
溴虫氟苯双酰胺对草地贪夜蛾的亚致死效应
2023-11-09 12:31谢云茜崔汝强彭文文傅小香石绪根
环境昆虫学报 2023年5期
张 勇,宁 旭,谢云茜,王 敏,崔汝强,彭文文,傅小香,石绪根**
(1.江西农业大学农学院,南昌 330045;2. 江西农业大学国土资源与环境学院,南昌 330045)
草地贪夜蛾Spodopterafrugiperda,是一种杂食性鳞翅目害虫,原产于美洲热带和亚热带地区,具有较强的迁飞性、繁殖能力强,生活周期短,可危害玉米、甘蔗、高粱、大麦和水稻等353种植物,并造成严重经济损失,2018年联合国粮农组织将其列为全球预警的超级害虫(FAO, 2018;Liuetal., 2020)。自2019年1月,草地贪夜蛾从云南入侵我国后,不到一年时间迅速蔓延至全国20余省(自治区、直辖市)发生为害(杨学礼等, 2019; 章婉贤等, 2019)。
化学防治具有见效迅速、防效彻底等特点,被广泛用于爆发性和突发性有害生物的控制(吴孔明, 2018)。目前,在农业生产中,草地贪夜蛾的防治主要以化学防治为主。然而,在农业害虫防治过程中,抗药性的产生是降低农药防治效果的主要原因之一。目前草地贪夜蛾已在全世界范围内对超过40种杀虫剂产生了不同程度的抗性,其中既包括有机磷类、氨基甲酸酯类和拟除虫菊酯类等传统杀虫剂,也包括双酰胺类、多杀菌素等新型杀虫剂(Sparksetal., 2020)。入侵我国的草地贪夜蛾同样对有机磷类等传统杀虫剂产生了高水平抗性(Zhangetal., 2021),而对氯虫苯甲酰胺等双酰胺类药剂的抗性处于较低水平(Lvetal., 2021)。但已有证据表明,该虫对氯虫苯甲酰胺、乙基多杀菌素等新型杀虫剂的抗性风险很高(Gutiérrez-Morenoetal., 2018; Boaventuraetal., 2020; Liraetal., 2020)。
溴虫氟苯双酰胺是一种新型γ-氨基丁酸(GABA)门控氯离子通道别构调节剂,不同于氯虫苯甲酰胺(作用于鱼尼丁受体)的作用机理,其作用于昆虫神经细胞中的GABA受体,别构抑制GABA激活的Cl-通道,抑制Cl-向神经细胞传递,引起受药昆虫过度的兴奋和抽搐,最后实现快速的杀虫作用(柳爱平等, 2020)。
草地贪夜蛾对化学杀虫剂的代谢抗性机制,主要包括微粒体多功能氧化酶、谷胱甘肽-S-转移酶和酯酶等相关酶系代谢能力的增强(Amezianetal., 2021; Hilliouetal., 2021)。保护酶与昆虫防御外来有害物质相关,可以避免害虫体内膜系统受损,降低虫体于逆境中所受伤害,其活性增强亦可能增强害虫对化学杀虫剂的抵抗力(Bolteretal., 1990)。
在田间施用化学杀虫剂时往往会存在药剂低剂量残留或喷洒不均匀,会对田间害虫产生亚致死效应(全林发等, 2016; Pérez-Aguilaretal., 2018)。化学杀虫剂对昆虫的亚致死效应涉及的学科领域包括害虫行为学、生物学、生理学及抗药性发展等很多方面,有关研究可为药剂的合理使用和抗药性治理提供一定的理论依据(Dongetal., 2017; 梁炜博等, 2017; Jungetal., 2018)。有关溴虫氟苯双酰胺对草地贪夜蛾的亚致死效应目前缺乏研究,因此,本研究采用亚致死剂量溴虫氟苯双酰胺处理草地贪夜蛾3龄幼虫,研究其对草地贪夜蛾生物学特性、体内解毒酶和保护酶活性的影响,以期为大田作物生产中合理使用全新杀虫剂溴虫氟苯双酰胺提供依据。
1 材料与方法
1.1 供试虫源
供试草地贪夜蛾种群幼虫于2020年6月采自江西省南昌市新建区乐化镇(115.87° W,28.82° N)前期未施过药的玉米田。将采集的幼虫在人工气候箱内(温度25±1℃、相对湿度70%±5%、光周期L∶D=16 h∶8 h)用新鲜未施药的玉米叶片饲养,成虫以10%蜂蜜水提供营养。以F10代3龄初孵幼虫作为试验虫源。
1.2 供试药剂
