
罗汉松幼苗在不同磷环境下添加氮的生理响应权佳惠
2023-11-11 15:55:02招礼军朱栗琼邱广龙
热带作物学报 2023年9期
招礼军 朱栗琼 邱广龙



关键词:罗汉松;幼苗;氮;磷;生理响应;光合作用;养分
中图分类号:S791.46 文献标识码:A
罗汉松(Podocarpus macrophyllus)为罗汉松科(Podocarpaceae)常绿针叶乔木或者灌木。罗汉松枝、叶、种子、果实和根皮均具有药用价值,对于心胃痛、血虚以及风湿和跌打损伤有较好的治疗效果[1],其种子提取物对胃癌细胞有明显的抑制作用[2]。罗汉松树形雅致且具有较强的吸附污染物能力[3],常被用作庭院观赏树种和街道绿化树种,而小型的盆栽罗汉松则是送友佳品,具有较高的经济价值[4],随着近年来人们养生观念的发展,罗汉松的药用价值需求空前,原生环境罗汉松数量稀缺,人工繁育成为培育罗汉松的主流方式,并带动了相关产业的发展[5]。高产、保质和节约成本是人工培育罗汉松的重要考虑因素,合理的氮磷施肥量有益于助力罗汉松在人工培育过程中实现价值最大化,促进产业经济效益提高。
土壤环境中的氮磷元素是参与并影响植物光合作用和生理代谢的重要环境因子,对植物的生长发育至关重要,土壤环境中氮磷元素的添加能够补充土壤养分,解除或减缓植物的生长限制[6]。亚热带地区土壤酸化度高[7],虽土壤环境中磷含量丰富,但大多数易与金属元素结合固定在土壤中,植物可利用的有效磷含量低[8],磷素缺乏会限制植物光合速率,使ATP 合成受阻,植物生长发育缓慢。我国南方面临严重的氮沉降问题,大气中的氮素通过干沉降直接进入植物气孔参与植物生长代谢或沉降到地表土壤中[9],氮素是植物叶绿体内相关酶合成的重要元素[10],氮肥过低植物叶绿素合成减缓,植物光合效率受限,植物生理反应过程中相关酶活性减弱,植物代谢缓慢;氮肥浓度过高则对植物产生胁迫[7]。此外,人工林场在罗汉松种植過程中存在肥料施加随意,过度施肥等问题[11],罗汉松生产力和人工培育成活率受限,经济效益大打折扣。罗汉松生长年限较长,生长缓慢,基于土壤环境磷限制和大气氮沉降现状,探究不同土壤磷环境下,土壤氮素含量增加时罗汉松幼苗生理活性、叶绿素含量、光合特征和叶片养分含量的响应机制,对人工大规模种植罗汉松时科学施肥提供理论依据、保护土壤可持续发展、提高罗汉松人工种植林地生产力和土地资源的有效性具有重要意义。因此,本研究以盆栽法培育的罗汉松幼苗为研究对象,设置低磷、高磷和低、中、高3 个氮浓度的试验,以期对罗汉松的人工种植、室内培育和土壤资源的有效利用等方面起到科学的指导作用。
