
Forest composition and red oak(Quercus sp.)response to elevation gradients across greentree reservoirs
2023-11-15 07:56CssndrHugPrdipSudKeithMcKnightDouglsOsorne
Forest Ecosystems 2023年5期
Cssndr Hug, Prdip Sud,*, Keith McKnight, Dougls C.Osorne,c
a College of Forestry, Agricultural, and Natural Resources, University of Arkansas at Monticello, Monticello, AR, 71656, USA
b USFWS/Lower Mississippi Valley Joint Venture, Tyler, TX, 75707, USA
c Five Oaks Ag Research and Education Center, Humphrey, AR, 72073, USA
Keywords:Bottomland hardwoods Diversity Forested wetland Overstory Red oak Soil Thinning and management
ABSTRACT Elevation gradients within forested wetlands have long been recognized for their role in defining species composition through factors such as hydrology and soil characteristics.Greentree reservoirs (GTRs) are leveeimpounded tracts of bottomland hardwood forest flooded throughout the winter months to provide habitat for overwintering waterfowl.Artificial flooding of GTRs alters the forest composition due to flood frequency, depth,and duration in combination with slight changes in topography.To evaluate the effect of elevation gradients,soil properties, and management techniques in the overstory species composition and red oak (Quercus spp.) species abundance, we inventoried 662 plots across 12 independent GTRs in eastern Arkansas.In the lower elevations ranging from 50.98 to 54.99 m above sea level, the importance value index (IVI) was highest for nuttall oak(Quercus texana)and overcup oak(Quercus lyrata),whereas IVI shifted to cherrybark oak (Quercus pagoda)in the higher elevations ranging from 54.99 to 58.00 m.Alpha diversity did not differ by elevation gradient, soil property, or management technique within GTRs.Beta diversity, using non-metric multi-dimensional scaling(NMDS) analysis, indicated site-specific variability significantly correlated with the environmental predictors,including elevation (R2 = 0.57), easting (R2 = 0.47), soil texture (R2 = 0.21), and pH (R2 = 0.12).Red oak species-specific mixed-effects modeling of abundance response using Poisson distribution suggested an inverse correlation of nuttall oak and a direct correlation of cherrybark oak abundance with elevation.However, willow oak (Quercus phellos) abundance was not significantly affected by elevation but was by silt loam soil texture and restoration management techniques.These findings will aid management efforts to reduce the dominance of less desirable species that are prominent under specific environmental conditions and promote the dominance of more desirable species.Ultimately GTR sustainability is increasingly important amid the unpredictable impacts of climate change on the preferred red oak species that are economically,ecologically,and environmentally valuable to the sustaining economy of the local community and managing habitats for wildlife.
1.Introduction
Elevational gradients across a landscape have long been recognized for their role in driving tree species composition (Whittaker, 1956;Hodges and Switzer, 1979; Gould et al., 2006; Malizia et al., 2020).Depending on the ecosystem, elevation can impact species composition through biotic and abiotic factors.Biotic interactions can be studied interspecifically,often by examining plant phenology at differing elevations(Halbritter et al.,2018;Rafferty et al.,2020).Species composition can also be studied intraspecifically, for instance, diversity in seedling species,overstory species,herbaceous species,and sapling species differs significantly by elevation class (Carpenter, 2005; Grell et al., 2005);variation in the growth and establishment of the saplings of deciduous forest species predicts forest composition change (Kobe, 1996).A meta analysis showed that such change could account for 25%of the total trait variation within communities (Siefert et al., 2015) and 32% variation among plant communities.Luo et al.(2016)reported that environmental factors could explain 50%-76% of the variation in individual species traits along the elevational gradient.Across elevation gradients, abiotic environmental factors such as temperature,soil composition,availability of space, and precipitation vary, leading to changes in species diversity and composition (Stage and Salas, 2007; Sundqvist et al., 2013; Morin et al.,2018).
Depending on the environmental factors, even small changes in topography can lead to slight changes or entirely unique species composition.In regions where the topography is subtle and elevational changes are small, such as the Mississippi Alluvial Valley (MAV), the most extensive floodplain in the USA, which covers 9.7 million ha,floodwater plays a significant role in shaping the elevation gradient(Grell et al.,2005;Lockhart et al.,2010).Due to continuous flooding,the speed and volume of floodwater create ridges, flats, and sloughs by carving out and depositing varying degrees of sediments,which leads to changing elevation gradients across the floodplains (Hodges, 1997;Twilley et al., 2016).Resulting from elevational changes, flood water depth and duration, and soil saturation contribute to differences in the arrangement of tree species in the lowland floodplain (Smith and Linnartz,1980;Huffman and Forsythe,1981).Species along ridges may be slightly more limited by water and higher in diversity,while those found in the sloughs may be more adapted to tolerating excess water conditions and low in diversity (Meadows and Stanturf, 1997; Grell et al., 2005).Such shifts in forest composition due to hydrogeomorphology within the MAV can result from a subtle elevation variation of less than a half meter above sea level within bottomland hardwood forests (Meadows and Hodges,1997).
Bottomland hardwood forests (BHF) are forested wetlands found mainly in alluvial floodplains along river systems in the southeastern United States.These BHFs provide crucial ecosystem services such as atmospheric carbon sink, flood and groundwater storage, improvement of water quality,and wildlife habitat(Schmidt et al.,2014;Mahaffey and Evans, 2016).Throughout the MAV, approximately 2.67 million ha of BHF have been deforested since 1937 (Macdonald et al., 1979).The effects of this widespread destruction of BHF have been documented as an increase in soil erosion and loss of native biotic diversity (Harris and Gosselink,2020).With less habitat and resources for species,there is also a decline in overwintering populations of waterfowl and forest-dependent songbirds along the Mississippi Flyway (Harris and Gosselink, 2020; Rosenberg et al., 2019; Meehan et al., 2021).Water quality is also reduced as a result of fewer nutrients from upland and agricultural runoff being filtered through and retained in BHF(Harris and Gosselink, 2020).However, much of the remnant BHFs are impounded by levee systems in managed areas known as greentree reservoirs(GTR).
