
Legacy effects of historical woodland changes on contemporary plant functional composition
2023-11-15 07:56JiZhongWanChunJingWang
Forest Ecosystems 2023年5期
Ji-Zhong Wan, Chun-Jing Wang
a State Key Laboratory of Plateau Ecology and Agriculture, Qinghai University, Xining 810016, China
b Departamento de Ecología, Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile, CP 8331150, Santiago, Chile
c Centro de Cambio Global UC, Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile, CP 8331150, Santiago, Chile
Keywords:Biome Community weighted means Global scale Historical woodland Land cover Plant functional trait
ABSTRACT Considering the legacy of plant functional composition can help assess ecosystem functions and ecosystem services across different spatial scales under land cover changes.Woody plants likely respond to natural and anthropogenic perturbations due to historical events (e.g., agricultural development), thus contemporary plant functional composition may be explained by historical woodland change,a type of land cover change.We propose that historical woodland changes may have legacy effects on contemporary plant functional composition.Here,we used partial least squares regression and linear mixed model analyses to test this assumption by coupling data on community weighted means (CWM) and community weighted variance (CWV) of vegetation plots and calculating the time of woodland existence across different periods from AD 0 to 2017.We found that the legacy effects of historical land cover changes on CWM and CWV during the existence time of woodland, particularly from AD 0 to 900,were drivers of contemporary plant functional composition at large spatial scales.Furthermore,historical woodland changes can affect contemporary plant functional composition,depending on the biome type.Particularly, the CWM of plant height, seed mass, and seed length showed the strongest correlations with woodland changes from AD 1910 to 2010 in tropics with year-round rain, and the CWM of leaf traits correlated with woodland changes from AD 0 to 1700 in tropics with summer rain.Our study provides the effective evidence on the legacy of historical woodland changes and the effects on contemporary plant functional composition,which is crucial with respect to effective management of plant diversity and assessing ecosystem functions and services from local to global scales over time.
1.Introduction
Plant functional composition, defined by both community-weighted mean (CWM) and community weighted variance (CWV) traits, is closely linked to variations in ecosystem functions and services, particularly with respect to functional traits (Díaz and Cabido, 2001; Lavorel et al.,2011;He et al.,2019).CWM and CWV are valuable tools describing community composition with respect to one given trait,indicating intraand interspecific trait variation,respectively,regarding the maintenance of plant species diversity (Lisner et al., 2021).These plant functional traits are critical for understanding ecological communities because of their effects on community assembly processes, potential species interactions, and other ecosystem functions (Díaz and Cabido, 2001; Lavorel et al., 2011; Adler et al., 2013; Kraft et al., 2015; Kunstler et al.,2016; He et al., 2019).To improve the effectiveness of ecosystem assessment and management, the spatial distributions of contemporary plant functional composition have been widely predicted at regional and global scales based on trait-environment relationships(Jetz et al.,2016;Blonder et al.,2018;Boonman et al.,2020).Respective studies(e.g.,Jetz et al., 2016; Boonman et al., 2020) used contemporary environmental variables (e.g., climate and soil) to assess contemporary plant trait distributions across different spatial scales; however, increasing evidence(e.g.,Blonder et al.,2018;Armstrong et al.,2021)suggests that historical environmental changes exert important direct and indirect effects on contemporary species composition and functional composition.
Land use and cover legacies exert long-term effects on plant community composition and ecosystem functions at local scales (Díaz et al.,2007; Chillo et al., 2018; Armstrong et al., 2021; Jiménez-Franco et al.,2022).In particular, woodland changes may be important drivers of community composition and function in the Anthropocene (Armesto et al., 2010; Wolkovich et al., 2014; Turvey and Crees, 2019; Swanson et al.,2021).Changes in woodland structure may affect biodiversity,and the extent of forest fragmentation and disturbance determine woodland change (Nordén et al., 2014; Liu et al., 2019; Swanson et al., 2021).Woodland loss and fragmentation have decreased forest availability within suitable climatic ranges, and these factors are jointly related to plant species loss (Cattarino et al., 2014; Nordén et al., 2014; Taubert et al., 2018).Woodland expansion and retraction can regulate biodiversity and ecosystem services to humans(Burton et al.,2018;Liu et al.,2019).Therefore, assessing legacy effects of past woodland changes on contemporary plant functional composition may help improve our understanding of the contribution of current woodland management to future ecosystem functions and services.