1 材料与方法
1.1 研究地概况
研究地点位于广西壮族自治区南宁市广西大学林学院校内教学基地苗圃温室大棚内,地理位置为22°50′N,108°17′E,大棚全年为南亚热带季风气候, 日照充足, 雨水充沛, 年均气温为21.6 ℃,夏季最热平均气温为28.2 ℃,冬季最冷平均气温为12.8 ℃,相对湿度为79%[12]。
1.2 实验材料
2019 年9 月中旬在广西壮族自治区南宁市隆安县金穗园林花木场购买平均株高为(49.9±0.78)cm、平均基径为(6.90±0.11)mm 无病虫害的2年生罗汉松幼苗作为实验材料。2019 年9 月末,将罗汉松幼苗根系泥土用清水冲洗干净,移植到长宽高为31.5 cm×31.5 cm×22.5 cm的塑料花盆中(带底座),每盆装红壤土∶沙土=1∶1 混合土壤15 kg,每盆1 株,试验幼苗培养于大棚内,缓苗2 个月后进行施肥处理。施肥实验前测得种植罗汉松幼苗土壤背景值如下:pH 为7.32±0.33,全氮(TN)为(0.99±0.06)g/kg,全磷(TP)为(0.28±0.08)g/kg,全钾(TK)为(11.29±1.15)g/kg,水解性氮为(111.09±8.13)mg/kg , 速效钾为(50.13±3.82)mg/kg,有机磷为(5.62±0.81)mg/kg。
1.3 实验设计
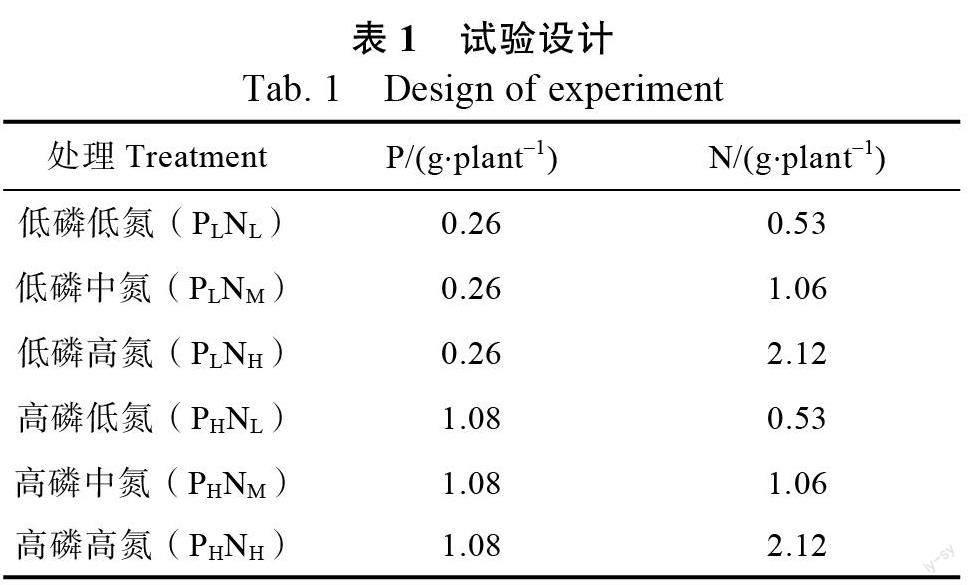
采用2×3 双因素正交实验设计(表1,2 种磷浓度,3 个氮浓度),每个处理3 个重复,每个重复5 盆,每盆种植1 株,共90 盆。供试肥料氮肥为尿素(含N 46.5%),磷肥为过磷酸钙(含PO 12.0%)。具体浓度及编号见表1,2019 年11 月末,将肥料按表1 的含量溶于1000 mL 水中,缓缓浇入幼苗根部,在整个实验期间保持各处理组供给一致,分别于2020 年的2 月(春季)、5月(夏季)、8 月(秋季)和10 月(冬季)末在天气晴朗的早上8 点采集测定叶绿素和生理指标的叶片;9:00—11:00 进行叶片气体交换参数的测定;养分含量测定所需叶片在实验结束时采集(10 月末)。
1.4 方法
