
成人胶质母细胞瘤TERT启动子和ATRX突变预后价值☆
2023-11-22 04:10:22阴鲁鑫徐义强肖维汉褚夫政邓李轶黎传清张路东
中国神经精神疾病杂志 2023年8期
阴鲁鑫 徐义强 肖维汉 褚夫政 邓李轶 黎传清 张路东
脑胶质瘤是成人颅内最常见的神经系统原发性恶性肿瘤[1]。胶质母细胞瘤(glioblastoma,GBM)为世界卫生组织(World Health Organization,WHO)定义的4 级脑肿瘤,具有病死率高,预后差的临床特点[2]。近年来随着分子病理学的不断发展,与胶质瘤发生发展相关的分子标记物逐渐被发现[3],分子诊断已成为胶质瘤诊断的重要依据[4]。端粒酶逆转录酶(telomerase reverse transcriptase,TERT)启动子在癌变中起着至关重要的作用[5]。脑胶质瘤中TERT 启动子C228T 和C250T 位点反复突变[6],这些位点的突变被认为是胶质瘤端粒酶激活的主要机制之一[7]。地中海贫血伴精神发育迟滞综合征基因(alpha-thalassemia/mental retardation syndrome Xlinked,ATRX),最早被发现于X 连锁智力迟钝综合征患者中[8]。在胶质瘤中,ATRX的缺失已被证实可促进端粒选择性延长,并与DNA 损伤和应激性复制密切相关[9]。胶质瘤患者体内TERT 启动子、ATRX突变情况对患者预后具有影响。现临床工作中主要依靠免疫组化对胶质瘤患者体内分子标记物进行间接预测。免疫组化作为一种以人主观判断为主的诊断方式,有一定的误差[10],且不能精确的检测出相关基因的突变位点[11]。本研究对GBM患者基因测序结果中TERT启动子、ATRX突变的情况进行分析,旨在发现二者在GBM 中相关性及预后价值,以指导临床诊疗。
1 对象与方法
1.1 研究对象选取2020年1月至2022年1月徐州医科大学附属医院神经外科行手术切除的GBM 患者,随访时间1~36 个月。纳入标准:①初发GBM患者,病理及随访资料较完整;②术前未接受过放化疗及手术治疗;③患者手术记录完善,病理诊断使用组织病理+分子病理相结合的诊断方法,术后病理符合2021 WHO 第5 版中枢神经系统GBM 的诊断标准[12]。排除标准:①年龄<18 周岁的未成年人;②患者纳入后随访无法进行及失联者,或术后未进行过复诊;③严重的心肺功能不全者;④与原发疾病无关的观察期内死亡,如生活意外等。2021 WHO 第5 版中枢神经系统GBM 的诊断标准为:患者分子病理检测结果提示IDH 基因为野生型且伴有TERT 启动子突变,EGFR 扩增,7 号染色体的获得及10号染色体缺失中的一项或者多项(即使在组织学级别较低的情况下),即可诊断为胶质母细胞瘤;或患者分子病理检测结果提示IDH 基因为野生型,且患者组织病理HE 染色切片中观察到肿瘤组织中有坏死或微血管增生的结构(高级别组织形态),也可诊断为胶质母细胞瘤。本研究获我院伦理委员会批准(批文号:XYFY-2022-KL476-01),患者及家属知晓并且签署知情同意书。流程图见图1。
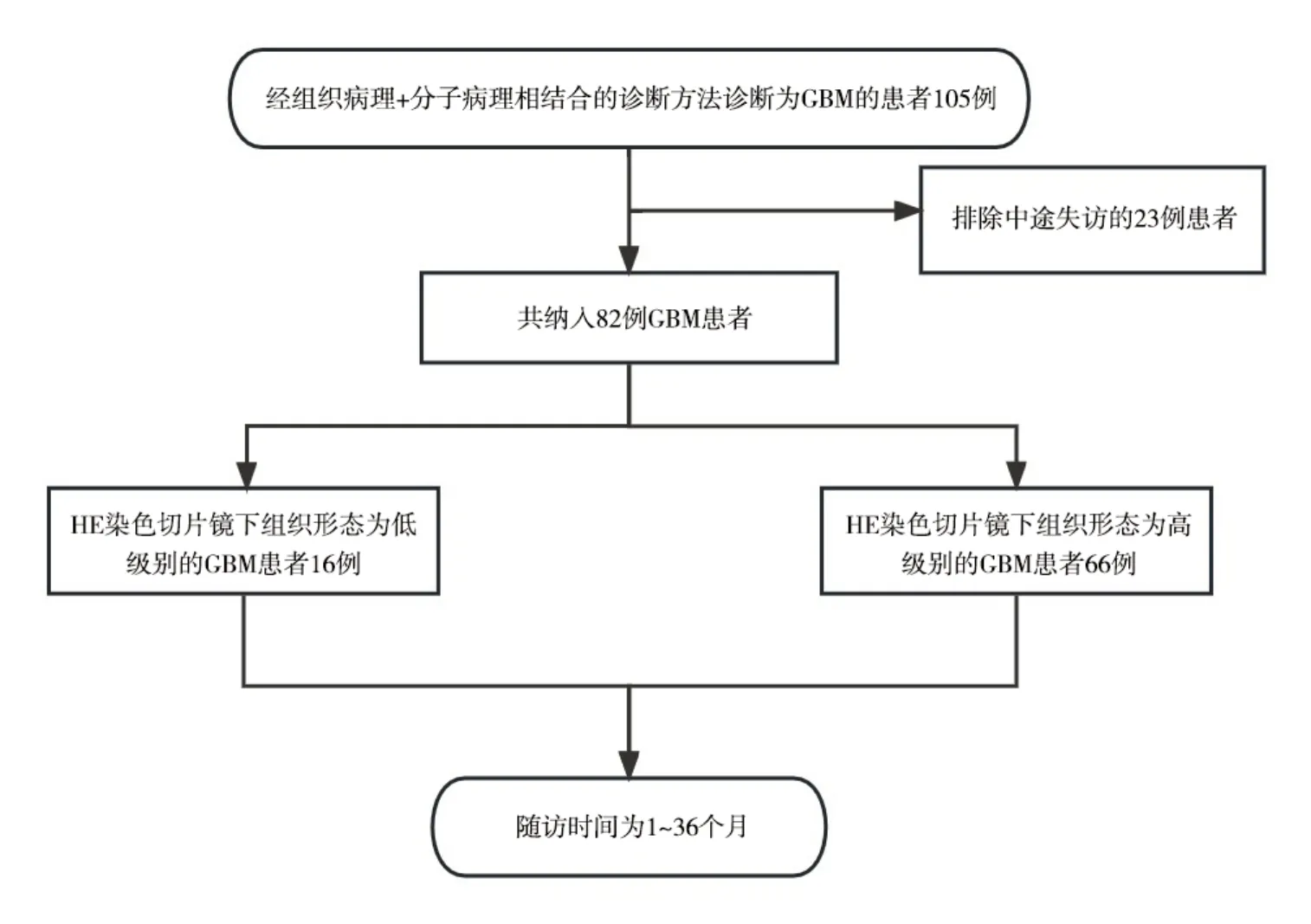
图1 研究流程图
1.2 研究方法
