
深层页岩干酪根纳米孔隙中甲烷微观赋存特征
2023-11-17 12:01冯鑫霓吴建发杨学锋
石油钻探技术 2023年5期
黄 亮,冯鑫霓,杨 琴,吴建发,杨学锋,黄 山
(1.油气藏地质及开发工程全国重点实验室(成都理工大学),四川成都 610059;2.成都理工大学能源学院,四川成都 610059;3.中国石油西南油气田分公司页岩气研究院,四川成都 610051)
四川盆地页岩气资源丰富,其中埋藏深度大于3 500 m 的超压深层页岩储层具备年产300×108m3页岩气的能力,是页岩气未来产能增长的重要领域[1-2]。与中深层页岩相比,深层页岩在岩石组成、孔隙结构、流体赋存和温度压力条件等方面存在显著差异[3]。受工程技术的限制,埋藏深度3 500~4 000 m 的页岩气尚未实现大规模商业化开采[4]。明确深层页岩纳米孔隙中气体微观赋存特征,有助于指导深层页岩气资源评估和开发设计[5-6]。页岩气主要以游离态、吸附态和溶解态赋存于复杂孔隙空间[7-8]。室内试验是研究页岩气赋存的重要手段,多位学者基于等温吸附试验探究了页岩气吸附规律。李倩文等人[9]在0~20 MPa 压力范围内量化了页岩气不同赋存状态的含量,并建立了页岩气赋存状态动态演化模型。唐鑫等人[10]基于等温吸附试验,在压力0~10 MPa、温度303.15~363.15 K 条件下探究了龙马溪组页岩孔隙结构中甲烷的赋存特征。纪文明等人[11]采用静态容积法,测定了页岩在压力0~11 MPa、温度20~100 ℃条件下的气体吸附规律。基于等温吸附试验探究页岩气吸附规律已较为成熟,但受试验设备条件限制,无法进行深层页岩储层高温高压条件试验。
相比室内吸附试验,分子模拟可以基于实际深层页岩储层高温高压条件开展,且能够揭示页岩气不同状态的赋存机理。石钰等人[12]采用分子模拟方法,揭示了多组分气体在页岩干酪根中的竞争吸附特征。黄亮等人[13-15]基于分子模拟方法,揭示了有机质类型和热成熟演化对甲烷/二氧化碳多组分吸附的控制机理,并阐明了干酪根吸附-膨胀耦合机理。方镕慧等人[16]模拟研究了温度-压力耦合作用下的页岩气吸附规律,对比了不同地质模型下的吸附特征差异。此外,页岩有机质是气体赋存的主要介质,多位学者对不同类型的有机质分子模型进行了研究[17-20],但主要针对油页岩有机质和中深层页岩有机质,其对川南地区深层页岩储层有机质的适用性有待验证。目前,基于深层页岩有机质分子模型开展高温高压下页岩气赋存相关模拟的研究尚未见到报道,深层页岩气微观赋存机理有待明确。
为此,笔者基于龙马溪组深层页岩干酪根结构单元,采用切割法和叠合法相结合的方法,构建了干酪根三维纳米孔隙模型;采用耦合分子动力学算法和巨正则蒙特卡洛算法,开展了高温高压条件下甲烷赋存模拟,分析了不同因素对甲烷赋存量的影响规律;探讨了甲烷在纳米孔隙中的赋存机理,揭示了甲烷微观分布特征、甲烷-壁面微观作用特征及甲烷优先吸附位。
1 孔隙模型构建及吸附特征
1.1 干酪根纳米孔隙分子模型构建
笔者先前通过有机地球化学测试表征并构建了川南泸州区块深层页岩龙马溪组干酪根的分子结构模型,干酪根三维基质结构密度及狭缝状孔隙中甲烷等温吸附曲线与试验结果吻合较好[13]。基于先前构建的龙马溪组干酪根基质结构,构建深层页岩干酪根不同形状和孔径的纳米孔隙分子模型。