1.4.1 叶绿素含量和生理指标的测定 在各处理组选取3 株长势良好、无病虫害的罗汉松幼苗,在树冠中部各个方向上选择10~12 片完整的成熟功能叶片,放入冰盒,带回实验室测定。采用氮蓝四唑(NBT)光化还原法测定超氧化歧物酶(SOD)活性;采用考马斯亮蓝G-250 染色法测定可溶性蛋白含量;采用酸性茚三酮比色法测定游离脯氨酸(Pro)含量;采用硫代巴比妥酸显色法测定丙二醛(MDA)含量;采用丙酮-乙醇混合液提取法测定[13]叶绿素含量;采用愈创木酚比色法测定[14]过氧化物酶(POD)活性;采用蒽酮比色法测定[15]可溶性糖含量。
1.4.2 叶片光合生理指标的测定 从各处理组挑选3 株无病虫害且长势基本一致的完整植株叶片,采用Li-6400 便携式光合测定仪每株取3 片完整的成熟叶片进行光合参数测定,得到罗汉松幼苗叶片净光合速率(P)、细胞间CO浓度(C)、气孔导度(G)和蒸腾速率(T)。
1.4.3 叶片养分含量测定 从各处理组叶片中随机选择2~3 片测定植物体内养分含量。叶片经HSO-HO 消煮后,使用凯氏定氮法测定罗汉松幼苗叶片全氮含量,使用钼锑抗比色法测定植物全磷含量[16]。
1.5 数据处理
运用Excel 软件对数据进行整理,利用SPSS25.0 软件进行双因素方差分析,使用origin 2018软件制图。表格和图中的数据均以平均值±标准偏差表示。
2 结果与分析
2.1 不同磷环境下施加氮肥罗汉松幼苗生理响应
由表2 可知,罗汉松幼苗可溶性蛋白含量受氮添加主效应及氮磷交互效应影响,POD 活性则同时受到氮添加和环境磷含量主效应影响。如图1 所示,罗汉松幼苗可溶性糖、Pro、MDA 含量和SOD 活性在实验期间均呈先降低后增加的趋势,POD 活性和可溶性蛋白含量则呈无规律的变化趋势。罗汉松幼苗可溶性蛋白和可溶性糖含量在低磷环境下随着氮浓度的增加而升高,与高氮浓度相比,低氮浓度平均增加35.4%和26.6%;在高磷环境下可溶性蛋白和可溶性糖含量,随着氮肥浓度的增加呈先增加后降低,中氮浓度下含量最高,高氮浓度则明显降低了可溶性蛋白和可溶性糖积累,与低氮浓度相比,分别平均下降了16.7%和4.7%。随着磷肥浓度的增加,低氮和中氮水平下,罗汉松幼苗可溶性糖和可溶性蛋白含量增加;相反,在高氮浓度下,增加磷肥浓度其含量下降, 较低磷环境分别下降了26.5% 和16.2%;氮肥和磷肥浓度增加均促进了POD 活性升高。