1.2.1 资料收集和分组 通过我院病历系统查询GBM 患者的相关资料,主要包括患者年龄,术前的卡氏功能状态评分(Kamofsky performance status,KPS),肿瘤切除程度,放化疗,手术记录,影像学及病理学资料。挑选符合诊断标准的GBM 患者,回顾分析入组患者的相关临床数据。根据手术记录和术后1 个月内复查的MRI 数据评估肿瘤切除程度。计算肿瘤切除程度(extent of resection,EOR),EOR=(术前肿瘤体积-术后手术体积)/术前肿瘤体积×100%,肿瘤全切除定义为EOR=100%。根据组织病理HE 染色切片的镜下结果将这些GBM 患者分为低级别组织学形态GBM 和高级别组织学形态GBM。WHO 2 级定义为低级别组织学形态;WHO 3级或4级定义为高级别组织学形态。
1.2.2 随访方法 根据我院病历系统采集患者的联系方式对入组患者进行电话随访,每月月底进行1次电话随访,末次随访时间为2023 年2 月,最长随访时间为36 个月,生存时间为手术日期至死亡日期或随访结束日期。共收集105 例胶质母细胞瘤患者,失访23例,失访率21.9%,随访资料完整患者82 例,完整率78.1%。截至到最后随访日期时,72例患者死亡,占随访资料完整患者总人数的87.8%,10 例患者存活,占随访资料完整患者总人数的12.2%。
1.2.3 TERT 启动子及ATRX基因突变的检测 TERT 启动子突变的检测位点包括C228T 和C250T,首先将肿瘤组织中提取的DNA 纯化,经过质检和初步定量确定标本合格后留备用。加入提前设计好的引物和需要的反应液(PCR 缓冲液、DNA 聚合酶、dNTPs、ddH2O)后,将TERT 启动子区经多重PCR 进行扩增,扩增后取适量PCR 产物利用Ion Torrent Proton 平台测序,并对测序结果进行分析。PCR 扩增循环条件为95℃预变性5 min,95℃ 变性15 s,退火60℃ 30s,延伸72℃ 30s,共40个循环,最后72℃总延长5 min。ATRX突变检测使用高通量二代测序的方法(next generation sequencing,NGS),通过探针对ATRX全部外显子区域进行全面捕获,通过标准化建库后,进行PCR 扩增。富集测序模板后使用二代测序仪器进行测序。
1.2.4 肿瘤形态学的检测 脑胶质瘤标本用10%中性甲醛溶液固定后,常规进行脱水,再用石蜡包埋后切4 μm 薄片,行HE 染色后在显微镜下观察胶质瘤的组织学形态。
1.3 统计学方法采用SPSS 26.0 进行分析,不同组织学形态的GBM 中TERT启动子和ATRX突变率的差异采用连续校正χ2检验;GBM患者预后的影响因素采用单多因素COX 回归分析,先将可能影响GBM 患者预后的因素先进行单因素COX 回归分析,后将单因素分析有统计学意义变量进行多因素Cox回归分析。检验水准α=0.05。
2 结果
2.1 一般资料共纳入GBM 患者82 例,男47 例(57.3%),女35 例(42.7%),年龄23~72 岁,平均(60±11)岁;术前KPS 评分20~100 分,中位KPS 评分70(60,80);术中全切除肿瘤63 例(76.8%),次全切除19 例(23.2%),术后接受放疗56 例(68.3%),化疗53例(64.6%),HE 染色切片下具有低级别组织学形态GBM 16 例(19.5%),高级别组织学形态GBM 66 例(80.5%)。TERT 启动子突变的GBM 54 例(65.9%),野生型28 例(34.1%);ATRX 突变GBM 7 例(8.5%),野生型75例(91.5%),病例随访1~36个月。