Greentree reservoirs were initially developed in Stuttgart, Arkansas,in the 1930s as BHFs were artificially flooded by public or private landowners (Wigley and Filer, 1989).This flooding is done primarily through the use of wells and pumps(Allen et al.,2004)and is designed to attract waterfowl by flooding impoundments during fall and winter seasons for hunting opportunities(Wigley and Filer,1989;Keeland et al.,2010).Along with the ecosystem functions found within BHFs,GTRs also serve as habitats for other game species including white-tailed deer(Odocoileus virginianus)and wild turkey(Meleagris gallopavo)(Heitmeyer,2006), and provide a source of timber.However, the current management practices used in many GTRs for hunting, timber production, and forest health are of major concern(Spears et al.,2022).There is a paucity of information concerning key characteristics of GTRs, particularly regarding management practices that could promote both ecosystem services and economic value (Wigley and Filer, 1989).Currently, most GTR management techniques involve constant and consistent flooding throughout the winter season, which is not congruous with the natural,irregular flood-pulse patterns historically occurring in BHFs.Many GTRs are often flooded for up to two and a half months longer than unimpounded BHFs (Ervin et al., 2006).Evidence demonstrates that such constant yearly flooding alters the forest community type and species composition (Deller and Baldassarre, 1998; Ervin et al., 2006; Keeland et al.,2010).
Plant species occurring in GTRs arrange themselves according to varying degrees of flood tolerance(Mcknight et al.,1981;Hodges,1997).Over time, forest compositional shifts can be seen in increased water-tolerant plant species, especially within lower elevation sites.These species, such as overcup oak (Quercus lyrata) and water hickory(Carya aquatica) are less desirable for waterfowl and timber production(Fredrickson, 1979; Hook,1984).Red oaks (Quercus spp.), which are of high economic value and desirable for timber harvesting and food (i.e.acorn)(Straub et al.,2019) for waterfowl, may be particularly susceptible to GTR flooding techniques(McKnight et al.,1981;Allen et al.,2004).They tend to be less water tolerant than other species found within BHF,often establishing along ridges or slightly higher elevations (Hodges,1997;Hodges and Switzer,1979; Lockhart et al.,2010).
This variation in forest composition, shaped by flooding, combined with common GTR management practices, poses a threat to the natural wildlife dependent on this habitat.The abundance and species richness of songbirds have been shown to be significantly lower in GTRs than naturally flooded BHF, in part due to the thinned understory, which reduces crucial breeding habitat for songbirds (Christman, 1984).Additionally,the diet of overwintering mallards(Anas platyrhynchos)has been shown to be 48%red oak species,whereas wood ducks(Aix sponsa)have been shown to occasionally eat white oak (Quercus sp.) acorns (Allen,1980).As many of these GTRs shift in forest composition, particularly away from red oak species,we will likely see changes in wildlife as well,causing concern for how best to manage GTRs.This has recently resulted in an increased desire for managers to make necessary management changes(Spears et al.,2022).
One management technique commonly used to promote the health and abundance of desirable tree species is thinning.Thinning is often recommended in order to increase gaps in the overstory canopy to promote the generation of shade intolerant species like oaks,but it is shown to have limited results, especially commercially (Sander, 1979; Loftis,1983).In addition, management of hydrology through the use of water control structures is crucial to moving water off and on GTRs in a way which promotes the health of desirable species.The lower areas in a GTR may trap water for longer periods,causing increased harm to less water tolerant tree species (Moorhead et al., 1991).By altering management techniques, land owners will be able to maintain healthier and more sustainable forests,to promote economic interest within GTRs to benefit local and nationwide economies.
Waterfowl hunting and timber extraction are two major contributors to the economic value of GTRs.Almost ninety eight percent of funds from federal duck stamps go directly to the National Wildlife Refuge system,much of which aids in acquiring wetland habitat(U.S.Fish and Wildlife Service,2011).In Arkansas,waterfowl hunting generates approximately$1 million per day during waterfowl hunting season, which benefits a multitude of local businesses (Zellers, 2021).Additionally, the production and extraction of timber from BHF throughout the United States is valued at $3-8 billion annually, and Arkansas is the 9th leading timber producer in the United States(Harris and Gosselink,2020).Harvesting of timber, especially red oak species, within GTRs contributes to this and highlights the potential economic importance of GTR management in the region.However,actionable information on the long-term effect of GTR management on vegetation composition is limited within both state and federally-owned properties and is largely lacking in privately owned properties.
This study aims to increase the understanding of the role of elevation and GTR management on tree species composition.The study site is located within the property of Five Oaks, and was selected because the GTRs are managed for duck hunting,which offers a prime opportunity to evaluate the species composition at fine scale elevations along with management practice.Our primary objectives for this study were to, 1)evaluate variation in overstory species composition across elevation gradients, and management strategies; and 2) analyze the effects of management and environmental variables on the abundance of three red oak species preferred by wintering ducks.By understanding forest composition, particularly of red oaks, across the landscape, research outcomes will provide information to landowners to inform management decisions that promote a healthy, sustainable, and more economically viable GTR in Arkansas and across the region.
2.Methods
2.1.Study area
This study was located throughout 731 ha across 12 GTRs on the property of Five Oaks, a duck hunting lodge in Jefferson and Arkansas counties.These GTRs have been privately owned and managed over 40 years to support the wintering waterfowl population in the lower delta of the MAV, and are located within the Bayou Meto River Basin (Fig.1).Historical annual flooding of these GTRs varied only slightly, and in general flooding dates range from early November to mid-February.The management techniques have varied throughout the 12 GTRs(Table 1).
According to MAV potential natural vegetation maps (Klimas et al.,2012),tree species likely to be found at these GTRs include red oaks such as willow oak (Quercus phellos) and nuttall oak (Quercus texana), white oak species such as overcup oak (Quercus lyrata), as well as sugarberry(Celtis laevetiga), green ash (Fraxinus pennsylvanica), elm species (Ulmus sp.) and hickory species (Carya sp.).Elevations range from 51.08 to 57.87 m above mean sea level (Table 1) and soil types for this region mostly consist of alfisols, inceptisols, and entisols, and are developed from alluvial sediments which range from sandy, to loamy and clayey(United States Soil Conservation Service et al., 1982).Yearly average mean temperature within the region for 2022 was 18°C with extreme highs up to 38°C and lows down to-15°C.Total rainfall for 2022 was 119 cm and total snowfall was 5.1 cm (National Centers for Environmental Information,2022).