双因素方差分析表明(表2),罗汉松幼苗Pro含量同时受环境磷含量主效应和氮磷交互效应的影响,SOD 活性主要受氮添加和环境磷含量主效应的影响,MDA 含量则受氮磷交互效应影响显著。由图1 可知,在不同的磷环境下,中浓度氮肥促进了罗汉松幼苗SOD 活性,低磷和高磷环境分别比低浓度氮肥处理组增加10.6%和5.9%,而高氮浓度则降低了SOD 活性;增加氮肥浓度,低磷环境下MDA 和Pro 含量均下降,高磷环境下MDA 含量增加,中氮浓度Pro 含量降低,高氮浓度Pro 含量上升。相同氮水平下,罗汉松幼苗SOD活性随磷肥浓度增加而升高,而Pro 和MDA 含量则在低氮和中氮浓度随着磷肥的增加而降低,高氮浓度随着磷肥浓度的增加而增加。
2.2 不同磷环境下施加氮肥罗汉松幼苗叶绿素含量变化
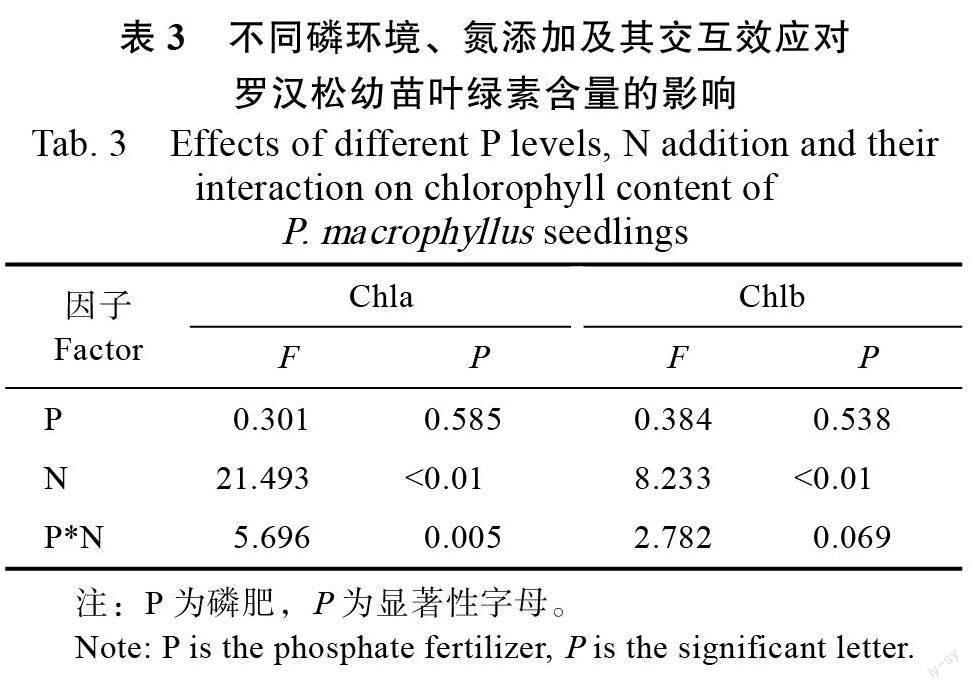
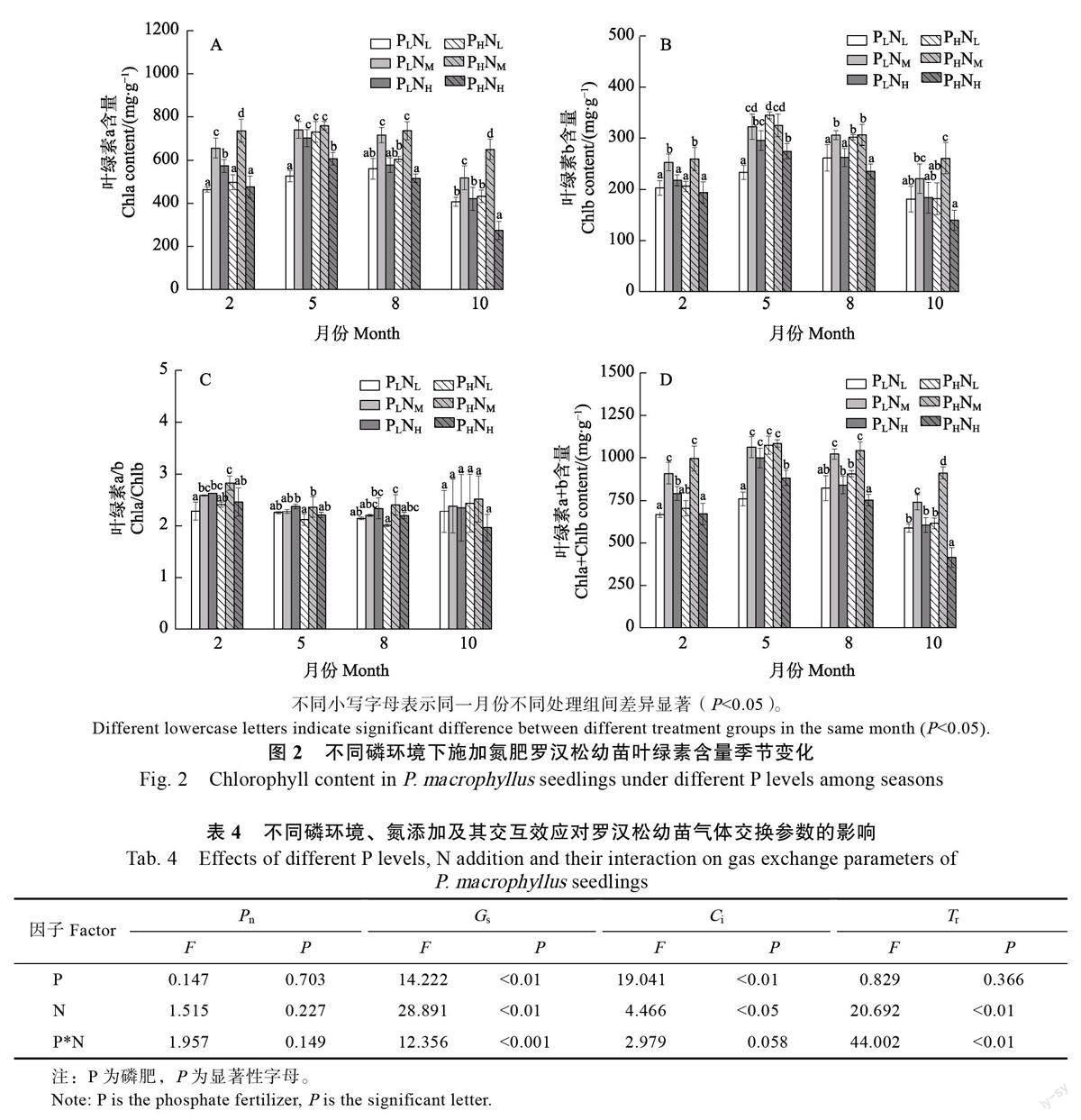
氮素是植物合成叶绿素的重要原料,双因素方差分析表明,叶绿素a(Chla)和叶绿素b(Chlb)仅受氮添加主效应影响(表3)。罗汉松幼苗叶绿素含量在整个实验期间呈现先增后降的变化趋势。在图2 中,高磷环境下,高氮处理组罗汉松幼苗叶绿素a/叶绿素b 含量在整个实验期间呈逐渐降低的趋势,其余处理组则夏季和秋季上升,冬季下降。中浓度的氮肥在低磷和高磷环境下均显著促进了罗汉松幼苗叶绿素a 和叶绿素b 的含量积累,相对低氮处理组平均增幅为32.45%和20.8%;低磷和高磷环境下,高浓度氮肥均减少了叶绿素a 和叶绿素b 含量的积累,低磷环境下高氮处理组低于中氮处理组但高于低氮处理组,高磷环境下高氮处理组低于低氮处理组,相较低氮处理组平均下降18.1%。低氮和中氮濃度下,磷肥浓度增加罗汉松幼苗叶绿素a、b 和总叶绿素含量增加,相反,高氮处理下磷肥浓度增加,罗汉松幼苗叶绿素a、b 和总叶绿素含量的增加量减少。
2.3 不同磷环境下施加氮肥罗汉松幼苗光合生理特性响应