2.2TERT 启动子和ATRX突变率在不同组织学形态GBM 中的差异 在16 例低级别组织形态的GBM中TERT 启动子突变4 例(25.0%),ATRX突变3 例(18.8%)。在66 例高级别组织形态的GBM 中TERT 启动子突变50 例(75.8%),ATRX突变4 例(6.1%)。不同组织学形态的GBM 中TERT 启动子突变率差异具有统计学意义(χ2=12.584,P<0.01)。但不同组织学形态的GBM 中ATRX突变率的差异无统计学意义(χ2=1.279,P>0.05)。见表1。

表1 不同组织学形态胶质母细胞瘤中TERT启动子、ATRX突变率的比较
2.3GBM患者的预后影响因素分析
2.3.1 单因素分析 单因素COX 回归分析结果 显示,患者年龄(HR=5.084,P<0.001),术前KPS 评分(HR=2.493,P<0.001),术中肿瘤是否全切除(HR=7.926,P<0.001),术后是否放疗(HR=4.840,P<0.001),化疗(HR=3.027,P<0.001),TERT 启动子是否突变(HR=2.943,P<0.001)是影响GBM 患者预后重要影响因素,见图2和表2。

表2 82例GBM患者单因素COX回归生存分析

图2 胶质母细胞瘤患者的单因素COX 生存分析
2.3.2 多因素分析 将上述单因素分析有意义的因素进行多因素COX 回归分析显示,患者年龄,术后是否放疗,化疗,术中肿瘤是否全切,TERT 启动子是否突变是影响GBM患者预后的独立因素(P<0.05)。见表3。

表3 82例GBM患者多因素COX回归生存分析
3 讨论
胶质瘤的发生发展是癌症基因和多因素共同作用的结果[13]。探究影响胶质瘤发生发展的因素,对胶质瘤患者的治疗及预后具有重要意义。
端粒控制着正常细胞的分裂,正常细胞中端粒随着细胞的分裂而缩短,但癌细胞的端粒可以被端粒酶持续拉长[14],从而实现无限分裂。端粒酶由RNA 亚基和逆转录酶(TERT)组成,通过在染色体末端添加六聚体重复序列来维持端粒长度[15]。具有TERT 启动子突变的肿瘤发挥高水平的TERT mRNA 和端粒酶活性,有助于维持端粒长度和细胞增殖[16]。端粒酶参与许多基本的细胞过程,其活性的增加能防止端粒缩短并抑制肿瘤细胞凋亡和衰老,促进肿瘤细胞永生化[17]。高级别组织形态GBM细胞密度显著增高,细胞核异型性及细胞增生极度活跃可能与TERT启动子突变导致肿瘤细胞中端粒酶活性升高致使细胞增生活跃有关。因此TERT启动子突变在GBM 的增殖与进展中中起着至关重要的作用,是判断GBM 患者预后的重要生物标志物[18]。2021 年WHO 第5 版中枢神经系统肿瘤分类标准中将TERT 启动子突变作为GBM 的诊断标准之一[19]。本研究通过对不同组织学形态的GBM 患者的预后进行分析,发现低级别组织学形态的GBM 患者与高级别组织学形态的GBM 患者预后无明显差异。但TERT 启动子突变的GBM 患者预后却较差。这一结果表明GBM 中TERT 启动子状态是一个比组织学诊断更重要的预后标志。且有研究报道称TERT 启动子突变可调节NF-κB 通路,从而促进炎症因子的释放,加重肿瘤进展[20],这可能也是TERT 启动子突变的GBM 患者预后较差的原因之一。