To our knowledge, most studies (e.g., Mayfield et al., 2010; Ford et al.,2018;Zambrano et al.,2019)have focused on the legacy effects of woodland change in contemporary plant functional composition based on current data.However, historical environmental changes may strongly affect the functional composition of contemporary plant communities (Blonder et al., 2018; Chillo et al., 2018; Armstrong et al.,2021).Blonder et al.(2018) showed that the climate prevailing tens of thousands of years ago influenced contemporary plant functional composition via slow assemblage dynamics.Slower evolutionary and biogeographic processes (e.g., speciation and changes in distribution ranges)typically predominate at regional and continental scales,whereas at local scales, ecological processes (e.g., dispersal, abiotic and biotic selection, and random drift) are more important regarding immigration and extinction (Ron et al., 2018; Fajardo et al., 2019; Mucina, 2019).Woody plants are comparably long-lived, thus adult individuals may be present even if the conditions are no longer suitable for reproduction(Samish, 1954).If the rates of woodland-driven immigration and/or exclusion of species are slow, the contemporary functional composition may be explained by historical woodland changes.A time delay would thus imply the amount of time that elapses before the state returns to an equilibrium for woody plants (Díaz et al., 1998; Gaüzˋere et al., 2020).Therefore, woody plant communities likely respond to natural and anthropogenic perturbations with a considerable time delay.Considering delayed effects allow for a more reliable estimation of legacy effects of historical woodland changes on contemporary plant functional composition.
We propose hypothetical scenarios for the relationship between historical woodland changes and contemporary plant functional composition, as illustrated in Fig.1 for legacy effects of historical woodland changes on contemporary plant functional composition.The time of woodland existence per period may have different effects on the functional composition of contemporary plants.The provision of various ecosystem services was assumed to vary across different periods from AD 0 to 2017; thus, ecosystem management strategies and land use can modify ecosystem functions, depending on the duration of woodland existence per period.Forest management can alter forest functionally both increasing and reducing the number of functions.Furthermore,the effects of ecosystem management may differ between biome types.
Specifically,we propose the following two hypotheses:
(1) Before 1700, nearly half of the terrestrial biosphere was wild,without human settlements or substantial land exploitation(Ellis et al., 2010).Between 1700 and 2000, the terrestrial biosphere made the critical transition from mostly wild to mostly anthropogenic, passing the 50% mark early in the 20th century (Ellis et al., 2010; Goldewijk et al., 2017).Hence, the trends of woodland changes may be different between the periods from AD 0 to 1700 and from 1710 to 2017.Here,we propose the first hypothesis(H1):the effects of woodland changes from AD 0 to 1700 on contemporary plant functional composition are different from those from 1710 to 2017.
(2) A biome is a biogeographical unit consisting of a biological community that has formed in response to environmental change(Mucina,2019).Plants belonging to different biomes have varied abilities to tolerate anthropogenic transformation of the biomes at a global scale (Ellis, 2011; Pletcher et al., 2022).Hence, it is possible that biome-level relationships exist between land cover changes and plant diversity.Here, we propose the second hypothesis (H2): the effects of historical woodland changes on contemporary plant functional composition vary depending on biome changes.
To test these hypotheses, we used a partial least squares (PLS)regression coupled with linear mixed model(LMM)fitting to explore the relationships between historical woodland changes across seven periods spanning over 2000 years and contemporary plant functional composition across different biomes on a global scale.
2.Materials and methods
2.1.Data on contemporary plant functional composition
Analyses were executed based on 18 plant functional traits,which are most closely associated with ecosystem functions and services,as follows:(a) leaf area (mm2); (b) stem specific density (g·cm-3); (c) specific leaf area(m2·kg-1);(d)leaf carbon concentration(mg·g-1);(e)leaf nitrogen(N) concentration (mg·g-1); (f) leaf phosphorus (P) concentration(mg·g-1);(g)mean plant height(m);(h)seed mass(mg);(i)seed length(mm);(j)leaf dry matter content(g·g-1);(k)leaf N per area(g·m-2);(l)leaf N:P ratio; (m) leaf δ15N (per million); (n) seed number per reproductive unit;(o)leaf fresh mass(g);(p)stem conduit density(per mm2);(q)dispersal unit length(mm);and(r)wood vessel element length(μm);all concentrations are based on dry weight(Sabatini et al.,2021).
We retrieved data on plant functional composition from the sPlot database v2.1 (Sabatini et al., 2021).This database contains 1,121,244 unique vegetation plots and 23,586,216 species records,originating from 110 different vegetation-plot datasets of regional, national, or continental extent, some of which stemmed from regional or continental initiatives.The metadata of each individual vegetation-plot dataset stored in sPlot are managed through the Global Index of Vegetation-Plot Databases(GIVD;Dengler et al.,2011).Most of the data in sPlot database v2.1 refer to natural and seminatural vegetation,while vegetation shaped by intensive and repeated human interference, such as cropland or ruderal communities, is hardly represented.Establishment time is from 1900 to 2015.We log-e-transformed the functional trait data before calculating CWM and CWV (Sabatini et al., 2021).CWM and CWV were calculated for every plant functional trait j and every vegetation plot k as follows:
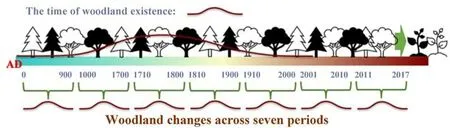
Fig.1.Hypothetical scenarios for the relationship between historical woodland changes(i.e.,the time of woodland existence) and contemporary plant functional composition across seven periods,i.e., from AD 0 to 900 (every 100 years), 1000 to 1700 (every 100 years), 1710 to 1800 (every 10 years), 1810 to 1900(every 10 years),1910 to 2000(every 10 years),2001 to 2010 (every year), and 2011 to 2017 (every year).In the case of the figure,the legacy effects of historical land cover changes on plant species diversity during existence of woodland from AD 0 to 2000 were the drivers of contemporary plant functional composition.