首先,将龙马溪组干酪根三维基质结构模型沿z方向扩展1 倍,扩展后的干酪根基质模型如图1(a)所示,其模型尺寸为3.27 nm×3.27 nm×6.54 nm。然后,采用切除法在扩展干酪根基质模型中心分别创建狭缝状孔隙和圆管状孔隙:在基质模型中心切除基质块体,形成狭缝状孔隙(见图1(b));在基质模型中心选定球心和半径,沿x方向不断移动球心并切除球体,形成圆管状孔隙(见图1(c))。切除法会导致干酪根形成部分断键,自动补氢饱和并删除结构中多余的氢原子。
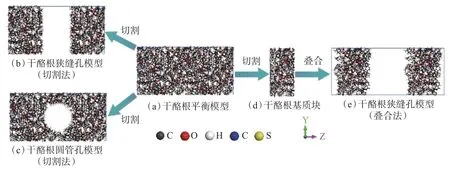
图1 干酪根纳米孔隙模型及构建流程Fig.1 Kerogen nanopore model and construction process
为研究孔隙尺寸对页岩气赋存行为的影响,需构建不同孔径的干酪根纳米孔隙模型。切除法会在干酪根模型中形成不同厚度的基质孔壁,干扰孔径的影响规律。为消除基质孔壁厚度的影响,采用叠合法构建不同狭缝孔径的干酪根纳米孔隙模型。叠合法基于切除法中的干酪根基质块体(见图1(d)),将其在z 方向叠合形成狭缝状孔隙,通过调整基质块体的距离,形成不同孔径的狭缝状孔隙模型(见图1(e))。研究构建了4 种狭缝孔径的干酪根纳米孔隙模型,狭缝孔径分别为2.5,3.0,4.0 和6.0 nm,各干酪根模型的基质部分相同。
1.2 甲烷吸附特征
基于LAMMPS 分子模拟程序,开展干酪根孔隙中甲烷赋存模拟。干酪根分子模型用CVFF 力场[21]描述,甲烷分子用TraPPE 联合原子力场[22]描述,不同原子间的非键相互作用采用L-J 12-6 势能函数描述。
采用巨正则蒙特卡洛和分子动力学耦合的方法(GCMC-MD 方法)模拟甲烷赋存平衡。GCMC-MD耦合模拟由10 000 个MD 步和2 500 个GCMC 交换步循环迭代构成,循环迭代直到体系的温度、能量项及甲烷分子数达到平衡。模拟在正则系综下开展,温度由Nosé-Hoover 法[23]控制;干酪根纳米孔隙结构在模拟中保持固定,甲烷可自由运动;模拟体系采用周期性边界条件,截断半径为1.40 nm;模拟时间步长为1 fs。
甲烷赋存模拟中体系压力由输入的化学势确定,化学势与压力的关系见文献[24]。赋存平衡后,对孔隙模型中的甲烷分子数开展系综平均得到甲烷总气体量,其标准误差小于5%。甲烷过剩吸着量由总气体量转换得到:
式中:ne为过剩吸着量,mmol/g;nt为总气体量,mmol/g;ρ为甲烷气体的体相密度,g/cm3;Vp为模型中甲烷的可接触孔隙体积,mmol·cm3/g2。
2 甲烷赋存量的影响因素分析
甲烷赋存量受压力、温度、孔径大小和孔隙形状等因素的影响,采用GCMC-MD 耦合方法,模拟干酪根孔隙中甲烷在深层高温高压下的赋存平衡,对比分析不同条件下的甲烷总气体量和过剩吸着量,以明确甲烷的微观赋存规律[25]。
2.1 压力和温度
基于切割法构建孔径为2.0 nm 的干酪根狭缝孔模型,开展温度为90,120,150 ℃和压力0~90 MPa下的甲烷赋存模拟,甲烷在干酪根孔隙中的吸附曲线如图2 所示。从图2 可见,随着温度升高,干酪根中甲烷总气体量在研究压力范围内单调降低(见图2(a))。甲烷总气体量包括孔隙表面的吸附气、孔隙中间的游离气以及干酪根基质内的溶解气。高温下甲烷分子运动加剧,当甲烷分子之间的相互作用强度降低时,甲烷体相密度减小,游离气量降低;当甲烷分子与孔壁分子的相互作用强度降低时,甲烷逃离干酪根外表面和超微孔内部束缚的能力增强,吸附气量和溶解气量降低。