ATRX蛋白是染色质重塑蛋白SWI/SNF2 家族的一部分[21],并与转录辅助因子死亡结构域相关蛋白(death-domain-associated protein,DAXX)结合,通过其在端粒沉积复制中独立的组蛋白变体H3.3 和端粒周围异染色质来维持基因组的稳定性[22]。在胶质瘤中,基本上所有的ATRX突变都可以导致蛋白表达的丧失[23]。虽然近年来在胶质瘤的分子分型方面取得了重大进展,但ATRX在胶质瘤中的确切作用仍不清楚,其对于患者预后的判断仍存在争议。与PEKMEZEI 等[22]研究成果一致,本研究也发现在GBM 患者中ATRX野生患者中位生存期短于ATRX突变患者,但这种生存差异不具有统计学意义,造成这种现象的原因可能与本研究纳入的ATRX突变GBM 患者病例数较少(n=7)有关,后续我们会纳入更多GBM患者进行进一步的研究验证。
TERT 与ATRX均通过控制端粒长度来促进癌症发生和进展[24]。TERT 启动子突变增加了TERT的表达并激活了端粒酶活性[25]。端粒酶的激活可维持端粒的长度,促进肿瘤细胞无限分裂[26]。ATRX突变细胞并不依赖激活端粒酶来延长端粒的长度,这种机制被称为端粒延长替代(alternative lengthening of telomeres,ALT)[27]。ALT 多出现在WHO II 级或III 级的星形细胞瘤中[28]。ALT 的激活与ATP 依赖性解旋酶ATRX或其相互作用配偶体(组蛋白H3.3)的功能丧失有关[29]。本研究发现在高级别组织形态的GBM 中TERT 启动子,ATRX突变多呈负相关,但这种相关性较弱(r=-0.301,P<0.05),因此未在文章中进行赘述。由于TERT 启动子突变和ATRX突变均可导致细胞端粒延长,但所涉及的机制不同。因此,由于功能冗余,神经胶质瘤组织中ATRX的突变和TERT 启动子区的突变可能是相互排斥的。此外,通过对GBM 患者进行生存分析后发现术中肿瘤是否全切是影响患者预后最显著的指标(HR=7.926),在术中全切除肿瘤能够延长GBM患者的总生存期[30]。
本项研究仍存在一些局限性。例如低级别组织形态的GBM 患者相对较少,TERT 启动子突变伴ATRX 突变的GBM 患者病例数少等,在后期的研究中我们会纳入更多的相关患者。
综上所述,TERT启动子与ATRX突变均为胶质瘤中十分重要的分子改变。TERT启动子突变常与GBM 中其他重要的分子标记物相关。因此,TERT启动子突变对GBM 患者的诊断以及预后评估具有重要意义。TERT 启动子突变和ATRX突变在GBM中提示的预后不同。在临床工作中常规检测TERT启动子是否突变并结合其他分子标记物有助于正确评估患者的预后,具有较高的临床应用价值。
猜你喜欢
肝博士(2021年1期)2021-03-29 02:32:08
癌变·畸变·突变(2019年3期)2019-06-03 06:13:04
海南医学(2016年8期)2016-06-08 05:43:00
健康管理(2016年2期)2016-05-30 21:36:03
恋爱婚姻家庭·养生版(2016年5期)2016-05-06 20:18:48
成都中医药大学学报(教育科学版)(2016年1期)2016-01-22 07:24:50
分析测试学报(2015年8期)2016-01-13 06:19:36
听力学及言语疾病杂志(2015年5期)2015-12-24 01:47:05
实用肝脏病杂志(2015年5期)2015-12-03 06:28:04
成都中医药大学学报(教育科学版)(2014年1期)2014-01-19 13:58:28