where nkis the number of species with trait information in vegetation plot k, pi,kis the relative abundance of species i in vegetation plot k calculated as the species'fraction in cover or abundance of total cover or abundance,and ti,jis the mean value of species i for trait j.
Data on CWM and CWV result in values by plot and trait were obtained for 90,923 unique vegetation plots from 105 constitutive datasets belonging to 10 biome types,including alpine,temperate,midlatitudes,subtropics with winter rain, dry tropics and subtropics, tropical with summer rain, tropics with year-round rain, subtropics with year-round rain, dry midlatitudes, boreal, and polar and subpolar zones (Sabatini et al., 2021).Here, we did not install only in forest areas because we intended to explore the effects of historical woodland changes including removed woodland areas on current CWM and CWV.The functional trait information was available for 21,854 species across all the plots for quantifying CWM and CWV of all 18 functional traits.The average trait coverage is 87%, with 74,151 plots having functional trait information for species cumulatively accounting for more than 80%of relative cover.The 68,041 plots have functional trait information for 80%or more of the species occurring in that plot.The distribution map of study plots was shown in Fig.2.
2.2.Assessing woodland changes
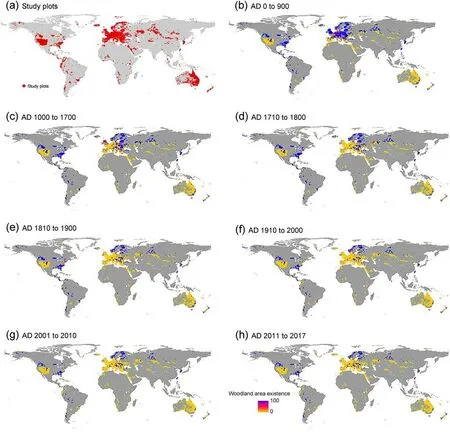
Fig.2.Distribution patterns of historical woodland changes based on the study plots (a) from AD 0 to 2017.Woodland change represents the time of woodland existence over the entire respective period,i.e.,from AD 0 to 900(every 100 years;b),1000 to 1700(every 100 years;c),1710 to 1800(every 10 years;d),1810 to 1900 (every 10 years; e), 1910 to 2000 (every 10 years; f), 2001 to 2010 (every year; g), and 2011 to 2017 (every year; h).Coloration from dark yellow to blue represented longer time of woodland existence over the entire respective period.(For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
The sPlot and HYDE datasets were merged in our study.We retrieved woodland maps at 5.0 arc-minutes resolution across different periods from the Historical Database of the Global Environment (HYDE; https:://themasites.pbl.nl/tridion/en/themasites/hyde/download/index-2.ht ml).HYDE is an internally consistent combination of historical population estimates and allocation algorithms with time-dependent weighting maps for land us.HYDE classified the areas into the following land covers: 1) cropland, 2) herbaceous cover, 3) tree cover, 4) shrubland cover, 5) lichens and mosses sparse vegetation, 6) urban areas, 7) bare areas, 8) water bodies and 9) permanent snow and ice.This database reveals woodland distribution during the last few thousand years globally at a spatial scale with a 5.0 arc-minutes grid resolution (Goldewijk et al.,2017).Here,we used different periods from AD 0-2017 to quantify the woodland changes.Based on the data availability of the HYDE, we used the duration of woodland area existence to assess historical woodland changes at a long temporal scale based on seven periods,i.e.,from AD 0-900 (every 100 years), 1000 to 1700 (every 100 years), 1710 to 1800 (every 10 years), 1810 to 1900 (every 10 years), 1910 to 2000(every 10 years), 2001 to 2010 (every year), and 2011 to 2017 (every year)as shown in Figs.1 and 2.