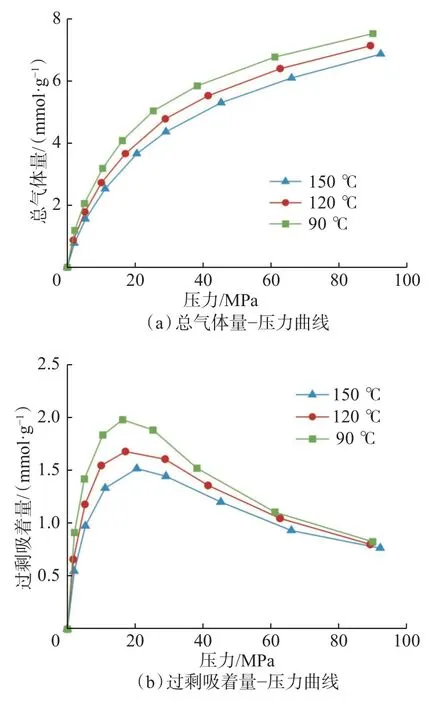
图2 不同温度下干酪根孔隙的甲烷吸着量曲线Fig.2 Methane sorption curves of kerogen model at different temperatures
过剩吸着量反映绝对吸附气和溶解气相比对应空隙空间内游离气多余的气体附着量,受气体分子与孔壁分子的相互作用强度控制。随压力升高,不同温度下甲烷过剩吸着量差值呈先增大后减小的趋势(见图2(b))。压力较低时,甲烷过剩吸着量在不同温度下的差异较小。这是因为低压下干酪根表面和超微孔隙内吸着空位丰富,气体优先以吸附和溶解状态赋存,甲烷与壁面的相互作用强度较大,导致温度对吸着气体逃逸能力的影响较小。高压条件下,干酪根模型中大部分吸着空位被充填,壁面分子与空位外侧甲烷分子的相互作用强度较低,温度对其影响程度接近于对体相游离气的影响程度。因此,干酪根模型中绝对吸着量和游离气量的下降幅度相近,使过剩吸着量基本不受温度影响。深层高压下,温度升高造成的总气体减小量由绝对吸着减小量和游离气减小量等额贡献。
与伊利石孔隙中温度的影响类似[24],干酪根中甲烷过剩吸着量在压力约为20 MPa 时达到最大,且最大过剩吸着量对应的压力随温度降低而略微降低。随着压力升高,甲烷绝对吸着气量逐渐增加。低压下,由于吸着空位较多,绝对吸着气量增加速度大于游离气量增加速度,使过剩吸着气量逐渐增加。压力约为20 MPa 时,绝对吸着气量与游离气量的增加速度差异达到最大;随着压力继续升高,由于吸着空位逐渐趋于饱和,绝对吸着气量增加速度小于游离气量增加速度,导致过剩吸着气量逐渐减小。低温条件下,甲烷分子与孔壁分子的相互作用强度更大,绝对吸着气量增加速度在更低压力下达到最大。
2.2 孔径影响
在温度120 ℃、压力分别为17.1 和89.2 MPa 条件下,开展了干酪根中甲烷赋存分子模拟,对比孔径范围2.5~6.0 nm 干酪根的甲烷吸着量(见图3)。

图3 不同干酪根孔径的甲烷吸着量Fig.3 Methane sorption capacity under different kerogen pore sizes
从图3 可以看出,随着干酪根孔径增加,甲烷总气体量逐渐增加,而过剩吸着量无明显变化规律[26]。在相同温度和压力条件下,甲烷体相密度不变,孔径增大使体相赋存空间增大,导致甲烷游离气量增加。干酪根介孔孔径范围2.5~6.0 nm 条件下,吸着气量基本不受干酪根孔径的影响,孔径增大造成的总气体量增加主要由游离气贡献。
相比中深层页岩储层压力(17.1 MPa)条件,深层高压(89.2 MPa)条件下的甲烷总气体量更大,过剩吸着量更小。压力由17.1 MPa 增大至89.2 MPa,干酪根纳米孔隙中甲烷游离气的增加幅度大于吸着气,导致吸着气在总气体量中的占比减小。
2.3 孔隙形状影响