氮肥能够通过影响植物叶绿素含量间接影响植物光合作用。双因素方差分析结果表明,罗汉松幼苗叶片G 同时受氮添加、环境磷含量主效应及二者交互效应影响,C 受氮添加和环境磷含量主效应影响,T 则主要受氮添加主效应及氮磷交互效应的影响(表4)。如图3 所示,罗汉松幼苗P 和C 夏季和秋季高,春季和冬季低,G 和T各处理变化趋势不一,无统一的规律。随着氮肥浓度增加,低磷环境高氮处理罗汉松幼苗叶片的P、G 和T 最高,分别较低氮处理上升28.5%、36.15%和50.3%;高磷环境中氮处理组P、G 和T 最高,显著高于低氮和高氮处理组,高氮浓度下P、G 和T降低;C 则随氮浓度的增加低磷环境下降低,高磷环境下先降后升。低氮和中氮水平高磷环境罗汉松幼苗叶片P、T和G 较低磷环境升高,高氮水平高磷环境P、T 和G 较低磷环境下降;相同氮水平下,磷浓度增加罗汉松幼苗Ci 增加。
2.4 不同磷环境下施加氮肥罗汉松幼苗叶片养分含量变化
由表5 可知,罗汉松幼苗叶片N 含量仅受氮磷交互效应影响,P 肥则同时受到氮添加、环境磷含量主效应及二者交互效应影响。叶片养分含量关系植物营养状况,如图4 所示,随着氮肥浓度增加,低磷环境中氮和高氮处理组罗汉松幼苗叶片N 含量较低氮处理组显著增加(图4A),高磷环境下则随氮肥浓度增加而降低;供氮浓度对叶片P 含量的影响均表现为随着供氮浓度的增加,叶片P 含量下降(图4B),且高磷环境下低氮、中氮和高氮处理组间差异显著(P<0.05)。低氮水平下,高磷环境罗汉松幼苗叶片N 含量较低磷环境显著增加,中氮和高氮水平则降低;高磷环境显著促进了叶片P 含量的积累,其中,低氮和中氮处理组间差异显著(P<0.05),高氮处理则差异不显著(P>0.05)。
2.5 罗汉松幼苗各指标相关性分析
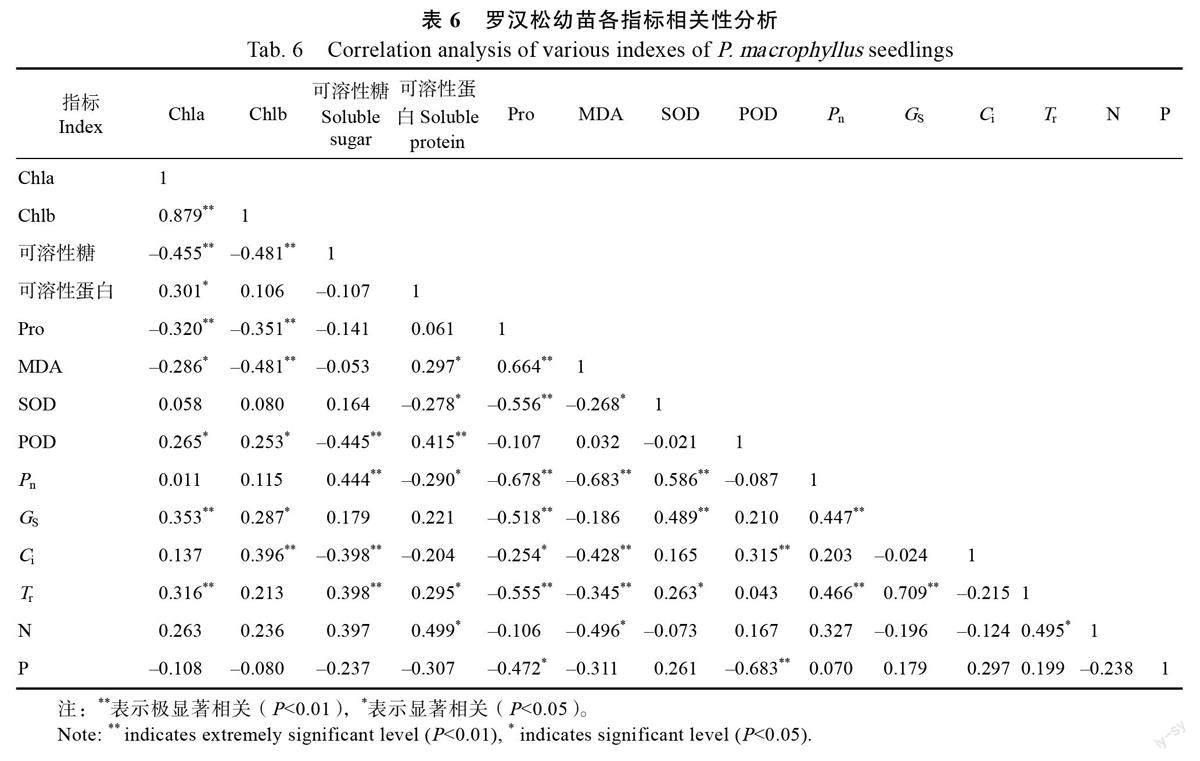
植物体内的各指标协同作用,共同为植物的生长发育提供保障。如表6 所示,各指标相关性分析表明,罗汉松幼苗叶绿素含量与Tr、POD、Gs、可溶性蛋白和Ci 呈显著正相关;可溶性糖含量与P和T 呈显著正相关,可溶性蛋白含量与MDA、POD和T 呈显著正相关;抗氧化酶SOD 与P、G 和T呈显著正相关,POD 与C呈极显著正相关;叶片Pro 含量和MDA 含量呈极显著正相关;罗汉松幼苗光合生理系统相关指标之间存在极显著的正相关关系,Tr 与叶片N 含量呈显著正相关。
3 讨论
3.1 罗汉松幼苗生理特性对不同氮磷浓度的响应