These seven periods were based on the data availability of HYDE datasets and human activities.There are the relatively large uncertainties for assessing woodland changes from AD 0 to 1700 (Goldewijk et al.,2017).Hence,we used two periods with 100 intervals from AD 0 to 1700.From AD 1710 to 2000, there are increasing human activities, e.g., the Industrial Revolution, and the types of human activities were changed,e.g.,the trends of woody trade development and cropland expansion.The time interval should be every 10 years from AD 1710 to 2000.The resulting demographic growth rates seem plausible,and thus the data are acceptable for the purpose of this study.Based on the time intervals of HYDE dataset, we used a larger interval for the older than nearer time periods.Hence, we used one year interval from AD 2001 to 2017 with two periods.We used prospects for the post-1950 period,but before 1950 there is a strong dependency on a few historical population data sources.Some regions and time periods are not undisputed and probably on the low side of estimates.Therefore, especially deeper into the past(pre-1500 CE), the numbers must be treated with caution (Goldewijk et al., 2017).The ranges of time length were quantified based on the percentage (%) of woodland existence time relative to the complete respective time period.Thus,the occupation percentage(%)of woodland area existence was calculated for the total time span per period.The average detailed percentages (%) were following across all the study plots:1)the period from AD 0 to 900:51.0±0.2;2)the period from AD 1000 to 1900:39.3±0.2; 3) the period from AD 1910 to 2000:28.8±0.1;4)the period from AD 2001 to 2010:28.6±0.1;5)the period from AD 2011 to 2017:28.7±0.1(Fig.S1).
2.3.Analyses
Values for all 18 functional traits were projected using principal component analyses (PCA) based on CWM and CWV (Dolédec et al.,2000).There is a specific value of CWM or CWV for each plot(Sabatini et al., 2021).We extracted the loadings of the respective first principal components(PCs)explaining more than 60%of the variance in CWM and CWV, respectively, which were significant according to the number of axes to be estimated using a sequential Bonferroni procedure (Dolédec et al.,2000).The first three and five PCs can capture most of the variation for CWM and CWV and the rest can be discarded without losing much information based on 60%threshold,respectively.We mapped the plant functional trait economics spectrum based on the first three CWM-PCs(i.e., three dimensions) and the first five CWV-PCs (i.e., five dimensions),which represent the most of 18 plant functional traits.
First,we used PLS regression to determine the best predictors of plant functional composition in independent analyses (Carrascal et al., 2009;Blonder et al., 2018).We conducted PLS regressions separately for the first three CWM-PCs and the first five CWV-PCs.The first three CWM-PCs and the first five CWV-PCs were used as response variables and historical woodland changes(i.e.,the ranges of time length per period)were used as predictors.The predictors are organized in a matrix where the rows are the sPlots and the columns are the historical woodland changes.The PLS approach accounts for statistical non-independence of large numbers of predictors by finding the rotation of the predictor matrix that best overlaps with the response vector, and it identifies the latent factors(components) that correspond to these rotations (Carrascal et al., 2009;Blonder et al., 2018).The PLS components describe the independent contribution of each predictor variable to the response variable and are ordered by their explanatory capacity, such that the first component(PLS1), by definition, explains the most variation in the data (Blonder et al., 2018).Thus, the approach can identify independent effects of multiple correlated predictors (i.e., separating the effects of historical woodland changes, even if they are sometimes correlated with each other).The first PLS component should explain >60%of the variation in the data, indicating good prediction performance for plant functional composition(Carrascal et al.,2009;Blonder et al.,2018).PLS regression was carried out using the package “pls” in R environment (https://www.r-project.org/).
Then, we used the loading coefficients of PLS to explain the contributions of woodland changes to CWM-PCs and CWV-PCs with the most variance in each model,based on different periods,i.e.,from AD 0-900(every 100 years),1000 to 1700(every 100 years),1710 to 1800(every 10 years),1810 to 1900(every 10 years),1910 to 2000(every 10 years),2001 to 2010 (every year), and 2011 to 2017 (every year) (Carrascal et al.,2009;Blonder et al.,2018).
Finally, we used a LMM to explore the relationships between historical woodland changes and CWM-PCs and CWV-PCs using the package“rdacca.hp”(Lai et al.,2022)in R environment(https://www.r-project.o rg/).Here, we used historical woodland changes as fixed factors and biome type as a random factor.The first three CWM-PCs and the first five CWV-PCs were used as response variables.The predictors are organized in a matrix where the rows are the sPlots and the columns are the historical woodland changes.We compared the results of the marginal r2and conditional r2to test whether biome type affected the relationship between historical woodland changes and CWM-PCs and CWV-PCs (Lai et al., 2022).We then used ordinary least squares regression to explore the relationships between woodland changes based on different periods individually, with three CWM-PCs and five CWV-PCs.All data analyses were conducted in R environment(https://www.r-project.org/)and JMP Software(https://www.jmp.com/en_gb/software.html).
3.Results
Based on the PCA results,the first three CWM-PCs and five CWV-PCs could explain 62.226%and 60.166%of the variance in CWM and CWV,respectively.CWM-PC1 was closely related to plant height, seed mass,and seed length; CWM-PC2 was closely related to specific leaf area and leaf N content per leaf dry mass; and CWM-PC3 was closely related to wood vessel element length and leaf P content per leaf dry mass with the positive effects (Table S1).CWV-PC1 was positively correlated with dispersal unit length;CWV-PC2 was negatively correlated with seed dry mass, seed length, and dispersal unit length; CWV-PC3 was negatively correlated with specific leaf area and leaf N content per leaf dry mass;CWV-PC4 was negatively correlated with leaf area and leaf fresh mass;and CWV-PC5 was positively correlated with leaf dry mass per leaf fresh mass and wood vessel element length(Fig.3 and Table S1).