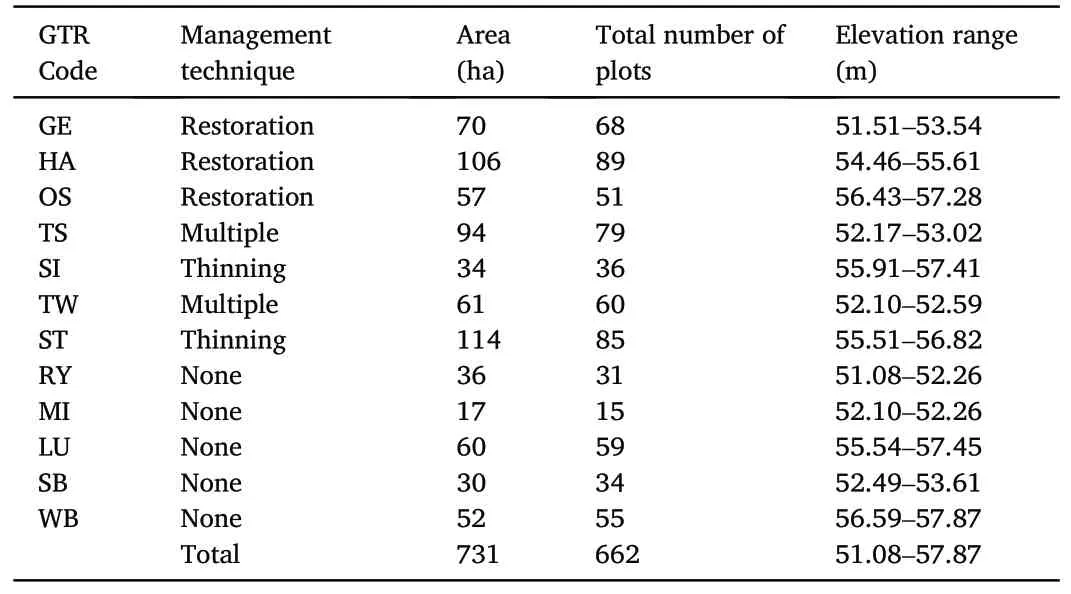
Table 1Twelve greentree reservoirs(GTRs)at Five Oaks,Humphrey,Arkansas,showing management technique, size, and the number of plots inventoried.
2.2.Sampling design and plot distribution
We used a systematic sampling approach to establish sampling plots.This sampling technique serves as the basis for conducting a high-density plot system,allowing for analysis of changes in forest communities across varying environmental conditions.A 100 m2Universal Transverse Mercator (UTM) grid system was superimposed over the 12 GTRs, and the center coordinates of the grid were used to establish plot center.Considering the GTRs sizes, the number of sampling plots distribution ranged between 15 and 89 plots.A total of 662 plots of 0.04 ha(0.1 acre)in size were sampled.Overstory data was recorded by establishing the plot center using a distance measuring tool, the Forestor DME 201(Haglof,Sweden).The Forestor DME 201 was used to measure and mark with flags,a distance of 12.4 m in each of four cardinal directions using a compass to establish a temporary plot to sample the trees inside the plot radius.
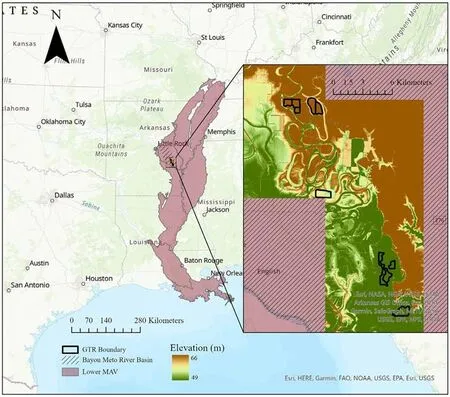
Fig.1.Study area within the Lower Mississippi Alluvial Valley and Bayou Meto River Basin.The polygons with black boundaries(figure on the right)are the 12 GTRs in the study site managed by Five Oaks across elevations (color gradient).
2.3.Data collection
We measured overstory forest structure and composition at 662 sample plots during September-November 2021 and May-August 2022,excluding plots that fell on levees and those with no overstory tree component(>12.7 cm diameter at breast height(dbh)).We recorded all overstory living and dead trees with dbh >12.7 cm (5 inches) in each plot, starting with the most northern outward tree from the plot center and working clockwise.Though we measured stems below 12.7 cm,which serves as a basis for understanding regeneration conditions,we did not include them in the analysis of overstory composition.Tree species were identified,and dbh measured with a diameter tape.Trees along the outermost edges of either plot were included in measurements if basal area was more than 50%in the plot.
We determined elevation at plot center from Light Detection and Ranging(LiDAR)raster data downloaded from USGS National Map using ArcGIS Pro version 3.0.3(Esri Inc.,2022).Soil texture classifications and soil pH values were determined from the Soil Survey geographic data(Soil Survey Staff,2023),downloaded as shapefiles,and extracted to plot coordinates.Soil texture fell into two classifications; clay and silt loam,and pH ranged from 4.8 to 7.5.Information regarding the management history for each GTR was categorized into four groups, including restoration, thinning, multiple, and none.The restoration group includes a GTR that has had renovation of hydrologic functions, such as levee management or replanting,which most commonly are the species of the red oak, such as willow oak and nuttall oak.The thinning management group includes any GTR with a history of thinning or cut treatments,which vary throughout the GTRs within this management type.However,thinning across the GTRs was done to favor desirable mast species like red oaks rather than for timber value.Gap selection was done to maintain uneven-aged stands and promote the regeneration of desirable species by providing gaps in the canopy to improve light availability.Multiple group includes GTRs with both thinning and restoration treatments,and the none group has no history of thinning or restoration treatments(Table 1).
2.4.Data analysis
All inventory data were processed and analyzed in R version 4.2.1(R Core Team,2021).We considered only live overstory trees in the analysis of forest species composition.We used descriptive statistics to summarize stem density (trees·ha-1), dbh (cm), and tree height (m) by GTRs and elevation class interval (m; 7 bins of 0.3 m).Variables were tested for normality and heterogeneity of the variance using the Shapiro-Wilk normality test (p-value >>0.05) and Levene's test (p-value >>0.05).Mixed effect modeling was used to determine if there were significantly different mean stem density,dbh and height across the 6 elevation bins.The random effect associated with mean stem density was the GTR and both dbh and height had nested random effects of the plot nested with the GTR.We calculated the importance value index (IVI) (Curtis and Mcintosh,1950;Skeen,1973)for all species across all GTRs to determine the overall dominance of each species within specific elevation bins.This was calculated using relative frequency (Eq.1), relative density (Eq.2)and relative dominance (Eq.3), and summarized by elevation bin.