植物的生理特征是植物体内物质代谢的表征,外界环境变化会引起植物体内代谢进程改变。SOD 酶和POD 酶是植物在长期适应环境中进化出的保护酶,植物通过调节酶活性来清除体内活性氧,维持植物正常的生理代谢[17];Pro 是植物体内重要的渗透调节物质,对于胁迫环境下植物细胞膜和酶的保护具有重要意义,MDA 是植物处于逆境时体内积累的活性氧加剧细胞膜脂过氧化的主要产物,是量化细胞膜受损伤程度的重要指标之一[18],可溶性蛋白能够提高细胞保水能力,可溶性糖是植物体内生物化学进程的表征[19];研究表明,施加適量氮肥能够减缓细胞膜损伤,提高植物内源酶活性,降低MDA 含量[20],促进植物可溶性糖和可溶性蛋白的积累,缓解植物在原生环境下受到的伤害,而过高的氮则会引起植物对胁迫环境的敏感。杨洁等[21]研究,POD 活性在不同氮水平下活性不同,中氮和高氮促进了POD活性,低氮则活性降低;在本研究中,在不同的磷环境下,在中氮浓度下SOD 活性最高,高氮浓度下SOD 活性下降,POD 活性则随着氮肥浓度的增加而升高,这表明高氮浓度降低了SOD 活性,却促进了POD 活性,低磷环境下POD 活性的增高清除了植物在原生环境积累的大量活性氧,减缓了原生环境下细胞膜损伤程度,植物体内Pro 含量和MDA 含量下降,罗汉松幼苗体内物质代谢加快,可溶性糖和可溶性蛋白含量增加;高磷环境下,高氮水平则对植物生理构成了胁迫,植物体内产生大量自由基,POD 对活性氧清除效率下降,植物体内Pro 含量增加以增强对氧化酶的保护,从而减缓活性氧对细胞膜损伤,但大量活性氧加剧了植物细胞膜脂过氧化,MDA 含量增加, 罗汉松幼苗物质代谢效率降低,可溶性糖和可溶性蛋白含量降低。此外,在本研究中,高磷环境下不同氮水平罗汉松幼苗POD 和SOD 活性要高于低磷环境下其活性,而高磷环境下高氮浓度则成为罗汉松幼苗生理代谢胁迫因子,可溶性糖和可溶性蛋白含量相较低磷环境下降,Pro 和MDA 含量增加。相关性分析表明,可溶性糖和可溶性蛋白是植物能源物质,光合作用的增强和叶绿素含量的积累对其含量的积累有正向的促进作用,酶活性的增加清除植物体内自由基,促进了植物叶绿素含量的积累,Pro 和MDA 是反映植物抗性的指标,均在植物受到胁迫时增加,二者具有正向的相关性。
3.2 罗汉松幼苗光合特征对不同氮磷浓度的响应