PLS1 explained 81.818% of CWM-PC1, 82.030% of CWM-PC2, and 83.752% of CWM-PC3, and 83.594% of CWV-PC1, 83.852% of CWVPC2, 65.229% of CWV-PC3, 19.718% of CWV-PC4, and 82.034% of CWV-PC5.PLS1 explained >70% of the variation in all three CWM-PCs(i.e., CWM-PC1, CWM-PC2, and CWM-PC3) and CWV-PCs (CWV-PC1,CWV-PC2, and CWV-PC5).PLS1 had the strongest effect on CWM-PCs and CWV-PCs for woodland changes from AD 0 to 900 ranging from 0.1011 to 0.4304 in absolute standard deviations based on the loading coefficients of PLS(Fig.4).Overall,the loading coefficient results of PLS showed that the contribution of woodland changes (i.e., the time of woodland existence) to CWM-PCs and CWV-PCs occurred from AD 0 to 1700 (>0.1 in absolute standard deviations for CWM-PC1, CWM-PC2,CWM-PC3, CWV-PC1, CWV-PC2, and CWV-PC5, respectively) at global scales (Fig.4).However, the effects of historical woodland changes on contemporary plant functional composition may differ across the seven historical periods(Fig.4).PLS1 had the strongest effect on CWM-PC1 for woodland changes from AD 0-900,while the effects on CWM-PC1 were the weakest from AD 1000 to 1700(Fig.4).
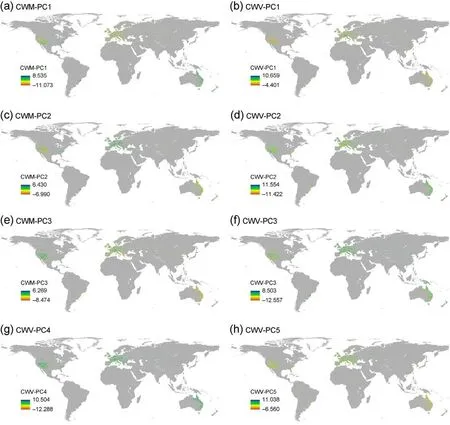
Fig.3.Distribution patterns of the first three CWM-PCs and the first five CWV-PCs based on the study plots.Coloration from dark yellow to dark blue represented higher values of PCs for CWM and CWV.(For interpretation of the references to colour in this figure legend,the reader is referred to the Web version of this article.)
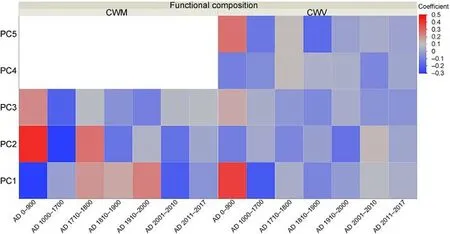
Fig.4.Loading coefficients of PLS to explain the contributions of woodland changes in different periods.Coloration from grey to red and to blue represented high loading coefficients of PLS to explain the contributions of woodland changes.Blue indicates negative effects, and red indicates positive effects.(For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
Based on the LMM, conditional r2values were larger for CWM-PCs than for CWV-PCs (Fig.5).Conditional r2values were relatively high,whereas marginal r2values were relatively low for the relationships of CWM-PCs and five CWV-PCs with woodland changes (i.e., the time of woodland existence), indicating that the biome type contributed significantly to such relationships (Fig.5).In particular, the strongest relationships were found between CWM-PC1 and woodland changes from AD 1910 to 2010 in the tropics with year-round rain (R2= 0.118; P <0.001), and between CWM-PC2 and woodland changes from AD 0 to 1900 in the tropics with summer rain(R2=0.116;P <0.001 for AD 0 to 900; R2= 0.107; P <0.001 for AD 1000-1700; Fig.6 and Table S2).Furthermore,the changing trends(i.e.,positive or negative)of CWM-PCs and CWV-PCs along the gradient of woodland changes could depend on the biome types (Fig.6 and Table S2).The strongest relationships occurred between historical woodland changes from AD 1000 to 2017 and CWV-PC1 in the boreal zone(positive;R2=0.100;P <0.001),and between CWV-PC2 and woodland changes from AD 0 to 1900 in subtropical areas with year-round rains (negative; R2= 0.100; P <0.001;Fig.6 and Table S2).
