
MC1R 基因多态性与杜黑猪毛色关系研究
2023-11-23 03:26:40巩静杨晴王艺盼王彦平朱晓东张传生耿立英王继英
山东农业科学 2023年10期
巩静,杨晴,王艺盼,王彦平,朱晓东,张传生,耿立英,王继英
(1. 河北科技师范学院动物科技学院,河北 秦皇岛 066600;2. 山东省农业科学院畜牧兽医研究所/山东省畜禽疫病防治与繁育重点实验室/农业农村部畜禽生物组学重点实验室,山东 济南 250100;3. 枣庄黑盖猪养殖有限公司,山东 枣庄 277100)
毛色是猪的重要品种特征之一[1],一直以来备受人们关注。 在养猪生产中,毛色被广泛应用于评判品种的纯度、遗传稳定性、亲缘关系,以及在配套系培育中确定亲本的杂交组合类型等方面。 揭示猪毛色的遗传机制及影响毛色的相关基因的遗传规律,对猪的育种具有重要意义[2-3]。毛色不同的品种或品系杂交时,F1代呈现显性毛色,F2代群体则会出现毛色分离[4],选择毛色一致性强的种猪,淘汰毛色分离严重的个体,提高群体的整齐度,是新品种或配套系培育过程中的重要步骤[5]。
哺乳动物毛色的形成是由一系列与黑色素细胞种类与形成有关的生理、生化反应的最终结果,遗传机制较复杂。 猪的毛色大致分为野生型毛色、纯黑色、纯白色、棕红色、两头乌、花猪等。 虽然属于质量性状,由少数几对基因决定,但由于基因之间有上位效应等遗传互作效应,调控毛色的基因及其作用机制尚未完全搞清楚。 黑色素皮质素受体1(MC1R)基因是调控哺乳动物毛色的一个关键基因[6-8]。MC1R是G 蛋白偶联受体家族的一员,由Extension 基因座编码,通过与其配体促进剂α-黑素细胞刺激激素(α-MSH)或颉颃剂刺鼠蛋白(Agouti)的竞争性结合分别形成真黑素或褐黑素, 从而影响猪毛色表型[9]。 Kijas等[10-11]在欧洲野猪、欧洲大黑猪、中国梅山猪、汉普夏猪、大白猪、皮特兰猪及杜洛克猪的MC1R基因部分序列中发现8 个突变构成5 种等位基因,其中E+对应于野猪的灰毛色,ED1对应于欧洲大黑猪和中国梅山猪的黑毛色,ED2对应于汉普夏猪的黑毛色,Ep对应于皮特兰猪的黑色斑点及大白猪的白毛色,e 对应于杜洛克猪的棕红毛色。Fang 等[12]在31 头欧洲家猪和19 个亚洲品种家猪及15 头亚欧野猪的MC1R基因编码区内共发现14 个突变,其中包括Kijas 等发现的8 个突变。
枣庄黑盖猪是山东省著名地方品种,毛色为纯黑色,具有肉质鲜嫩、产仔数多、抗病能力强、适应性强等优良特征,但其生长缓慢,产肉性能和胴体瘦肉率低[13-14]。 利用杜洛克对枣庄黑盖猪进行杂交改良,瘦肉率、肉质性能及胴体性能均明显提高,为枣庄黑盖猪杂交改良的优良杂交组合之一。 我国多数黑猪毛色黑色对棕红色呈显性遗传,杜洛克猪与枣庄黑盖猪(以下简称杜黑猪)F1杂交后代的毛色全部为全黑色,F2代群体出现了毛色分离。 常规育种对猪毛色的选育是横交固定后,通过持续的世代选育逐渐剔除后代中毛色出现分离的个体,周期长、效率低,严重影响和制约了专门化品系选育和新品种(配套系)育种进程。
本研究以杜黑猪F2杂交群体和枣庄纯种黑盖猪为研究对象,对F2群体进行毛色表型类型分组,利用液相芯片捕获测序技术[15]鉴定MC1R基因中影响毛色的10 个SNP 的基因型,研究不同基因型与毛色表型的关系,筛选可用于毛色分子标记辅助育种的SNP 位点,为快速建立毛色稳定的杜黑猪新品种培育体系奠定基础。
1 材料与方法
1.1 试验动物及毛色表型收集
试验动物包括390 头杜黑猪F2杂交后代和18 头枣庄纯种黑盖猪,均来自枣庄黑盖猪养殖有限公司。 将毛色分为全黑色(枣庄黑盖猪毛色)、棕红色(杜洛克毛色)、黑红条纹、棕色偏黄色、棕色偏白色等类型,对F2代群体打耳号时记录每头试验猪的毛色,按毛色表型类型进行分组,统计每组毛色的个体数。
1.2 基因组提取及SNP 基因型检测
