相关系数最高,表明早材密度在全轮密度中所占比重较大,可能对气候因子的响应存在相似性。红松和臭冷杉的树轮密度和宽度年表间呈极显著正相关关系,表明密度和宽度年表之间存在较好的相关性,其树轮变化可能受同一种气候因素影响。
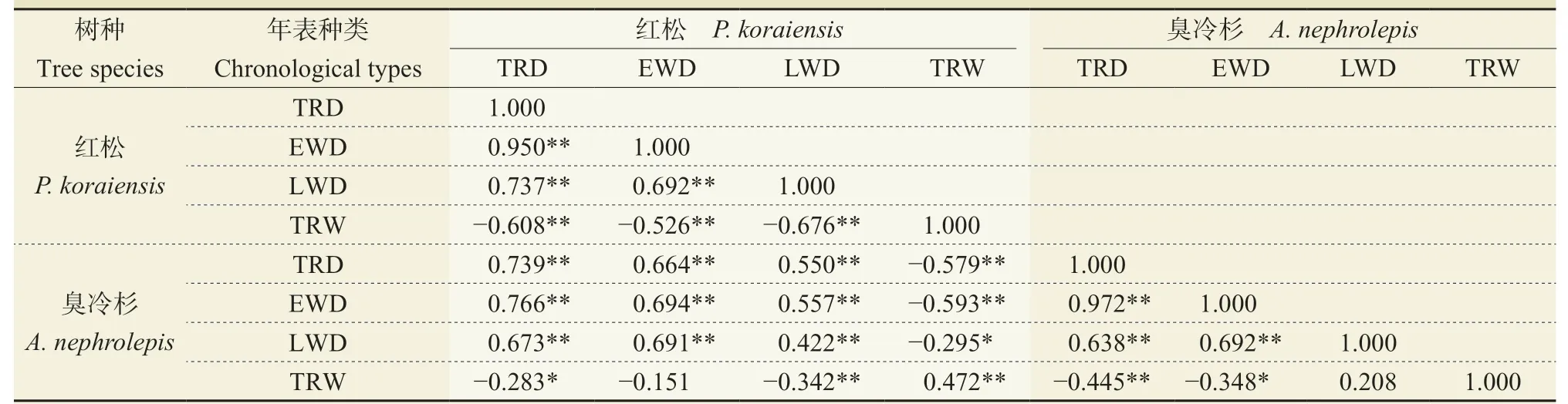
表2 长白山红松和臭冷杉树轮密度、宽度标准化年表相关系数(公共区间:1949 — 2015 年)Tab. 2 Correlation coefficients of tree ring density and width chronologies for the 1949 — 2015 common period
2.2 树轮密度与气候因子相关关系
红松和臭冷杉年轮密度标准年表与气候因子的响应分析(图3、图4)结果表明:整体而言,两树种树轮密度变化与平均气温、平均最低气温呈正相关关系,而对平均最高气温和降水量的响应较弱。其与平均最低气温在全生长期呈显著正相关,尤其与生长季早期(4 — 5 月)的平均最低气温相关性最大。红松和臭冷杉的早材密度年表受平均最低气温影响的月份较其他密度年表更广泛,均与上年10 — 11 月、当年4 — 6 月、当年10 月的平均最低气温呈极显著正相关。晚材密度受平均气温影响的月份较其他密度年表更广泛,均与当年生长季晚期的平均气温显著相关。两树种各树轮密度年表与气候因子相关性的差异具体表现为:红松树轮密度变化对气候因子的响应较臭冷杉更敏感,其各密度年表与气候因子相关性显著的月份多于臭冷杉。同时,红松的树轮密度受上年气温限制作用的月份比臭冷杉更广泛。红松的早材密度年表与上年9 月的平均气温和上年10 — 11 月的平均最低气温呈极显著正相关,晚材密度年表与上年8 月、9 月的平均气温,上年8 月的平均最高气温,上年9 月、11 月的平均最低气温呈极显著正相关,而臭冷杉全轮和早材密度与上年10 月的平均气温和上年10 月、11 月的平均最低气温呈显著相关。
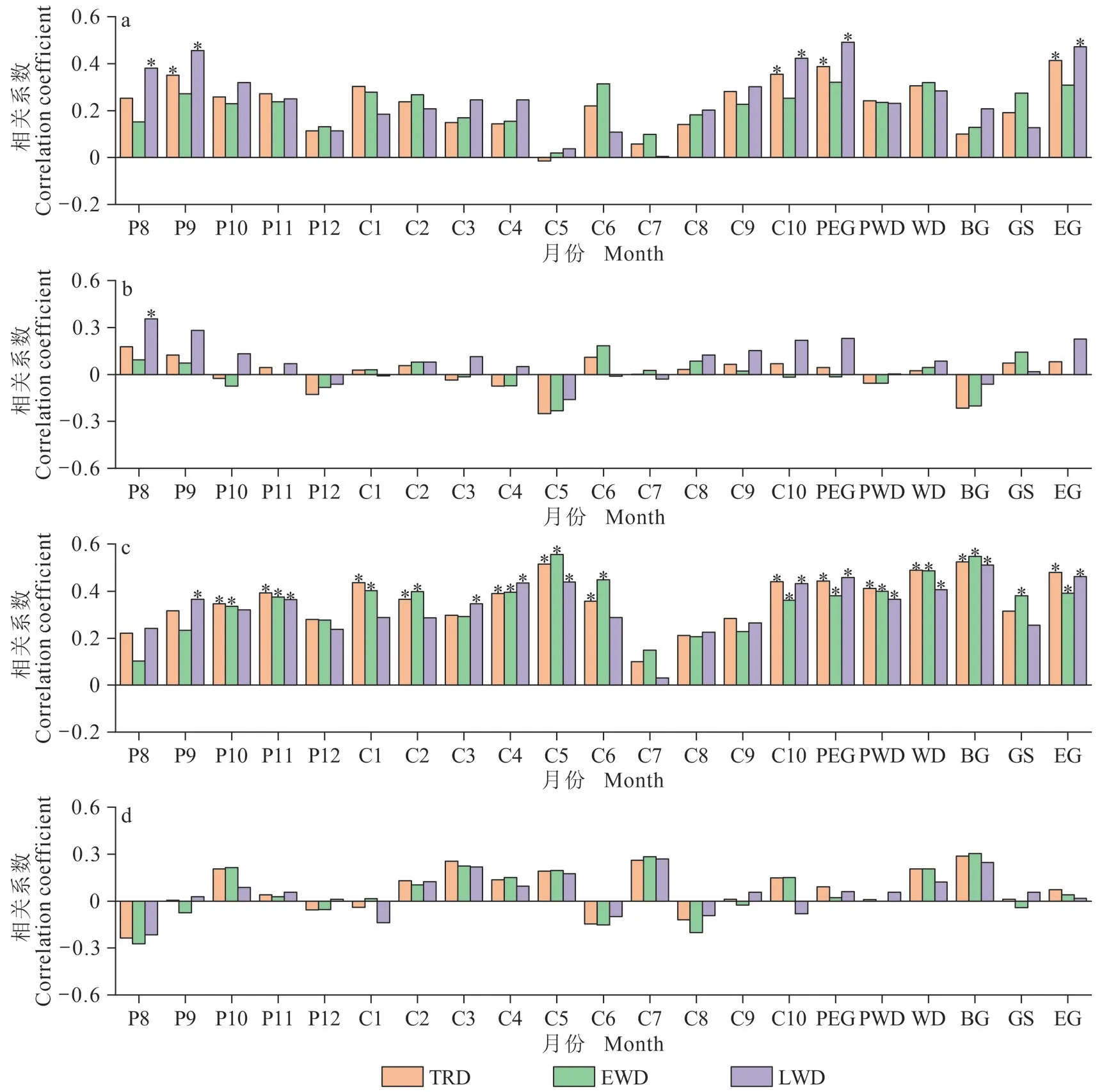
图3 长白山红松的标准化密度年表与气候因子的相关分析;红松与平均气温的相关分析(a);红松与平均最高气温的相关分析(b);红松与平均最低气温的相关分析(c);红松与降水量的相关分析(d)Fig. 3 Correlation analysis of P. koraiensis standardized chronologies with climate factors, average temperature (a);average maximum temperature (b); average minimum temperature (c); precipitation (d)