叶绿素是植物吸收和利用光能,产生能量物质的基础,其含量能够反映植物光合作用大小,常被作为指导农业施肥的依据[22];植物叶片P、G、C 和T 是表征植物光合作用内在特征的重要指标[23]。在本研究中,罗汉松幼苗叶绿素a、b与P、T和G 变化趋势相同,均在夏季和秋季升高,冬季降低,这主要是由于夏秋光合有效辐射最强,叶绿素可捕获的光能增多,提升了罗汉松幼苗Pn,叶片气孔扩张,为减少高温对叶片的灼伤,幼苗叶片Tr 增强[24]。土壤环境中氮磷元素影响着植物的生长发育,主要参与构成植物叶绿体中类囊体蛋白氮[7],植物生长环境中氮磷缺乏时,施加氮磷肥能够显著促进植物叶绿素含量增加和光合速率提升[25],研究表明增加土壤氮肥浓度可促进冬小麦旗叶(Triticum aestivum)[26]和虎耳草(Saxifraga stolonifera)[27]叶片叶绿素合成,显著提升其光合效率,而植物生长环境中氮磷元素过饱和时,施加氮磷肥则对植物光合作用产生抑制[28];在本研究中,低磷环境下低氮和中氮浓度罗汉松幼苗叶绿素含量的增加促进了罗汉松幼苗Pn 的效率,叶片气孔扩张,加速了对CO 的吸收和转化,C值低,气体交换速率增加,Tr 增强,高氮浓度则减缓了罗汉松幼苗叶绿素增加,但未超过植物的耐受范围,因此植物通过提高Pn 和Tr,扩张气孔加速对CO 能源物质的吸收来补偿氮肥浓度过高产生的影响,从而维持体内稳态环境,C 值降低;高磷环境下,低氮和中氮同样促进了罗汉松幼苗叶绿素积累,植物光合效率提高,而磷过饱和高浓度氮肥成为罗汉松幼苗环境胁迫因子,其叶绿素合成受限,叶绿素a/叶绿素b 含量降低,抗性下降,叶片Pn 和Tr 效率较低,罗汉松幼苗虽通过扩张气孔,加速对细胞间CO 的吸收来弥补高磷环境下高氮浓度产生的胁迫,但对细胞间CO的利用效率下降,Ci 值升高。
此外,气孔限制和非气孔限制均会导致植物P 降低,当P 与G 变化趋势一致时,P 的下降由G 引起,相反,则由非气孔因素叶肉细胞的羧化能力降低引起[29]。在本研究中,随着施加氮肥或磷肥浓度的增加,罗汉松幼苗叶片P 与G 变化趋势一致,这表明气孔因素是导致罗汉松幼苗Pn 下降的主要限制因子。土壤环境中磷稀缺和磷饱和均会降低米槁(Cinnamomum migao)卡尔文循环中关键酶的活性,使其光合能力大幅降低[30];在本研究中,低氮和中氮浓度下磷肥浓度增加促进了罗汉松幼苗叶绿素含量增加,叶片光合效率提升,高氮浓度下磷肥浓度增加则限制了叶片叶绿素合成,光合效率降低,这可能是因为高氮浓度下高磷环境磷元素过饱和,限制了罗汉松幼苗叶绿素积累和植物光合效率增加。相关性分析表明Pn 影响植物光合速率,继而影响可溶性糖含量的积累,P 主要受气孔因素的限制。