2.4.1.Species diversity
Alpha diversity was examined at the plot level to index diversity within the localized overstory community.This method of calculating diversity of a single component of the forest ecosystem(i.e.overstory)is common (Ervin et al., 2006; Gracia et al., 2007; Yeom and Kim, 2011)and useful given the contribution of a variety of different overstory tree species to wildlife.Three indices were used to calculate alpha diversity;species richness,calculated as the number of different species in the plot,Shannon-Wiener diversity index (Eq.5) (Shannon and Weaver, 1949),and Pielou's evenness index (Eq.6) (Pielou, 1966).The mathematical equations for diversity and evenness are as follows:
where,S is species richness,piis the proportion made up of each species in the sample and ln is the natural log.Pielou's evenness index is derived from Shannon-Wiener diversity index(H′),the mathematical equation is:
After estimating alpha diversity, we fitted a linear regression to test the correlation with elevation.Again,normality assumptions were tested using the Shapiro-Wilk normality test (p-value >>0.05) and Levene's test(p-value >>0.05).One-way ANOVA was used to determine if there was a significant difference in the means between the categorical variables(management type and soil texture)and the three indices.
Beta diversity refers to the differences in community composition across multiple samples.To visually explore this, we used non-metric multi-dimensional scaling (NMDS) to determine the dissimilarity in the community abundance data of overstory tree species(Cram et al.,2017).NMDS is a non-parametric indirect gradient analysis that ordinates community data using rank-order rather than numeric distances.NMDS is a common ordination method used in ecological data because of its ability to handle a large ratio of species absences (0's).NMDS was calculated using function metaMDS in vegan package in R (Oksanen et al., 2022), to ordinate species abundance across all plots (n = 662),using the Bray-Curtis dissimilarity calculation,with three dimensions(k=3)and 20 random restarts.The stress value was calculated to insure the interpretabilty of the solution with guideline for an accepted stress value<0.20(Clarke,1993).The Envfit function of the vegan library in R was used to fit environmental vectors for elevation and easting to determine the strength of each variable influence on the first and second axis.The goodness of fit was also determined for soil texture classifications and management type to evaluate the relationship between these variables and the ordination analysis.
2.4.2.Species abundance
We modeled the abundance of three red oak species (nuttall oak,willow oak,and cherrybark oak) to determine the influence of environmental variables on their abundance response.These species were chosen because of their nutritional value to waterfowl and economic value to timber industries (Fredrickson, 1979).Generalized linear mixed effect models were fitted by each species.The response variable(i.e.plot level species abundance)was fitted with the Poisson distribution(Eq.7)using R package glmmTMB to account for a zero-inflated model, because the abundance of a species was zero in some plots(Brooks et al.,2017).In the model,covariates include soil texture,management technique,overstory stem density, and overstory species richness as fixed effects, and were determined at the plot level.To help detect fine-scale effects of elevation on individual species abundance, we re-grouped elevation into 14 bins,approximately at 0.5 m intervals.In the model,we included the GTR as the random effect associated with the variability in red oak species abundance.This is because the plot level species abundance within the GTR tends to be more similar than between GTRs.Overdispersion of the model was checked and calculated by dividing the residuals of the model by the degrees of freedom of the residuals.We also evaluated the variance inflation factors (VIF) to gauge potential multicollinearity of covariates in the model.All covariates with a VIF less than 5 were retained in the final model form (Eq.8).The mathematic equation for the Poisson distribution is:
where,p(x)is the probability of x the number of occurrences and λ is the expected mean value of occurrences.The generalized Poisson distribution model(Eq.8) with random effect is shown as:
where,y is the abundance of oak species(nuttall,willow,cherrybark),β0is the intercept, μkis the GTR level random effect, β1,...,βnare the coefficient to be estimated,X1,…,Xnare the covariates and εiis the error.
3.Results
3.1.Overstory summary by elevation
We measured 6,432 live overstory trees in 662 plots across 12 GTRs.Mean stem density varied across the seven elevation bins, ranging from 192.5 trees·ha-1(±115.0; standard deviation) at elevation bin 54.00-54.99 m, up to 313.5 trees·ha-1(±124.8) at elevation bin 52.99-54.00 m.Mean dbh of overstory tree species was lowest atelevation bin 54.00-54.99 m(27.61±14.50 cm)and highest at elevation bin 50.98-52.00 m (34.29 ± 14.48 cm).Across the elevation range,average tree height ranged from 15.15 ± 4.31 m to 17.51 ± 5.97 m.Elevation bin 54.00-54.99 m had the shortest mean tree height(15.15±4.31 m), while elevation bin 56.99-58.00 m had the tallest mean tree height at 18.11±5.92 m.Though some elevation bins were significantly different in stem density, dbh, and height than others, there were no consistent trends across elevation bins(Table 2).The mixed effect model indicated only 14.48% of the variability in the stand density by the random effect of GTR across the elevation bin.However, mixed effect with nested random effect explained higher variability in the average mean height(36.7%)than in the average mean dbh(20%)because of a plot nested within a GTR across the elevational bin.While, the GTR explained the variability of only 7.9% and 3.7% in the average mean height and dbh,respectively.