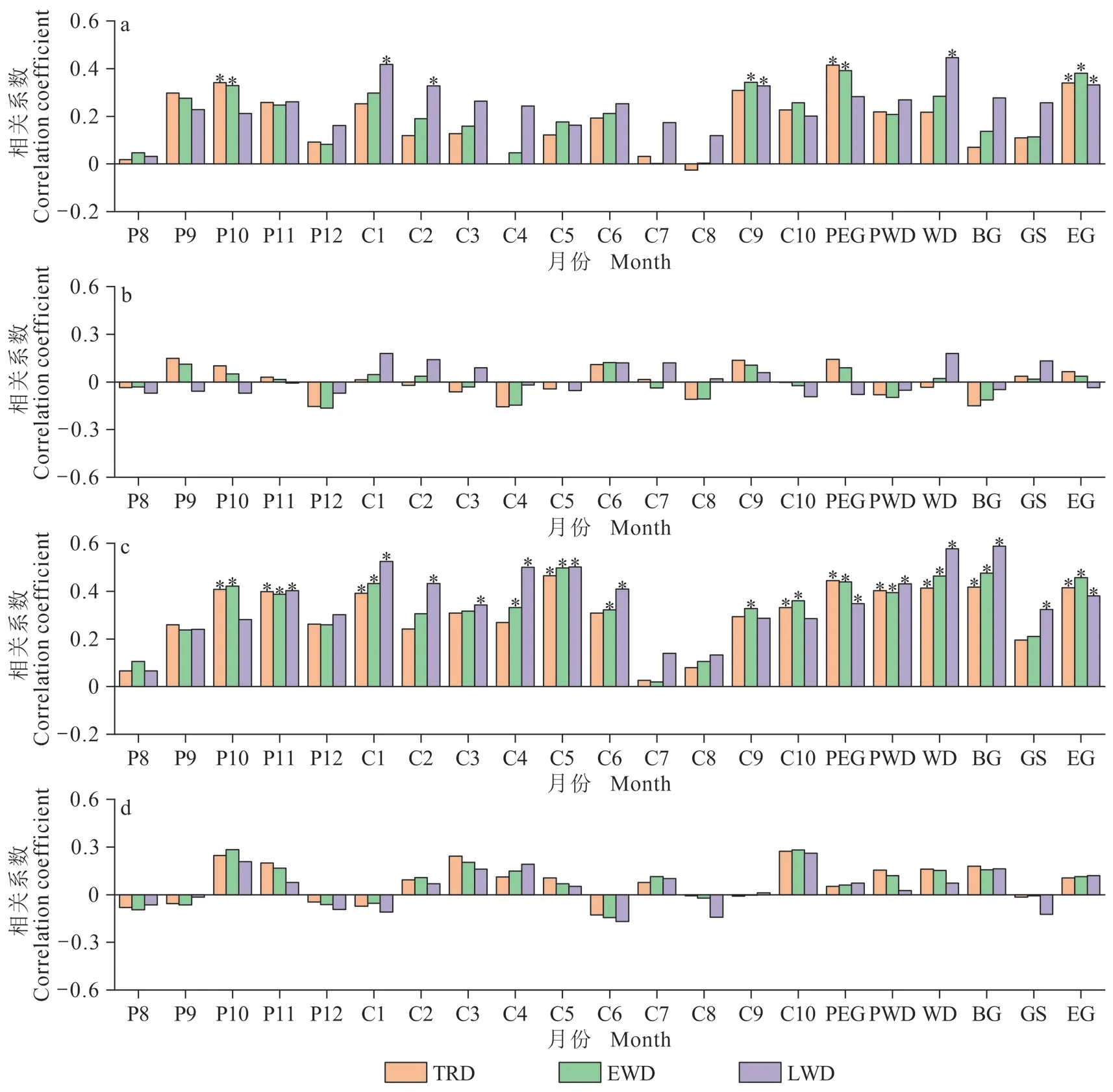
图4 长白山臭冷杉的标准化密度年表与气候因子的相关分析;臭冷杉与平均气温的相关分析(a);臭冷杉与平均最高气温的相关分析(b);臭冷杉与平均最低气温的相关分析(c);臭冷杉与降水量的相关分析(d)Fig. 4 Correlation analysis of A. nephrolepis standardized chronologies with climate factors; average temperature (a);average maximum temperature (b); average minimum temperature (c); precipitation (d)
2.3 树轮密度与气候因子的响应面分析
由图3、图4 可知:红松和臭冷杉早材密度均与生长季早期(4 — 5 月)的平均最低气温相关系数最大。为了进一步了解生长季早期最低气温和降水对两树种早材密度变化的影响,以早材密度指数为因变量,生长季早期平均最低气温和降水量为自变量,通过响应面分析客观地表达出两树种早材密度年表与生长季早期气候因子间的相互作用。
由图5 可知:红松和臭冷杉树轮早材密度对生长季早期的平均最低气温和降水量响应大致相同。最低气温对早材的影响更大。在气温较高时,随着降水量的增加,早材树轮密度值呈现先增大后减小的趋势,且降水量越多,这种趋势越明显。当降水量较多时,随着气温的上升,早材密度呈现先减小后增大的趋势。臭冷杉树轮密度只有在较高气温和较高降水量时以上趋势显著,表明红松树轮密度对气候因子更敏感。

图5 长白山红松(a)和臭冷杉(b)的标准化早材密度年表与气候因子的响应面分析Fig. 5 Response surface of P. koraiensis (a) and A. nephrolepis (b) standard chronologies with climatic factors
2.4 树轮密度与气候因子的稳定性关系
根据图3、图4,树轮密度变化与单月气候因子和季节性气候因子响应结果大致相同。进一步研究红松和臭冷杉树轮密度变化与单月气候因子的长期动态变化过程,滑动相关分析结果显示:红松和臭冷杉在1953 — 2015 年,树轮密度年表对水热因子的敏感性有所变化,在部分时段出现相关系数正负变换的情况。