采集试验猪耳组织放入装有75%乙醇的离心管中,-20 ℃冰箱存储。 利用血液/组织/细胞提取试剂盒[天根生化科技(北京)有限公司]提取耳组织基因组DNA。 经Nanodrop(260/280 在1.7~2.1 范围之间,浓度>50 ng/μL)和0.8%琼脂糖凝胶电泳(条带清晰)检测合格后,委托博瑞迪生物技术有限公司利用定制的液相芯片进行基因组SNP 测定。 该液相芯片为山东省农业科学院畜牧兽医研究所猪繁育饲养团队集成了近年来候选基因、功能基因组研究方面公开发表的功能SNP、Del、Insertion 等标记,采用博瑞迪GBTS 技术体系中的GenoBaits 技术[15]设计并优化开发的一款中低密度(6K)猪功能标记液相芯片,其中MC1R基因中影响毛色的10 个SNP 位点信息参考Fang 等[12],详见表1。
1.3 数据处理与分析
利用Microsoft Excel 软件对杜黑猪毛色表型类型进行统计分析。 利用PLINK v1.90 软件[16]从液相芯片数据中调取出MC1R基因10 个SNP的基因型数据,并计算基因型频率、基因频率和哈代温伯格平衡显著性。 使用HaploView4.1 软件[17]对SNP 进行单倍型分析。
2 结果与分析
2.1 杜黑猪F2 代群体毛色表型分析
杜黑猪F2代个体出现了毛色的分离。 其中,与母本(枣庄黑盖猪)全黑色毛色一致的个体最多(293 头),占总数的75.13%;呈现父本(杜洛克)棕红色毛色的51 头,占总数的13.08%;其余46 头(11.79%)毛色呈现混杂颜色,其中5 头呈黑红条纹,另外41 头毛色既不是黑色也不是棕红色,而是呈棕色偏黄(24 头)或棕色偏白(17 头)。
2.2 杜黑猪F2 代MC1R 基因SNP 基因型和等位基因频率分析
本研究测定的MC1R的10 个SNP 中,5 个SNP(rs337764723、rs786441673、rs326921593、rs705222433、rs691729809)在杜黑F2代群体中只有一种基因型,为单态SNP。 另外5 个多态SNP 的基因型和等位基因信息详见表2。 可以看出,5 个多态SNP的等位基因频率均在0.5 左右,处于哈代温伯格平衡状态(P>0.05),这与试验猪为未经毛色选择的F2代群体有关。 另外,5 个多态SNP 不同基因型的个体数基本一致,推测可能存在紧密连锁。利用HaploView 4.1 进一步分析了5 个多态SNP单倍型,定义了1 个单倍型块(图2)。 单倍型块中各个方块的颜色由浅至深,表示变异位点连锁程度由低到高,深红色表示完全连锁。 结果显示,组成5 个SNP 的各个方块均为深红色,表示它们紧密连锁在一起。 单倍型CGGGT 的频率为0.526,含有该单倍型的个体全部为黑色毛色,单倍型TAAAC 的频率为0.469,含有该单倍型的个体全部为棕红色毛色。

图2 MC1R 基因5 个多态SNP 位点单倍型块
2.3 毛色与基因型关系分析
按照Kijas 等[10]的划分标准,本研究中的5个多态SNP 位点可以划分为ED1和e 等位基因,其中ED1等位基因包含rs45435031、rs45434630、rs45434629 3 个位点, e 等位基因包含rs45435032、rs321432333 2 个位点。 ED1等位基因SNP 位点中,突变纯合基因型(GG、GG、TT 型)的F2代杜黑猪全部为黑色毛色,野生纯合基因型(AA、AA、CC)为棕红色,杂合基因型(GA、GA、TC)个体中大部分(80.83%)为黑色毛色,少部分个体(19.17%)毛色为黑红条纹、棕色偏黄和棕色偏白(表3)。 e 等位基因SNP 位点中,突变纯合基因型(AA、TT)个体全部为棕红毛色,野生纯合基因型(GG、CC)个体全部为黑色毛色,杂合基因型(GA、 TC, AA、 TC, GA、 TT) 大部分个体(80.99%)为黑色毛色,少部分个体(19.01%)毛色为黑红条纹、棕色偏黄和棕色偏白(表3)。
5 个多态SNP 位点在枣庄黑盖猪群体内均只有2 种基因型。 ED1等位基因有突变纯合基因型和杂合基因型,频率分别为83.33%和16.67%;e等位基因有野生纯合基因型和杂合基因型,频率分别为83.33%和16.67%(表4)。