3.3 罗汉松幼苗叶片养分含量对不同氮磷浓度的响
应植物叶片养分含量是判断植物营养状况的基础,其关系植物光合效率和生理活性[31]。在本研究中,低磷环境下罗汉松幼苗叶片氮含量随施加氮肥浓度的增加而显著增加,这与大多数研究结果一致[32-33],说明低磷的环境下,氮输入增加了土壤可利用氮含量,植物对氮素的积累增加;高磷环境下,随着施氮浓度的增加,罗汉松幼苗叶片氮含量下降,原因可能是因为高磷环境阻碍了植物对氮素的吸收和积累[34];低磷和高磷环境下,罗汉松幼苗叶片磷含量随着添加氮浓度的增加下降,这是因为土壤氮含量的增加间接影响了土壤可利用磷含量[35],罗汉松幼苗可吸收利用的磷素含量降低;此外,高磷环境下罗汉松幼苗叶片磷含量高于低磷环境下其含量,这与盖甜甜等[36]的研究结果一致,低磷环境下,罗汉松幼苗受环境磷限制,叶片磷含量积累少,增加磷肥浓度后,叶片磷含量显著增加[7],因此在土壤磷含量较低时,通过施加磷肥能够增加罗汉松幼苗叶片磷含量。在本研究中,罗汉松幼苗叶片氮磷比在3.3~13.6 之间,这表明罗汉松幼苗生长可能主要受氮限制,其叶绿素含量和光合作用主要受氮添加主效应的调控。
综上所述,不同磷环境下施加适量的氮肥促进了罗汉松幼苗叶绿素含量的积累和光合效率的提升,且氮磷浓度的增加明显提升了罗汉松幼苗体内物质代谢进程,植物在原生环境下积累的损伤得到恢复,但过高的氮磷浓度则成为罗汉松幼苗环境胁迫因子,不利于其生长,在罗汉松繁殖和培育过程中,应合理施加氮磷肥浓度,高氮高磷浓度会影响其正常生长发育。
猜你喜欢
作文大王·低年级(2023年2期)2023-02-10 13:20:59
科学(2022年4期)2022-10-25 02:43:00
落叶果树(2021年6期)2021-02-12 01:28:54
农村实用技术(2020年4期)2020-12-15 15:53:29
文苑(2020年12期)2020-04-13 00:54:10
林业与生态(2019年7期)2019-08-06 17:34:30
海峡姐妹(2019年1期)2019-03-23 02:42:46
现代园艺(2017年23期)2018-01-18 06:57:42
Coco薇(2016年5期)2016-06-03 09:17:41
考试周刊(2016年6期)2016-03-11 08:14:32