由图6 可知:整体而言,随着气候变暖,两树种树轮密度年表与平均气温、平均最低气温的正响应增强,而对平均最高气温的负响应增强,对降水的响应较弱且相对稳定。红松全轮和晚材密度年表对上年8 月、9 月、当年9 月、10 月(生长季晚期)平均气温和上年9 月、当年4 — 7 月、当年10 月的平均最低气温呈显著正相关,早材密度年表与上年9 月和当年6 月平均气温的正响应增强。而晚材密度年表与上年8 — 10 月平均最高气温的正相关性显著减弱。臭冷杉树轮密度年表对上年10 月和当年9 月平均气温(生长季晚期)和上年10 — 11 月、当年4 — 5 月、当年9 — 10 月最低气温的正响应显著增强。臭冷杉全轮密度年表与上年9 — 10 月、当年6 月平均最高气温的正响应显著减弱。臭冷杉全轮和早材密度与上年12月平均最高气温和当年4 月平均最低气温的相关性分别出现显著的正转负和负转正的情况。
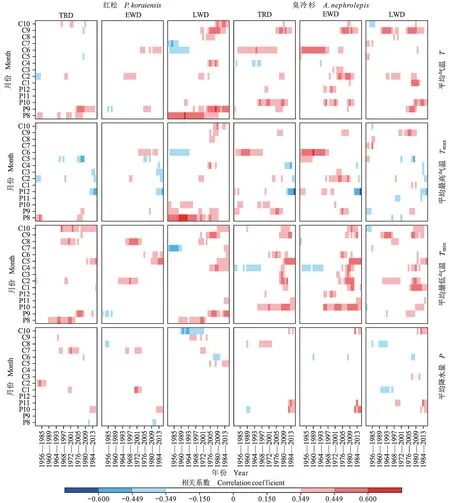
图6 长白山红松和臭冷杉的标准化密度年表与气候因子的滑动相关分析Fig. 6 Moving correlation analysis of standard chronologies with climatic factors
3 讨论
3.1 树木年轮密度对气候因子的响应
红松和臭冷杉的树轮密度和宽度年表呈显著负相关(表2),表明树木径向生长越快,两树种的树轮密度值越大。这与其他研究结果相似(Rathgeber,2017;张辉等,2021),树轮密度间接反映了形成层细胞壁沉积物质的积累和细胞体积的大小,细胞体积越大,细胞壁物质积累越少,树轮密度值越小,反之亦然(徐金梅等,2011)。将同一区域的树木年轮密度年表与月份和季节气候要素进行相关性分析发现(图3、图4),不同树种对同一气候要素的响应存在异同。红松和臭冷杉的树轮密度变化主要受气温的影响,与最低平均气温的正响应更加显著,而与降水量的相关相对较弱。可能是由于研究区在我国气候区划中属于中温带湿润区(李明等,2016),降水充沛,使得树木生长对降水量的敏感性较低。红松和臭冷杉早材密度受平均最低气温的影响较其他年表更广泛,均与上年10 —11月、当年4 — 6月、当年10月及全季(除了臭冷杉早材密度与当年生长季最低气温不显著)的平均最低气温呈极显著正相关。其中,与生长季早期(4 — 5 月)平均最低气温的相关系数最高,表明树轮密度能够很好地指示生长季早期平均最低气温的变化。