4.Discussion
4.1.Legacy effects of historical woodland changes on plant functional composition
We identified temporally variable effects of woodland changes on contemporary plant functional composition at a global scale based on the time of woodland existence across different periods.We proposed that the legacy effects of historical land cover changes on plant functional composition during the existence of woodlands from AD 0 to 1700,particularly from AD 0 to 900, were the drivers of contemporary plant functional composition (i.e., CWM-PCs and CWV-PCs).Woodland changes can lead to variation in habitat availability for plant species(Goldewijk et al.,2017;Machado et al.,2020).A given land cover change involves habitat availability moving from forest to grasslands(Goldewijk et al., 2017).The existence of redundancy in the distributions of plant functional traits within local communities has been associated with an increase in the resilience of ecosystems to habitat changes due to species with similar plant functional traits coexisting(Pillar et al.,2013;Cadotte et al., 2015; Biggs et al.,2020).Desirable conditions can be maintained through the capacity of a system to respond to habitat changes to retain fundamental functionality (Wohl et al., 2004; Pillar et al., 2013; Biggs et al.,2020).The similar functional traits of plant species(i.e.,density of woody plants) may disappear with land cover changes from forest to crops (Cadotte and Fukami, 2005; Pillar et al., 2013).The ecosystem services(i.e., woody production) may be lost(Zalesny et al.,2012).
4.2.Effects of woodland changes on plant functional composition depending on trait types
We found that a contribution of woodland changes to different trait types, i.e., CWM-PCs (i.e., CWM-PC1, CWM-PC2, and CWM-PC3) and CWV-PCs (i.e., CWV-PC1, CWV-PC2, and CWV- PC5) occurred from AD 0 to 1700 (>0.1 in absolute standard deviations for CWM-PC1, CWMPC2,CWM-PC3,CWV-PC1,CWV-PC2,and CWV-PC5).The conditional r2values were larger for intra-than for interspecific variation in plant functional traits,indicating that woodland changes had larger effects on intra- and interspecific variations in plant functional traits (Lai et al.,2022).Although CWV-PC1 (i.e., dispersal unit length), CWV-PC2 (i.e.,seed dry mass,seed length,and dispersal unit length),and CWV-PC5(i.e.,leaf dry mass per leaf fresh mass and wood vessel element length)can be explained by historical woodland changes, the respective prediction power may be weak.Here, CWM-PCs and CWV-PCs could explain plant functional composition based on different trait types.Thus, local experimental methods should be used to assess the influence of woodland changes on plant functional composition depending on trait types.
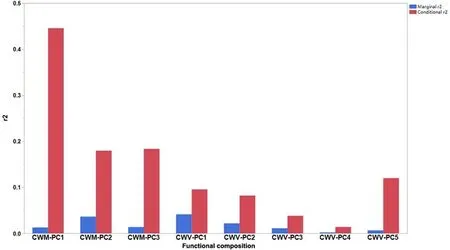
Fig.5.Results of conditional r2 and marginal r2 for the relationship between historical woodland changes and contemporary plant functional composition based on LMM.Red and blue bar graphs represented conditional r2 and marginal r2,respectively.(For interpretation of the references to colour in this figure legend,the reader is referred to the Web version of this article.)

Fig.6.Relationships(R2 and slope)between historical woodland changes and contemporary plant functional composition across different biomes based on ordinary least squares regression.Slope represented the slopes of regression equations regarding the relationships between woodland changes based on different periods individually, with three CWM-PCs and five CWV-PCs.The slope values (i.e., - or +) represented the negative or positive relationships.All the relationships were significant(P <0.05)in the figure.The relationships between historical woodland changes and contemporary plant functional composition were not shown for alpine biome due to extremely low values.
Historical woodland changes from AD 0 to 1700 can affect plant functional composition, which are closely associated with leaf traits.Specific leaf area and leaf N content per leaf dry mass are related to the potential relative growth rate or mass-based maximum photosynthetic rate (Wilson et al., 1999; Liang et al., 2020).Current global primary productivity may thus be predicted by historical woodland changes from AD 0 to 1700 given the roles of leaf traits in production (Wilson et al.,1999; Luo et al., 2004).Closed-canopy woodlands favor shade-tolerant species, and woodland changes can affect photosynthetic rates and leaf traits in multiple layers(e.g.,canopy,understory,shrub layers,and forest floor; Canham, 1988; Power et al., 2021).Changes in vegetation cover can thus shape contemporary plant functional composition(e.g.,leaf and seed traits),with a gradient of vegetation cover and a time delay.However, the predictions of historical woodland changes on seed traits are relatively weaker than those on leaf traits based on CWV-PCs.There is a strong and consistent pattern that species with dormant seeds have reduced spatial dispersal(Chen et al.,2019,2020).Plant seeds have their own dormancy period, which is closely related to seed traits affecting spatial dispersal, i.e., seed dry mass, seed length, and dispersal unit length.The time cycle of seed dormancy is typically short(Harper et al.,1970;Chen et al.,2019,2020).Hence,a long-time woodland change may have small effects on seed traits.These findings support the position that trait types can regulate effects of woodland changes on plant functional composition (e.g., between the CWV of leaf and seed traits).Hence,legacy effects of woodland changes on plant functional composition can depend on trait types.