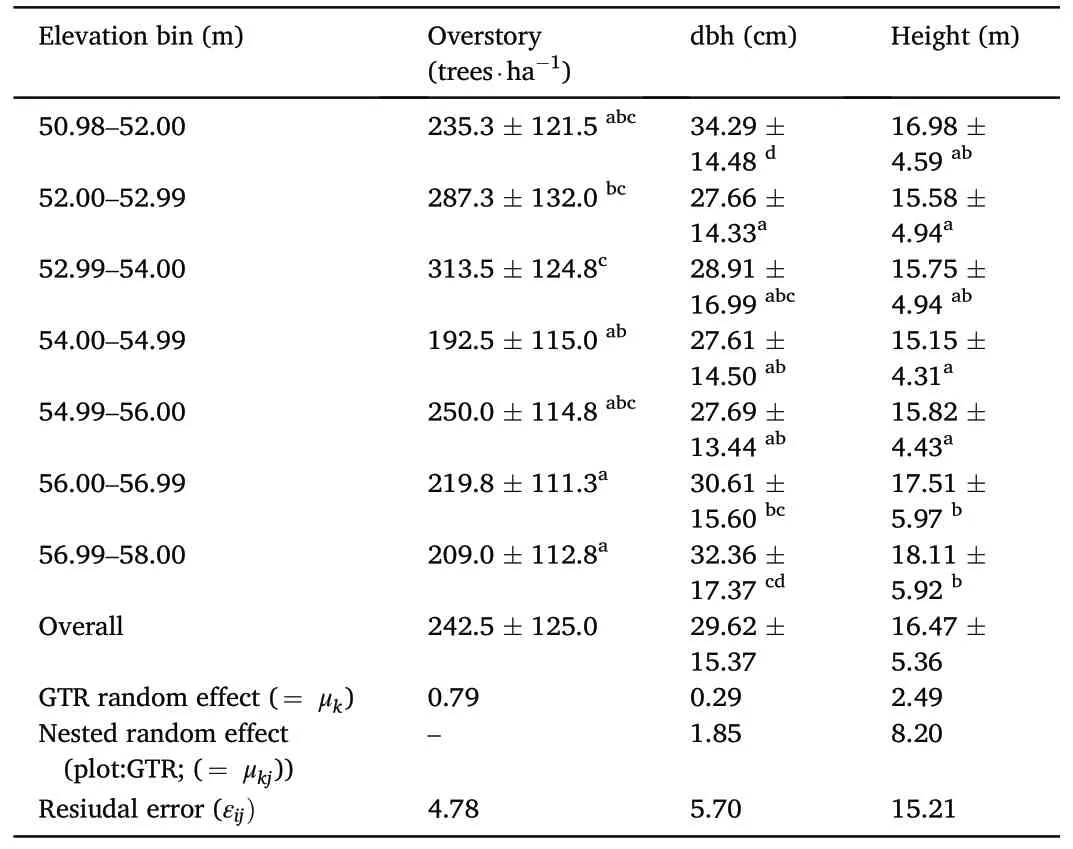
Table 2Mean(±standard deviation)of tree density,diameter at breast height(dbh),and tree height of overstory trees at seven elevation bins (n = 6,432) in greentree reservoirs in Arkansas during 2021-2022.Same letters denote no statistical difference (p-value <<0.05) in means within columns, determined using mixed effect modeling.
3.2.Importance of tree species by elevation
We identified 33 overstory tree species across 662 plots,of which 13 species were found to hold top five highest IVI across all elevations,although the IVI species order was not consistent among elevation bins(Table 3).All overstory tree species composition with IVI across the study sites is shown in Table S1.Nuttall oak had high IVI ranging from 33%to 53% throughout the lower elevation bins (50.98-52.00, 52.00-52.99,54.00-54.99 m) alongside overcup oak, which had the highest IVI ranging from 26% to 118% in the five lowest elevation bins.Shared throughout the lower elevation bins’top IVI were species such as water hickory and sugarberry.Cherrybark oak had higher IVI ranging from 21% to 29% throughout the higher elevation bins (54.99-56.00 to 56.99-58.00 m)and was accompanied by sweet gum,mockernut hickory(Carya tomentosa),and shagbark hickory(Carya ovata),all of which were found with higher IVI than oak species at the highest two elevation bins.White oak was found in the top five IVI only within the highest elevation bin, 56.99-58.00 m.Willow oak was amongst the top five species of highest IVI across all elevation bins except the highest elevation bin(56.99-58.00).This was similar for American elm(Ulmus americana)and cedar elm(Ulmus crassifolia),which were also found amongst the highest IVI values in all elevation bins except the highest(Table 3).
3.3.Alpha diversity across elevation, management types and soil texture
The variability in the Shannon-Wiener diversity index explained by elevation was very small(adjusted R2=0.010,p-value=0.005),as was also true for the variability in Pielou's evenness index explained by elevation(adjusted R2=0.040,p-value <<0.001).However,our study did not detect a significant effect between species richness and elevation(adjusted R2=0.003,p-value=0.070,Fig.2).One way ANOVA analysis indicated a significant difference between Shannon-Wiener diversity across management types (F-value = 3.620, p-value = 0.013, Fig.3).Further,the TukeyHSD multiple comparisons of means test revealed that thinning had significantly lower mean diversity that the none treatment group (p-value = 0.015).There was also a significant difference in Pielou's evenness index across management techniques(F-value=4.690,pvalue = 0.003), with a difference in means between thinning and no treatment(post hoc test,p-value=0.001).No significant difference was found between species richness and management types(F-value=1.230,p-value = 0.281).Species diversity was not significantly different between soil textures(F-value=2.060,p-value=0.150),nor did richness vary significantly between soil textures (F-value = 3.816, p-value =0.0512).However Pielou's evenness index was greater in silt loam than in clay (F-value=12.330,p-value <<0.001)(Fig.3).
3.4.Dissimilarity of overstory community abundance and environmental drivers
NMDS fit the species abundance data well, with a stress value of0.162.Correlations between NMDS axes and environmental variables,including elevation,easting and soil pH were highly significant(p-value<0.001; Table4).Goodnessof fit values indicated some variability in species abundance due to management type (R2= 0.123, p-valu<0.001) and soil texture (R2= 0.212, p-value<<0.001).Along the first e axis,the centroid value of management was positive for multiple(0.10),and none (0.27), and negative for restoration (-0.13) and thinning(-0.33),indicating that thinning may have the strongest negative effect on species abundance.Likewise,along the first axis, the centroid of soil texture, such as silt (0.2) and clay (-0.37) had both a positive and negative impact on species abundance,respectively.We observed a shift in soil substrate from silt loam to clay with a decrease in elevation(Fig.4).Species clusters reflect the distribution of overstory communities across elevation and soil characteristics.For example, overcup oak and nuttall oak are clustered among lower elevation contour lines mixed among sites of both clay and silt loam.