研究发现(Antonova and Stasova,1993;Hosoo et al.,2002),最低气温一般出现在夜间,同时夜间温度是影响细胞壁物质积累和木质素化的主要因素,进而会影响树轮密度值的大小,因此树木光合作用和呼吸作用强度对树轮密度的变化十分重要。夜间升温促进叶片呼吸和根系呼吸,刺激细胞内碳水化合物等营养物质的消耗(勾晓华等,2007;朱军涛和郑家禾,2022),导致累积的营养物质减少,此时形成的早材细胞管胞扩大较慢,细胞壁厚,细胞充实度高,易形成较大的树轮密度(刘可祥等,2021)。其次,树木的早材在生长季早期生长(Atkin et al.,2007),此时气温回升,树木体内代谢加快,其与大气、土壤间的物质和能量交换加快,产生树干液流(白志强等,2016)。生长季早期夜间气温偏高加大了土壤蒸发量和树木的蒸腾作用,从而使树木液流减少,使得白天积累的光合产物逐渐被消耗(张同文等,2011b),不利于树木的早材细胞生长,增大了早材密度值。红松和臭冷杉的晚材密度年表对平均气温的敏感性强于其他密度年表,均与当年生长季晚期的平均气温显著相关。晚材的生长期主要在生长季晚期(9 —10 月),主要表现为细胞壁物质的积累。生长季晚期的高温,不利于研究区红松和臭冷杉的光合作用,细胞分裂速率减小,而细胞径向生长时间缩短,延长了细胞增厚时间,产生了窄而厚的晚材细胞(Cuny et al.,2014)。
响应面分析进一步明确了生长季早期平均最低气温对早材密度值的影响。气温较高时,随着降水量的增多,早材密度值先增大后减小。当降水量较多时,随着气温的上升,早材密度呈现先减小后增大的趋势。当平均最低气温较高,降水量偏少时,树木的呼吸作用和蒸腾作用增强,影响细胞的径向生长,从而增大了早材密度值,而当降水量达到一定阈值时,水分比较充足,最低温度高,生长期延长,树木的营养物质积累增加,利于树木径向生长,密度值反而减小(喻树龙等,2008)。低温和多雨的气候条件限制新芽、叶片和根系的形成,从而影响到水分和无机物的吸收及光合效率,细胞分裂速度减小,形成的细胞导管窄,形成较大的早材密度(吴祥定,1990)。