4.3.Effects of woodland changes on plant functional composition depending on biome types
Our results showed that the impact of historical woodland changes on contemporary plant functional composition varies across biomes.In the 1900s,tropical forests were the primary source of new agricultural land and forest resources (Swamy et al., 2018; Edwards et al., 2019; Senior et al., 2019); the increasing presence of mechanized agriculture in the Brazilian Amazon serves as a prime example of this(Morton et al.,2006;Matricardi et al.,2020).Carbon fluxes from deforestation are a function of the area of forest loss and are related to forest disturbances such as fire and logging, variations in forest biomass across the basin, and land use change or abandonment after forest clearing(Morton et al.,2006;Keith et al., 2009; Swamy et al., 2018).The functions are closely related to plant functional composition (Cadotte et al., 2015).Tropical forest deforestation may lead to changes in plant height, seed traits (i.e., seed mass and seed length), and leaf traits (specific leaf area, leaf N content per leaf dry mass, and leaf P content per leaf dry mass, i.e., CWM-PCs).Accordingly, our results showed the strongest relationships of CWM-PC1 with woodland changes from AD 1910 to 2010 in tropics with year-round rain.
Historical woodland changes affected vegetation cover in the boreal zone and subtropical areas with year-round rain (Staver et al., 2011;Armstrong et al.,2021).For example,current valuable deciduous forest and wetland habitats were mostly grazed in grassland biomes after AD 1900 (Armstrong et al., 2021).However, the relationships between historical woodland changes and seed traits were the opposite: seed trait diversity was strongly correlated with vegetation height and forest cover,and differences in vegetation height and forest cover between the boreal zone and the subtropics with year-round rain were observed(Klimeˇsová et al., 2016; Chen et al., 2019, 2020).Furthermore, seed and leaf traits may have conferred resilience in species’ responses to changing environmental conditions in the boreal zone from AD 1000 to 2017 and in the subtropics with year-round rain from AD 0-1900.To sum up,the legacy effects of woodland changes on contemporary plant functional composition may depend on biome changes.
4.4.Implications for forest management
Our results on legacy effects of historical woodland changes on plant functional composition can contribute to effective forest management.The main objective of forest management is to maintain ecosystem functions and services, which can be measured with plant functional composition.Human land-use legacy effects are potentially important drivers of the structure and function of contemporary plant communities(Laliberté et al., 2010; Carreño-Rocabado et al., 2016; Reinhart et al.,2022).Extensive human activities have strong negative effects on ecosystem functions and services through threats to plant species diversity and functional traits (Beckmann et al., 2019; Carlucci et al.,2020).Ongoing land-use changes resulting from urbanization and agricultural development are considered major threats to biodiversity and tend to favor human production needs(Kremen and Merenlender,2018;Beckmann et al.,2019;Marques et al.,2019).Changes in plant functional composition can reflect ecosystem functions and services(e.g.,forest CO2sequestration, forest growth, and wood use interactions) under forest management.During the Industrial Revolution(with the year 1700 as the timeline of demarcation), solid wood products and wood energy (i.e.,fuel) could be produced by forest resources, and effective management should be conducted to maintain ecosystem functions and services(Demirbas¸,2001;Oliver et al.,2014).Wood harvested to create open and savanna structures can be used for construction products and wood energy (Oliver et al., 2014).Thus, historical woodland changes closely related to wood harvesting can lead to the variation in plant functional composition.
Legacy effects of historical woodland changes on plant functional composition may negatively affect forest management effectiveness for a long time.For example,indigenous land management practices reduced variations in plant functional composition,and the effects persisted more than 100 years after management in British Columbia, Canada (Armstrong et al.,2021).Thus,plant functional composition likely responds to woodland changes caused by natural and anthropogenic perturbations with a considerable delay in time from AD 0 to 1900, as shown in our results.Natural and anthropogenic forests have different functions for biodiversity and ecosystems(Matricardi et al.,2020;Lesiv et al.,2022).Natural forests may have more complex species and functional compositions than anthropogenic forests, and vice versa (van der Plas, 2019;Matricardi et al., 2020).Here, we need to consider the exception of single-species forest or some mixed forests.In monospecific forests, the new species can have alternative traits, and the coexisting species can have complementary yet rather similar traits in mixed forests (van der Plas,2019;Cassart et al.,2020).Hence,we should apply plant functional composition as an effective ecological indicator for forest management and prioritize long-term monitoring of trends of plant functional composition to maintain ecosystem functions and services.