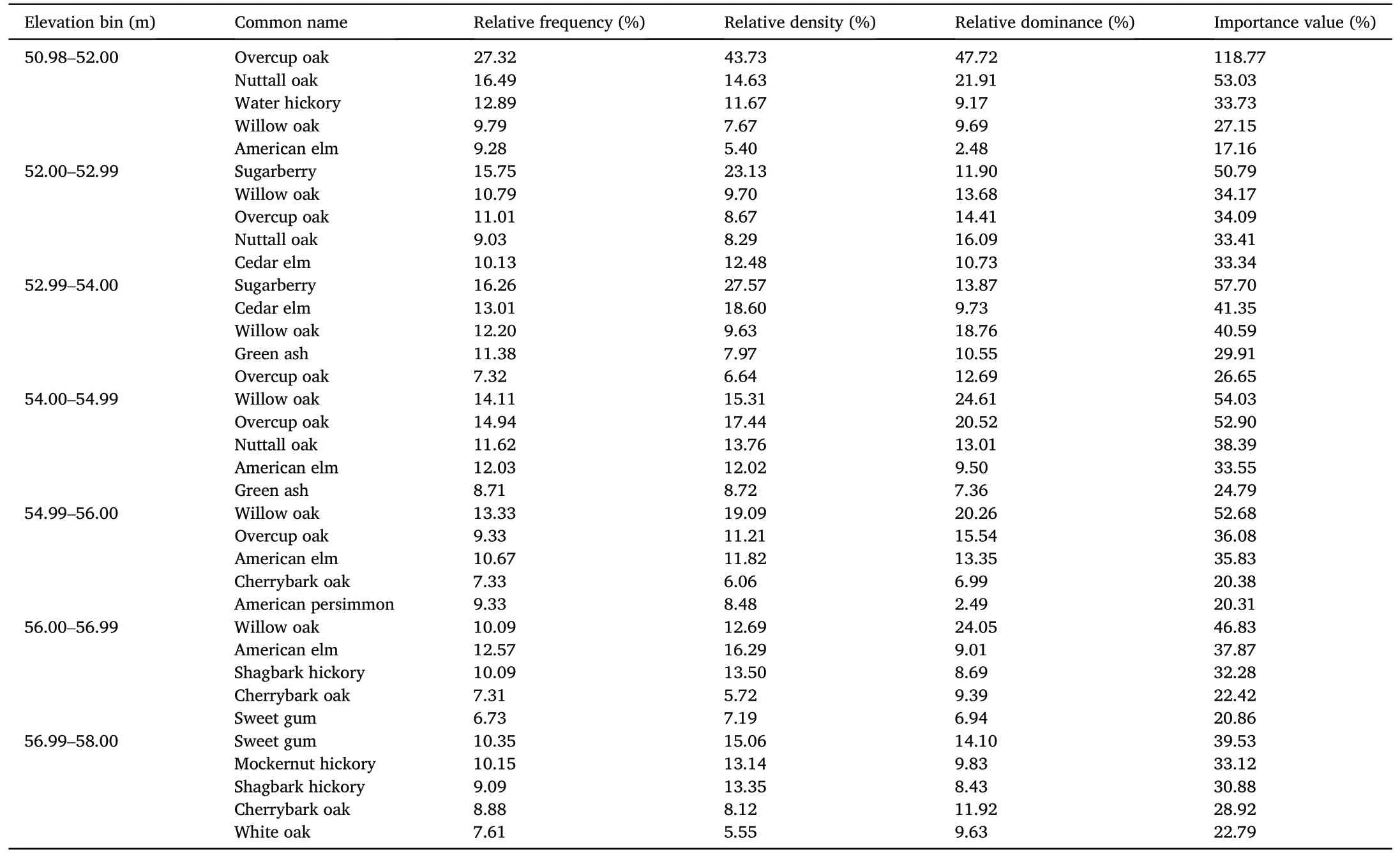
Table 3Importance values,including relative frequency,relative density,and relative dominance(n=6,432)for the top five tree species within each of seven elevation bins in GTRs in Arkansas during 2021-2022.
3.5.Factors affecting red oak species abundance
The generalized linear mixed-effects model with Possion distribution was well fitted to the abundance of three red oak species(Table 5).The model AIC and overdispersion values were 1,377.8 and 1.2 for nuttall oak,1,786 and 1.1 for willow oak,and 945 and 1.1 for cherrybark oak,respectively.The fitted explanatory variables, elevation,soil type, management type,stem density,and species richness,did not have the same significant effect on the individual oak species abundance.Much of the variability in oak species abundance was explained by the random effect associated with the GTR,which was higher for nuttall oak and cherrybark oak abundance (Table 5).The significant coefficient of elevation in the respective models had a negative effect on nuttall oak abundance(-0.16)and a positive effect on the cherrybark oak(0.31).In comparison to clay soil texture, silt loam had a significant positive affect on willow oak abundance (0.44).In comparison to no (“None”) management, the management types, multiple and thinning had no significant effect on nuttall oak and cherrybark oak,but restoration had a positive impact on willow oak abundance(Table 5).Stem density had a significant positive coefficient estimate within each species model,indicating that increases in stem density were due,at least in part,to abundance of red oak species.However,richness had a significant negative coefficient estimate across nutall and willow oak species models, indicating that as the number of species increased,the abundance of these oak species decreased.
4.Discussion
4.1.Elevation as a driver of tree species composition and diversity
Previous BHF research has been conducted to understand forest composition while assuming relatively homogenous elevation throughout (Ervin et al., 2006), or across larger elevation intervals(Meadows and Hodges, 1997; Grell et al., 2005; Rives et al., 2020).However,limited analysis has focused on the forest species composition of GTRs across fine-scale elevation variation, as this study does.The number and type of overstory tree species found were consistent with other research on BHF and GTRs in the lower MAV (Ervin et al., 2006).Our findings show that species diversity and evenness increased with elevation,resulting in higher diversity at higher elevation sites,which is consistent with research on lowland hardwood forests in central Illinois,USA (Adams and Anderson, 1980).This pattern likely is a result of unfavorable abiotic factors in lowland areas, such as anoxic soil conditions during the growing season,precluding all but those species adapted to such challenging conditions.Elevation is associated with differences in native grasses (D’Antonio et al., 2000) and non woody species, specifically ferns, herbs, and shrubs, are found to increase with elevation in lowland montane forests (Jimbo et al., 2023).However, this pattern is opposite in forests among mountainous regions where lower elevations result in higher species diversity (Zhu et al., 2019; Haq et al., 2022).These varying responses to elevation and species diversity within different ecosystems can potentially be attributed to the mid-domain effect, which is a common ecological pattern where richness increases with environmental gradients to a point and then begins to decrease(Colwell and Hurtt, 1994; Colwell and Lees, 2000).This effect is hypothesized to be the result of many factors, such as area, rainfall, temperature,and competition(Menge and Sutherland,1976;Rahbek,1995;Wang et al.,2022).

Fig.2.Alpha diversity indices for (a) Shannon-Wiener diversity, (b) Pielou's evenness index, and (c) Species richness across elevation gradients by management technique in GTRs in Arkansas during 2021-2022.Solid lines are significant (p-value <<0.05).

Fig.3.Alpha diversity by management type (a, b, c) and soil texture (d, e, f) in GTRs in Arkansas during 2021-2022.

Table 4Correlations of overstory tree species(35 species)abundance and environmental variables on the first two NMDS axes in GTRs in Arkansas during 2021-2022.