干酪根孔隙的形状不仅影响孔隙的容积和表面积,还影响孔隙壁面与气体分子的相互作用强度,进而影响气体赋存特征[27],笔者基于干酪根中最基本的圆管孔和狭缝孔分析孔隙形状对甲烷赋存量的影响规律。甲烷赋存量可以以mmol/m2、mmol/m3和mmol/g 等多种单位表示。温度120 ℃条件下,不同赋存量单位下甲烷在不同孔隙结构(形状和孔径)干酪根中的赋存量如图4 所示。从图4 可以看出,甲烷赋存量单位对干酪根孔隙中甲烷的赋存规律具有显著影响。赋存量单位为mmol/m2时,不同孔隙形状下甲烷总气体量受孔径的影响(见图4(a));赋存量单位为mmol/m3时,不同孔隙形状下甲烷过剩吸着量受孔径和压力的影响(见图4(d));赋存量单位为mmol/g 时,不同孔隙形状下甲烷过剩吸着量受孔隙容积和表面积的影响(见图4(f))。因此,为分析孔隙形状对甲烷赋存的影响规律,需针对特定的甲烷赋存状态采用不同的赋存量单位。
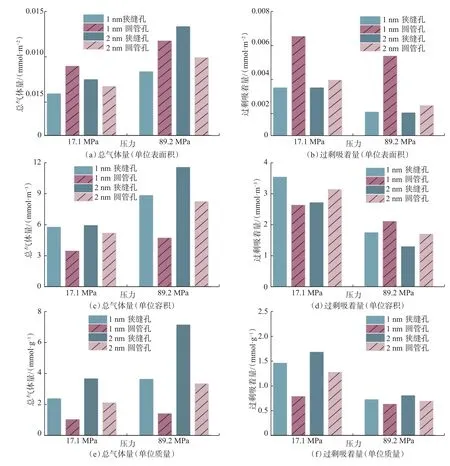
图4 不同赋存量单位下孔隙结构的甲烷吸着量Fig.4 Methane sorption capacity under different occurrence units and pore structures
干酪根孔隙中的甲烷总气体量受甲烷分子间相互作用以及甲烷分子与孔壁分子间相互作用共同控制,对比总气体量需要同时考虑不同形状孔隙容积和表面积的差异,因此单位宜采用mmol/g。干酪根孔隙中的甲烷过剩吸着量主要受甲烷分子与孔壁分子间相互作用控制,过剩吸着量单位宜采用mmol/m2,以消除不同形状孔隙表面积不同的影响。由图4(e)和图4(b)可以看出,干酪根狭缝孔中甲烷总气体量大于对应的圆管孔,而狭缝孔中甲烷过剩吸着量小于对应的圆管孔,孔隙形状对甲烷赋存量的影响规律在研究的孔径和压力下类似。单位质量干酪根模型中,相比圆管孔,狭缝孔的容积和表面积更大,促成更大的甲烷气体量。相同干酪根孔隙表面积下,相比狭缝孔,圆管孔的孔壁与甲烷分子的相互作用强度更大,造成甲烷过剩吸着量更大。
3 甲烷微观赋存机理研究
3.1 甲烷微观分布特征
在压力89.2 MPa、温度分别为90,120 和150 ℃条件下,通过分子模拟得到孔径为2.0 nm 的干酪根的甲烷密度剖面和吸附相密度(见图5)。从图5(a)可以看出:孔径为2.0 nm 的干酪根狭缝孔隙中存在多个甲烷密度峰,其中靠近狭缝壁面的第1 密度峰最为明显,其密度远大于对应温度和压力的NIST体相结果,可被识别为甲烷吸附层;第2 密度峰的密度略大于对应的NIST 体相结果,可被识别为过渡层[22];孔隙中央区域第3 密度峰的密度接近于NIST体相结果,可被识别为甲烷游离相。此外,干酪根基质内部还存在密集分布的甲烷密度峰,其密度远低于NIST 体相结果,可被识别为甲烷溶解相。
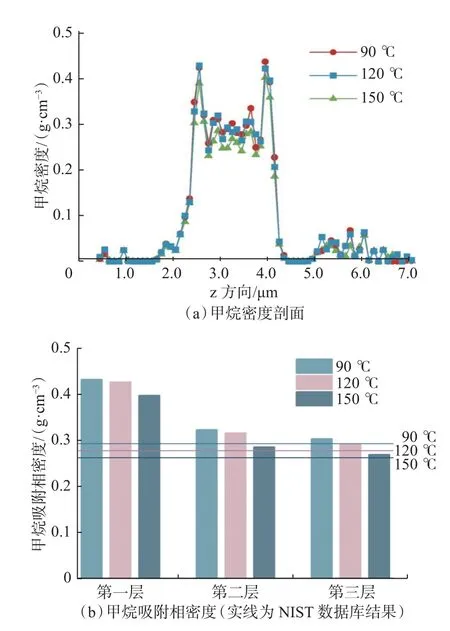
图5 不同温度下甲烷密度剖面和吸附相密度Fig.5 Methane density profile and adsorption phase density at different temperatures
在深层高压条件下(89.2 MPa),随温度升高,干酪根狭缝孔隙中甲烷第1 密度峰、第2 密度峰和第3 密度峰的密度单调下降。其中,吸附层密度和游离相密度的下降幅度接近,其主要原因是高压下甲烷过剩吸着量受温度影响不明显。此外,不同温度下干酪根基质内部的甲烷溶解相密度差异较小,温度升高对甲烷溶解相的影响不明显。