Plant functional composition (i.e., CWM and CWV) could indicate woodland changes from the perspective of plant functional composition(Lavorel et al.,2011).Plant height and leaf traits such as leaf dry matter content can reflect land cover and land use at the large scale, i.e., the continental scales(Gross et al.,2007).At the continental scales,trait axes such as wood density and seed length should be applied to assess the ecosystem services lying at the core of the interactions between humans and forest ecosystems (Lavorel and Grigulis, 2012).Spatial scale effects exist widely for plant functional composition from local to continental scales (Qian and Ricklefs, 2012; Richardson et al., 2014; Stein et al.,2014).However,similar leaf traits(e.g.,specific leaf area and leaf C and N concentrations) may still exist, contributing to redundancy in the distributions of plant functional traits at the continental scales (Pillar et al., 2013; Cadotte et al., 2015).Furthermore, a wide variety of functional traits would have the ability to buffer the impact of a perturbation due to a longer woodland time (Ficken and Rooney, 2020).Hence, it is possible that historical woodland changes leave a legacy that impacts contemporary plant functional composition at the continental scales.
Application of plant functional composition on forest management practices should depend on biome types.Different biomes (e.g., nonforest and forest biomes) were subjected to various human land-use and resource management strategies for plant diversity from AD 0 to 2017(Hansen et al.,2001).Spatial patterns of agricultural land use made large contributions to historical woodland changes (Goldewijk et al.,2017;Pérez-Hoyos et al.,2017).The expansion of cropland and pasture has been extensive across various biome types (Ramankutty and Foley,1999;Goldewijk et al.,2017).In current and past periods,cropland has been largely rainfed, and pasture areas can be found in most regions of the world(Goldewijk et al.,2017;Pérez-Hoyos et al.,2017).Thus,plant functional composition can be affected by historical woodland changes through the conversion from forest to cropland.The clearing of a forest is generally conducted if insufficient natural grassland is available for grazing, suggesting that historical woodland changes have effects on plant functional composition because of conversion from forest to pasture(Goldewijk et al.,2017;Filazzola et al.,2020;Guerra et al., 2020).
4.5.Limitations
Although our work provides an innovative perspective on plant community assembly and advances our understanding of the role historical woodland changes play under plant community succession to a substantial degree, it has two limitations.1) Our study did not consider the effects of climate factors on plant functional composition.The distribution patterns of functional trait diversity are predicted better when including historical as well as contemporary climate predictors(Blonder et al.,2018).Given the Late Quaternary climate effects on contemporary plant functional composition (Blonder et al., 2018), legacy effects of environmental changes on contemporary plant functional composition may contribute to the understanding of the mechanisms underlying large-scale distribution patterns of plant functional composition to accurately predict assemblage responses to future land use and land cover.2)Due to the limitations on data selection,there are uncertainties in assessing woodland changes in historical periods.A key shortcoming for application of the reconstruction on a fine spatial scale is the implicit assumption of constant land use patterns over time (Goldewijk et al.,2017).Hence, the intervals between different periods of woodland changes appear inconsistent.A 1000-year interval is followed by a 700-year interval before 1700, while 90-year or less-than-ten-year intervals follow 1700-2017.These uncertainties may affect the accuracy of legacy effects of historical woodland changes on contemporary plant functional composition.The redundancy in the distributions of plant functional traits may be more significant at current than past periods.However, human activities are heavier at current than past periods,leading to a higher impact on plants.Thus, the time intervals should be shortened from past to current periods.
5.Conclusions
We conclude that historical woodland changes (i.e., the time of woodland existence) affect contemporary plant functional composition on a global scale over time.Furthermore,the legacy effects of historical woodland changes on contemporary plant functional composition depend on types of traits and biomes.Particularly, the CWM of plant height and seed traits showed the strongest correlations with woodland changes from AD 1910 to 2010 in tropics with year-round rain,and the CWM of leaf traits correlated with woodland changes from AD 0 to 1700 in tropics with summer rain.We provide the first evidence of legacy effects of historical woodland changes on contemporary plant functional composition, which are crucial for effective management of plant diversity and assessment of ecosystem functions and services from local to global scales.
Authors’contributions
Ji-Zhong Wan and Chun-Jing Wang conceived the study, performed all analyses,and wrote the paper equally.
Funding
We acknowledge support from the National Natural Science Foundation of China (NSFC; 32060385 and 31860668), and the Project of Qinghai Science&Technology Department(2020-ZJ-733).
Data accessibility statement
All the data on plant functional traits and woodland changes used in this manuscript are publicly available and sources described in the text.
Ethics approval
Not applicable.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.i.org/10.1016/j.fecs.2023.100142.

- Forest Ecosystems的其它文章
- Growth phenology adjusts to seasonal changes in water availability in coexisting evergreen and deciduous mediterranean oaks
- Assessing spatiotemporal variations of forest carbon density using bi-temporal discrete aerial laser scanning data in Chinese boreal forests
- Stabilizing forest productivity and resilience at multiple scales
- Habitat heterogeneity and biotic interactions mediate climate influences on seedling survival in a temperate forest
- Trees species’ dispersal mode and habitat heterogeneity shape negative density dependence in a temperate forest
- Tree sapling vitality and recovery following the unprecedented 2018 drought in central Europe