Fig.4.NMDS showing the relationship among overstory tree species along the first two axes due to environmental variables; elevation, easting, soil pH, soil texture,and management type.Axis of NMDS vectors show correlations between abundance data and environmental variables; easting, soil pH, elevation.Contour lines depict elevation (m), site colors indicate soil texture and site shapes denote management type.Species that rank significantly within the abundance matrix(p-value ≤0.001),determined using the Tau rank correlation,are labeled with first two letters of genus and first two letters of species combined:AcRu(Acer rubrum),CaAq(Carya aquatica),CaLa(Carya laciniosa),CaOv(Carya ovata), CaTo (Carya tomentosa), CeLa (Celtis laevigata), FrPe (Fraxinus pennsylvainca), LiSt (Liquidumbar styraciflua), NySy (Nyssa sylvatica), QuAl (Quercus alba), QuLy (Quercus lyrata), QuPa (Quercus pagoda), QuPh (Quercus phellos),QuSi (Quercus similis), QuTe (Quercus texana), UlAl (Ulmus alata), UlAm (Ulmus americana), and UlCr (Ulmus crassifolia).
Our analysis shows that elevation plays a role in the abundance of nuttall oak and cherrybark oak,but less so for willow oak.As expected,within the lower elevation classes,overcup oak and nuttall oak maintain higher dominance (Fredrickson, 1979; Hook, 1984; Tanner, 1986), and at least for nuttall oak, the model revealed higher abundance of this species within lower elevations of the GTRs.Found alongside nuttall oak and overcup oak within lower elevation classes were water hickory,sugarberry, green ash and elm species.These are often found as early successional species,adapted to growing in low-elevation sites with poor drainage(Hodges,1997;Rives et al.,2020).Cherrybark oak had a higher dominance and abundance in the higher elevation sites, confirmed by both NMDS analysis and our model.White oak (Quercus alba) was prevalent in higher elevation sites and had an increased dominance at the highest elevation bin.These findings are consistent with previous literature that shows these two species are typically found along the ridges of major river bottoms (Hodges, 1997).Found in higher elevation sites in community with cherrybark oak were Hickory sp., American elm, and sweet gum, which tend to be the earlier successional species in better-drained sites within higher elevations along major river bottoms(Clatterbuck and Hodges, 1988; Hodges, 1997).These results are likely driven by water availability and relatively low tolerance for waterlogged soil.Species found in our higher elevation sites tend to be less water tolerant and less drought sensitive, whereas species found in our lower elevation sites tend to be moderately water tolerant and more sensitive to drought(Kassahun and Renninger,2021).
Despite differences in elevation, willow oak was consistently dominant across all GTRs.This was slightly unexpected given willow oak is considered very water tolerant and tends to be more prominent in sloughs(McKnight et al.,1981;Hodges,1997).Discrepancy between our study and previous literature may be explained by additional effects from soil texture or management history, revealed in our willow oak abundance model.Willow oak is a common red oak species planted in BHF restoration in Arkansas, which could explain the significant correlation between our willow oak abundance model and restoration management type.Ulmus sp.seemed to be similarly unaffected by elevation within the GTRs, although specific elm species had a shift from cedar elm to American elm, as elevation increased,amongst our mid-range elevation bins.In the southeastern United States, American elm is known to be abundant in clay and silt loam soils (Bey, 1990), which may explain itswidespread abundance with increasing elevations and silt loam texture in our study.
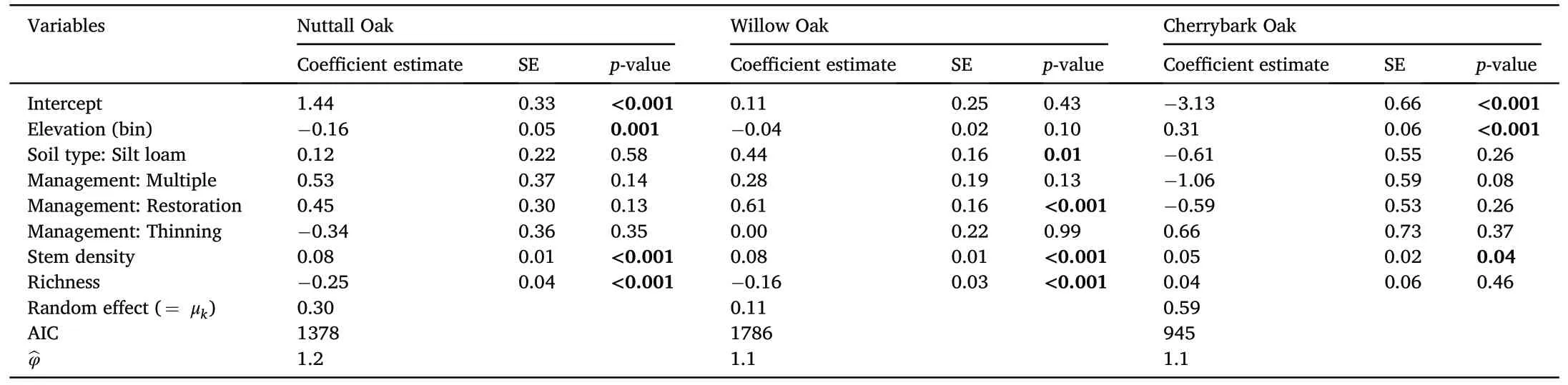
Table 5Parameter estimates of mixed-effects model fitted with Poisson distribution of abundance response of the three red oak species(nutall oak,willow oak,cherrybark oak;n = 662).Bold values represent significant coefficients within our 95% probability range.