在温度120 ℃、压力89.2 MPa 条件下,通过分子模拟对比孔径范围2.5~6.0 nm 干酪根的甲烷密度剖面和吸附相密度,结果如图6 所示。从图6 可以看出:随着介孔孔径增大,狭缝孔隙内2 个壁面对甲烷分子的耦合相互作用减弱,狭缝孔壁面主要为单层吸附,孔径为2.0 nm 狭缝孔壁面的过渡层趋于退化或消失;在研究的介孔孔径范围内,不同孔径下甲烷吸附相密度无明显差异。
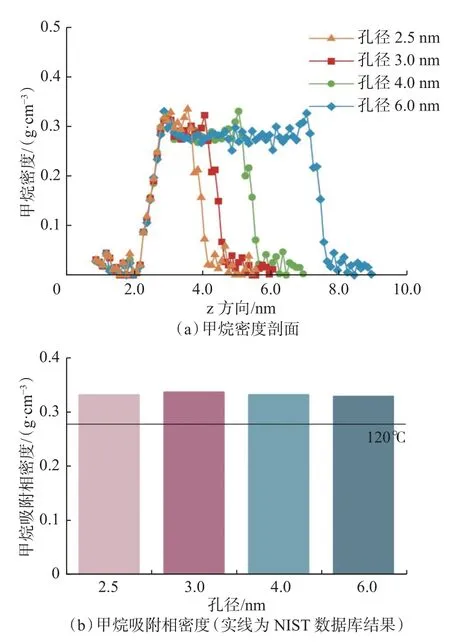
图6 不同孔径下甲烷密度剖面和吸附相密度Fig.6 Methane density profile and adsorption phase density at different pore sizes
3.2 甲烷-壁面微观作用特征
干酪根孔隙模型中,甲烷分子与孔壁分子之间存在非键相互作用,总相互作用能为模型中所有甲烷分子与孔壁分子的相互作用能之和,受甲烷分子数量影响。比作用能是孔壁分子与单个甲烷分子的平均相互作用能,可反映甲烷与孔壁的平均作用强度。作用能为负,代表组分间相互吸引;作用能为正,代表组分间相互排斥。图7(a)为甲烷-孔壁在不同压力下的总作用能和比作用能,均为负值,表明甲烷分子和孔壁分子相互吸引;随着压力升高,总作用能绝对值单调增大,比作用能绝对值呈减小趋势。压力低于20 MPa 时,总作用能和比作用能急剧变化;压力大于20 MPa 时,总作用能近似呈线性增加,比作用能降低幅度变小。甲烷分子优先吸附于孔壁的高能位点,压力升高使甲烷分子数增多,甲烷逐渐在较低能量位点吸附。孔壁上的吸附位点由高能到低能逐步被占据,使比作用能降低,而甲烷吸附分子数逐步增多使总作用能增大。深层高压下,甲烷吸附相趋于饱和,压力升高主要增加游离相中甲烷分子数。
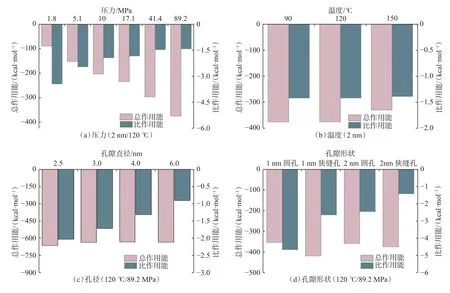
图7 甲烷-孔壁的总作用能和比作用能Fig.7 Total action energy and specific action energy of methane and pore wall
图7(b)为不同温度下甲烷-孔壁间的总作用能和比作用能(压力约90 MPa)。随着温度升高,总作用能绝对值呈减小趋势,主要是因为孔隙中甲烷总气体量随温度升高而逐渐减小(见图2(a));不同温度下比作用能的差异较小,甲烷与孔壁的平均作用强度变化较小,这是由于高压下温度升高,使孔隙中吸附相气体数量和游离相气体数量近似同步减少(见图5(b))。
图7(c)为不同孔径下甲烷-孔壁的总作用能和比作用能。在孔径2.5 nm 干酪根狭缝孔隙中,两侧孔壁对孔隙内甲烷分子具有较强的耦合作用,使总作用能绝对值较大。在更大孔径下,孔壁的耦合作用消失,孔隙中央游离相中甲烷分子增多对总作用能的贡献较小,导致总作用能变化较小。随着孔径增大,比作用能近似呈线性下降,游离相甲烷分子增多,使甲烷与孔壁的平均作用强度降低。
图7(d)为不同孔隙形状下的总作用能和比作用能。在相同孔径下,狭缝孔内总作用能的绝对值大于圆管孔,这是由于狭缝孔模型中孔隙表面积更大,甲烷赋存量更大。相比狭缝孔模型,圆管孔内比作用能的绝对值更大,甲烷分子与圆管孔壁的平均相互作用强度更大。
3.3 甲烷优先吸附位