分析红松和臭冷杉树轮密度变化对气候因子的响应发现,红松树轮密度变化对气候因子的响应更敏感,各密度年表与气候因子相关性显著的月份多于臭冷杉,且响应面分析结果也表明其响应更敏感。红松的树轮密度受上年气温限制作用的月份比臭冷杉更广泛。树轮密度年表特征参数的一阶自相关结果(表2)也表明其受上年气候因子响应的“滞后效应”明显强于臭冷杉。红松的早材密度年表与上年8 — 11 月的气候因子呈正相关,而臭冷杉全轮和早材密度与上年10 月的平均气温和上年10 月、11 月的平均最低气温呈显著相关。红松喜光性强,属半阳性树种,光饱和点较高(郑万钧,1983;朱良军等,2015),耐旱性比臭冷杉强,球果第二年9 — 10 月成熟;而臭冷杉为耐荫性树种,喜冷湿的环境(张继澍,1999;覃鑫浩等,2016),光合能力比红松弱。同时,不同树种的叶片结构不同,如叶片表面的绒毛、蜡质以及叶肉细胞的排列等都会影响其叶绿素浓度和光合作用速率。刘伟国(2010)研究长白山主要树种光谱反射特征,将红松归为叶片叶绿素含量较低,但光合作用速率相对较高的树种。因此,升温导致的生长季延长和有效积温的增加(张辉等,2021),可能是好光且喜温、光合作用速率相对较高的红松树轮密度变化对气候因子的响应更敏感的原因(朱良军等,2015)。说明树种特性可能是造成两树种树轮密度对气候响应差异的关键因素。
3.2 树木年轮密度与气候因子关系的稳定性
通过分析红松和臭冷杉与气候关系随时间变化的稳定性,以探讨气候变暖对长白山两树种树轮密度变化的影响。结果显示(图6):红松和臭冷杉树轮密度变化整体上呈现对平均气温和平均最低气温的正响应增强,对平均最高气温的响应减弱,对降水的响应相对稳定。可能是由于近年来,我国东北地区的昼夜温差(DTR)降幅大,平均最低气温上升速率明显高于平均最高气温,使当地出现日夜增暖的不对称现象(勾晓华等,2007;Gao et al.,2022)。昼夜的不对称变暖影响了植被的生长期、生长季长短,同时对气候因子关系的稳定性产生了一定的影响(Rehmani et al.,2014)。红松和臭冷杉的树轮密度均对9 月、10 月(上年和当年生长季晚期)的平均气温,4 — 5 月(生长季早期)、生长季晚期的平均最低气温的正响应显著增强,与生长季晚期最高气温的正响应显著降低。生长季早期和晚期较高的温度和低降水加强了植物的呼吸速率和蒸腾作用,造成土壤水分亏缺并提高蒸气压差,减少了养分的运输,不利于树木径向生长,树轮密度值增大(于健等,2021)。但随着气候变暖,最高气温(日间温度)上升,导致植物通过叶片光合作用的碳吸收补偿夜间温度升高引起的呼吸碳释放,促使植物积累更多的碳(朱军涛和郑家禾,2022),增强了植物的光合作用,用于树木径向生长(勾晓华等,2007),从而降低了密度值,树轮密度与最高气温的正相关性便开始减弱。这也可能是适宜的温度条件下,树木本身趋于适应外界环境(张辉等,2020)。