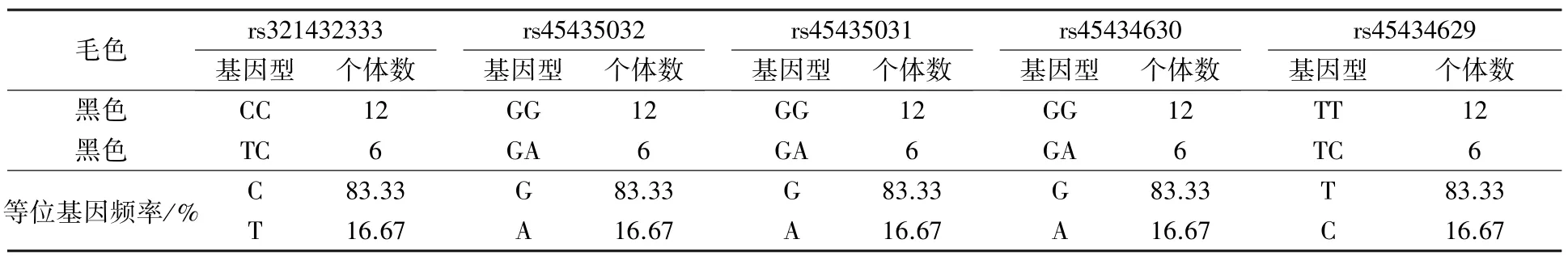
表4 枣庄黑盖猪毛色与SNP 基因型关系
3 讨论与结论
我国绝大多数地方猪种毛色均为黑色,而我国消费者对黑猪肉具有一种与生俱来的青睐感。市场需求决定育种目标,因此,利用国内外两类基因资源,培育优质高效、适应不同市场需求的专门化品系并配套生产黑色优质风味猪成为猪育种领域的主攻目标。 然而,很多黑毛色的专门化品系选育和新品种(配套系)培育过程中后代会出现明显毛色分离,从而影响产品经济价值。 研究毛色关键基因的突变位点与毛色类型的关系,寻找目标毛色的分子标记,可利用分子标记辅助育种快速建立育种目标毛色类型种群。
MC1R基因位于E 座位,定位在猪第6 号染色体短壁的末端[18]。 Kijas 等[10]研究认为ED1等位基因rs45435031、rs45434630 和rs45434629 对应于欧洲大黑猪和中国梅山猪的黑毛色,其中,rs45435031为同义突变,rs45434630 和rs45434629 是错义突变,使所编码的氨基酸Leu 和Val 突变为Pro 和Met。 本研究中枣庄黑盖猪毛色为黑色,大部分个体(83.33%)为ED1等位基因突变基因型(GG、GG和TT),与邓素华[19]、师科荣[20]等的研究结果相一致,表明我国地方猪种的黑毛色可能主要由显性黑等位基因ED1调控。 杜黑猪群体中,ED1等位基因突变基因型个体全部表现为黑色毛色,杂合基因型(GA、GA 和TC)个体中大部分(80.83%)表现为黑色毛色,其他表现为黑红条纹、棕色偏黄和棕色偏白,表明黑色对棕红色为不完全显性,与张建[21]、曾检华[22]等的报道一致。 rs45435032和rs321432333 2 个SNP 位点皆为错义突变,使Ala 分别突变为Thr 和Val,对应于Kijas 等[10]研究中的等位基因e,其突变纯合基因型(AA 和TT)决定了杜洛克猪的棕红毛色。 本研究中,因为没有采集杜黑杂交猪的父本杜洛克样本,故没有检测杜洛克的MC1R基因SNP 基因型。 但杜黑猪F2中棕红色猪全部为突变纯合基因型,与Kijas 等[10]报道的等位基因e 突变纯合基因型影响棕红色毛色的结论一致。
MC1R基因中ED1和e 等位基因的5 个多态SNP 位于444 bp 的序列上,在分析杜黑猪F2个体时发现呈现相似的基因频率,推测可能存在紧密连锁。 利用HaploView 4.1 进行5 个SNP 单倍型分析,结果显示5 个SNP 定义了1 个高度连锁的单倍型块。 单倍型CGGGT 的频率为0.526,含有该单倍型的个体全部为黑色毛色,单倍型TAAAC 的频率为0.469,含有该单倍型的个体全部为棕红色毛色。 该结果提示在杜黑猪新品种选育中,对MC1R基因的5 个SNP 位点进行毛色分子标记辅助选择,有助于快速剔除隐性棕红色等位基因,使杜黑猪新品种的毛色迅速固定为全黑色。
综上,本研究分析了MC1R基因多态性与杜黑猪F2群体毛色的关系,揭示了杜黑猪毛色中黑色对棕红色为不完全显性,利用MC1R基因的5个多态SNP 标记进行分子标记辅助选择可快速剔除隐性棕红色等位基因,为杜黑猪新品种培育中黑色毛色固定奠定基础。
猜你喜欢
生物学杂志(2022年6期)2022-12-17 02:40:42
作文周刊·小学三年级版(2022年28期)2022-07-20 14:02:47
养猪(2021年4期)2021-08-26 10:57:46
大科技·百科新说(2021年12期)2021-03-03 07:22:55
读友·少年文学(清雅版)(2019年1期)2019-05-09 02:35:26
畜牧兽医科学(2018年14期)2018-02-14 01:44:16
畜牧兽医学报(2015年3期)2015-07-05 08:22:51
江南诗(2015年3期)2015-06-01 17:49:57
娃娃画报(2014年9期)2014-10-15 16:31:59
小学生作文·小学中高年级适用(2014年3期)2014-08-18 05:12:23