干酪根具有多种不同类型的基团,不同基团对甲烷的亲和性存在差异。在温度120 ℃、压力89.2 MPa条件下,通过分子模拟计算得到甲烷分子与干酪根上不同基团的径向分布函数(见图8),分析甲烷在干酪根结构中的优先吸附位点。径向分布函数曲线中第一个作用峰的高度可以反映两粒子间亲和性的强弱,作用峰的高度越大,亲和性越强。由图8(a)可知,干酪根中含硫基团是甲烷分子的优先吸附位点。另外,甲烷分子与干酪根中含氧和含氮基团在0.345 nm 处具有明显的作用峰,表明含氧和含氮基团对甲烷的潜在亲和性较强。为明确干酪根中甲烷的优先吸附位点,分别计算了甲烷分子与不同含氧、含氮和含硫基团的径向分布函数曲线(见图8(b)—(d))。由图8(b)—(d)可知,含硫基团中的噻吩是甲烷分子在干酪根中的优先吸附位点,含氧基团中的羟基对甲烷分子的亲和性较强,含氮基团中无明显的甲烷优先吸附位点。

图8 甲烷分子与干酪根不同类型基团的径向分布函数特征曲线Fig.8 Characteristic curves of radial distribution functions of methane molecules and different types of kerogen groups
干酪根二维结构单元和三维结构模型中甲烷的优先吸附基团分布如图9 和图10 所示。由图9 和图10 可以看出干酪根狭缝孔隙表面和基质内部均分布大量的甲烷优先吸附位点,甲烷分子既能以吸附相状态赋存于孔隙表面,也能以溶解相状态赋存于干酪根基质内部。
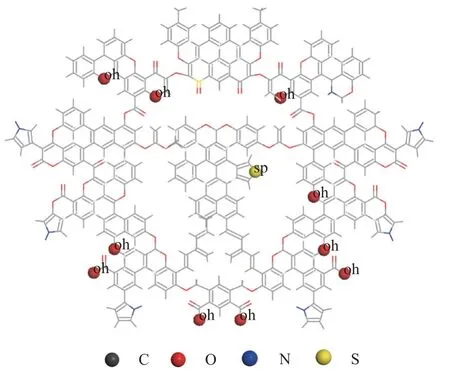
图9 干酪根结构单元中甲烷优先吸附基团Fig.9 Preferential adsorption groups for methane in kerogen structural units

图10 孔径2.0 nm 的干酪根三维狭缝孔模型中甲烷优先吸附基团Fig.10 Preferential adsorption groups for methane in 3D slit-like pore model of kerogen with a pore size of2.0 nm
4 结论
1)深层高压下温度升高造成甲烷绝对吸着量和游离气量等额减小,这与前人的研究结果相吻合。相比中深层压力,深层页岩高压下甲烷总气体量更大,过剩吸着量更小,吸着气在总气体量中的占比更小。
2)干酪根介孔孔径增加使游离相体积增加,造成总气体量增大,游离相甲烷分子增多使甲烷与孔壁的平均作用强度降低。
3)干酪根中甲烷总气体量单位宜采用单位质量单位,过剩吸着量单位宜采用单位表面积单位,孔隙形状对甲烷赋存量的影响规律在研究孔径和压力下类似。
4)研究的干酪根结构中,含硫基团中的噻吩是甲烷分子的优先吸附位点,含氧基团中的羟基对甲烷分子的亲和性较强,含氮基团中无明显的甲烷优先吸附位点。
猜你喜欢
地球化学(2022年6期)2023-01-03
油气藏评价与开发(2022年3期)2022-06-23
实验室研究与探索(2021年4期)2021-05-29
军民两用技术与产品(2021年10期)2021-03-16
科技创新与应用(2021年7期)2021-02-04
中外公路(2020年4期)2020-09-14
水上消防(2020年1期)2020-07-24
建材发展导向(2019年5期)2019-09-09
疯狂英语·新读写(2018年3期)2018-11-29
自动化与仪表(2014年10期)2014-02-26