在气候变暖下,臭冷杉树轮密度变化与气候因子的关系不如红松稳定。其早材密度年表与上年12 月平均最高气温、当年4 月平均最低气温的相关性分别出现了显著的正转负和负转正的情况。气候的持续变暖,尤其是最高最低温的非对称变化可能改变了喜冷湿树种的生长环境,使臭冷杉的树轮密度变化出现了响应“分离现象”。以往的研究表明,北半球高纬度地区森林常常会发生响应“分异现象”(D’Arrigo et al.,2008)。这些研究多针对树木径向生长 — 气候响应,而树轮密度对气候因子响应的稳定性研究较少,为此后续还将进一步加强研究。
综上所述,长白山红松和臭冷杉的树轮密度数据能够较好地指示生长季早期的最低气温变化,且红松树轮密度包含更多的气候信息,其树轮密度信息用来重建研究区过去气候变化有较大的潜力。
4 结论
本文研究了长白山红松和臭冷杉树轮密度变化特征及其对气候因子的响应。两树种各树轮密度年表的特征参数及年表变化趋势表明,红松包含更多的气候信号,而臭冷杉树轮密度变化较稳定。红松和臭冷杉的密度变化主要受气温的影响。生长季早期最低气温对早材密度变化影响最显著,表明生长季早期最低气温可能具有重建研究区过去气候变化的潜力。相对臭冷杉而言,红松树轮密度对气候变化更敏感,且受上年气候因子响应的“滞后效应”明显强于臭冷杉,表明红松树轮密度可能具有记录研究区过去气候信息的巨大潜力。两树种不同树轮密度年表对气候因子的响应存在差异,晚材密度对平均气温敏感,而早材密度年表与平均最低气温的相关性较高。随着气候变暖,红松和臭冷杉树轮密度年表对气候因子响应的动态稳定性出现对生长季晚期平均气温和生长季早晚期平均最低气温的正响应增强,对生长季晚期平均最高气温的正响应减弱的趋势,而对降水的响应稳定性较高。臭冷杉早材密度年表与上年12 月平均最高气温、当年4 月平均最低气温的相关性分别出现了显著的正转负和负转正的情况。
猜你喜欢
世界建筑(2022年10期)2022-10-25
山西林业科技(2022年4期)2022-03-12
广西农学报(2021年2期)2021-11-21
生态学报(2021年14期)2021-09-01
景德镇陶瓷(2021年1期)2021-03-24
艺术品鉴(2019年12期)2020-01-18
花火A(2019年1期)2019-03-22
现代养生·上半月(2017年7期)2017-07-07
环球人文地理·评论版(2016年6期)2016-08-23
沙漠与绿洲气象(2014年3期)2014-03-25