4.2.Forest composition and soil texture
Mississippi alluvial floodplains originated from past geological events(i.e.glaciation) that created river valleys.Over time, meandering rivers have eroded and deposited the poorly consolidated sediment creating varying topography (Hodges, 1997; Kellison et al., 2019).Soils upslope from the river valley tend to be older,derived from sedimentary material,whereas soils closer to the river are younger, deposited from the river,and stratified by sediment size, transforming the soil types and textures across the topography (Hodges, 1997; Hatten et al., 2014).Finer soil textures are typically slower to drain and found at lower elevations,whereas coarser soil textures are better drained,found typically at higher elevations.Soil texture is ecologically important due to its effects on fertility and moisture availability.Specifically, soil textures have shown to contribute to variation in a forest community in southeastern longleaf pine (Pinus palustris) stands, and upland hardwood stands in the southeastern United States (Gilliam et al., 1993).This was illustrated in our study which showed correlations between soil texture and species abundance; specifically that silt loam correlated with willow oak abundance.Similar to Adams and Anderson's (1980) study of species composition within lowland hardwood forests, our results showed that the tree species with highest abundances distributed within silt loam soil types were cherrybark oak,white oak,shagbark hickory and mockernut hickory.This is likely due to these sites having increased pedogenesis,which increases the abundance of these oak and hickory species(Gardiner,2001).Species associated with clay soil types were cedar elm and sugarberry,which was similar to findings in central Illionois where Ulmus sp.and hackberry (Celtis sp.) were shown to have a high importance within wet-mesic sights (Adams and Anderson, 1980).Although neither soil texture nor soil pH explained large percentages of the variability for our ordination analysis, both were found to be significant explanatory variables.Perhaps other physical and chemical soil properties,such as porosity and organic material,could help explain more of the variability.
4.3.Management for desired forest communities
Forest management actions can result in varying levels of success in meeting objectives.Hillside forest restoration has been shown to decrease soil erosion, surface runoff, and flooding, in China (Yu et al.,2021).Restored BHFs in Kentucky, did not sufficiently replace the wildlife value of mature BHFs “due to insufficient establishment and ingrowth of heavy mast species” (Shear et al., 1996).Our findings suggest that restoration management techniques may be a valid method for increasing willow oak abundance, which was shown to be more responsive to restoration techniques in contrast to nuttall oak and cherrybark oak.Because willow oak significantly responds to restoration within GTRs, and is relatively unaffected by fine scale elevational changes,it may be desirable to land managers to promote within GTRs.Likewise, thinning and multiple management types did not have a discernible impact on the red oak species analyzed in this study, but thinning did result in a decrease in species diversity across the GTRs.In broadleaf forests found in temperate regions of Nepal,thinning has been shown to have no effect on species diversity(Webb and Gautam,2001),but resulted in a higher functional diversity in tropical forests of Borneo(Mahayani et al., 2022).This difference could potentially be accounted for by the specific target species being thinned, as well as the primary goals of forest stand management.In our GTRs, thinning is primarily conducted to decrease competition and increase the growth of dominant red oak trees and understory growth, with less emphasis on promoting overstory species diversity.
To manage GTRs towards desirable species, such as red oaks, it is important to understand the initial forest community and its natural successional pattern under certain environmental conditions, and then guide the community towards a preferred forest type.One study has shown, in the absence of management, the dominant tree species could shift from dominant to co-dominant (Cram et al., 2017).While others suggest that both restored and mature stands appear to have a succession effect from hydric to mesic species composition on managed BHFs in Kentucky,USA(Shear et al.,1996)and that deeper water can prevent the establishment of less water tolerant hardwood species(Casey and Ewel,2006).With this is mind,our findings may lead land managers towards restoration management techniques that alter the hydrology within a GTR.For example, reducing the hydroperiod and depth within GTR impoundments may be a useful method to shift forest composition away from hydric and early successional species towards red oak species.For instance,in lower elevations, discouraging the growth of earlier successional species like the water hickory-sugarberry-elm group may aid in increased dominance of nuttall oak,an economically valuable species.In higher elevations this may require methods of subduing growth of hickory-sweet gum-elm to increase red oak species like cherrybark oak.
Knowing the appropriate species to promote under specific elevation ranges and soil textures will facilitate better economic outcomes to landowners and is crucial for promoting the ecosystem services of GTRs.Furthermore, restoration efforts of BHFs on private lands continue to thrive through initiatives such as the Wetlands Reserve Easement Program, that joins private landowners with the Natural Resource Conservation Service to protect, restore and enhance wetlands.The success of such programs in the US is enhanced by increased understanding of plant communities within this ecosystem (King and Keeland, 1999) and how best to manage them.Expanding the depth of understanding of relationships between tree health and microenvironments within BHF ecosystems will increase the success of these programs by informing the planting and/or promoting of species most suited for specific microenvironments.
5.Conclusions
Our results offer important insights into management strategies and their implications within GTRs.There have been limited studies encompassing the sampling intensity achieved in our study which has allowed for a fine-scale understanding of how small elevation changes alter forest composition.Elevation played a role in forest composition in our study, particularly nuttall and cherrybark oak abundance, whereas soil texture and restoration influenced the distribution of willow oak.This study was limited in management history information and evaluating more specific management history that may provide a more specific understanding of the direct effects of management methods towards forest composition.Future research examining a detailed heath evaluation of red oak species across fine scale elevation gradients would be useful in monitoring abundance and vigor.Ultimately,recognizing how the changes in elevation and management strategies alter forest communities,particularly red oak species,provides information on how best to manage GTRs to preserve their ecosystem services and economic viability.
Authors’contribution
P.S.conceived the study design,sample collection,methodology,and data analysis.C.H.carried out data collection and analysis and wrote the manuscript.P.S.supervised the project and reviewed and edited the manuscript.K.M., and D.O.reviewed and edited the manuscript and contributed to the content.All authors checked and approved the final content of the manuscript.
Availability of data and materials
The data used for this study involves information on the private landowner's property.The study aims to continue to collect repeated measurements,and the data sharing limitation may occur if a conflict of interest exists.So, the disclosures of data supporting this study are available from the corresponding author upon reasonable request.
Declaration of conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
Financial and logistic support for this research was provided by Five Oaks Ag Research&Education Center(DS18849),University of Arkansas Division of Agriculture (PR02276), and University of Arkansas at Monticello.Funding support was provided by U.S.Department of Agriculture,National Institute of Food and Agriculture, McIntire-Stennis Capacity Grant(2729 YR21-25 and ARK02594).We would like to thank R.Askren for his assistance on plot distribution and input on analysis.We are also grateful to many field technicians who helped collect and compile field data.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://do i.org/10.1016/j.fecs.2023.100141.

- Forest Ecosystems的其它文章
- Growth phenology adjusts to seasonal changes in water availability in coexisting evergreen and deciduous mediterranean oaks
- Assessing spatiotemporal variations of forest carbon density using bi-temporal discrete aerial laser scanning data in Chinese boreal forests
- Stabilizing forest productivity and resilience at multiple scales
- Legacy effects of historical woodland changes on contemporary plant functional composition
- Habitat heterogeneity and biotic interactions mediate climate influences on seedling survival in a temperate forest
- Trees species’ dispersal mode and habitat heterogeneity shape negative density dependence in a temperate forest