97.0%溴虫氟苯双酰胺原药(Broflanilide)来源于德国巴斯夫公司;乙二胺四乙酸(EDTA)来源于石家庄杰克化工有限公司;苯基硫脲(PTU)来源于上海阿拉丁生化科技有限公司;二硫苏糖醇(DTT)来源于上海吉至生化科技有限公司;苯甲基硫酰氟(PMSF)、二硝基氯苯(CDNB)均来源于上海恪敏生物科技有限公司;牛血清蛋白(BSA)来源于上海蓝季生物有限公司;十二水磷酸氢二钠(Na2HPO4·12H2O)、十二水磷酸二氢钠(NaH2PO4·12H2O)均来源于天津市大茂化学制剂厂;多功能氧化酶(MFO)酶活性测定试剂盒(ELISA法)购自江苏酶免实业有限公司;超氧化物歧化酶(SOD)、过氧化物酶(POD)及过氧化氢酶(CAT)酶活性测定试剂盒购自南京建成生物工程研究所;底物和显色剂混合液:0.2 mol/L pH6.0 PBS与固蓝RR盐与100 mmol/L α-NA乙醇溶液按1∶2∶0.01比例混合(定性滤纸过滤,并现用现配)。
1.3 试验仪器
人工气候箱(上海新苗QHX-300BSH-Ⅲ)、万分之一天平(日本岛津ATX224型)、手动微量点滴仪(Burkard PDE0003)、培养皿(直径9 cm)、酶标仪(SpectraMax190)、高速冷冻离心机(Eppendorf Centrifuge5430R)、涡旋仪(美国SIvortex-genie2)、玻璃匀浆器(5 mL)。
1.4 试验方法
1.4.1室内毒力测定方法
用点滴法对玉米草地贪夜蛾3龄初孵幼虫进行毒力测定,虫重平均3 mg。将原药用丙酮配置成1×104mg/L的母液,然后用丙酮将母液稀释成5个系列浓度梯度(10、15、20、25、30 μg/mL),并用丙酮处理作空白对照。用微量点滴器将药液逐个点滴于3龄初孵幼虫(长度约0.8 cm)的前胸背板上,每头0.2 μL,点滴后的幼虫放入培养皿(9 cm),给予充足的新鲜玉米叶片并每天更换。每浓度4次重复,每重复10头3龄幼虫。药剂处理后的试虫放在人工气候箱(温度25±1℃、相对湿度70%±5%、光周期L∶D=16 h∶8 h)中饲养。48 h后观察结果,毛笔轻触无反应或出现畸形、抽搐、停止取食等明显中毒症状,判定为死亡。记录死亡虫数、存活虫数,计算校正死亡率。
1.4.2试虫处理
1.4.2.1 生物学指标观察
利用室内生物测定测得的亚致死剂量(LD15和LD30),分别点滴处理草地贪夜蛾3龄幼虫。每浓度5次重复,每重复20头幼虫,并用丙酮处理组做对照,处理好的幼虫放入培养皿(9 cm),给予新鲜充足玉米叶片。48 h后将对应浓度剩余活虫单头饲养于培养皿中,每天定时给予新鲜且充足玉米叶片,每间隔24 h观察记载不同处理组各生物学指标。在各处理组中随机选取同日羽化的雌、雄成虫各1头配对,每对作为1次重复,每处理至少重复25次。将配对成虫放入保鲜袋(28 cm×25 cm)内,袋中放已浸泡10%蜂蜜水的脱脂棉,给予成虫补充营养,并用标签纸封口。每天更换保鲜袋和含10%蜂蜜水的脱脂棉,直到雌蛾死亡,收集并记录产于保鲜袋上的虫卵量,计算单雌产卵量。试虫的生长在人工气候箱(温度25±1℃、相对湿度70%±5%、光周期L∶D=16 h∶8 h)条件下进行。
1.4.2.2 酶活性试验
分别采用溴虫氟苯双酰胺LD15、LD30、LD503个剂量点滴处理草地贪夜蛾3龄初孵幼虫。每浓度5个重复,每重复30头幼虫,并用丙酮处理作对照。48 h后收集仍存活幼虫,液氮速冻,置于-80℃保存,作为酶活测定的样本备用。
1.4.3酶活性测定
1.4.3.1 解毒酶酶液制备
用万分之一电子天平,称取8~12 mg (3~4头)的3龄幼虫为一个样本,加入1.2 mL 0.1 mol/L磷酸缓冲液(pH7.6,含1 mmol/L EDTA,1 mmol/L DTT,1 mmol/LPTU,1 mmol/L PMSF,20%甘油)冰浴匀浆。放入4℃,13 000 g离心30 min,取上清液到新的1.5 mL离心管中。再次在4℃,13 000 g离心10 min,取上清液到新的1.5 mL离心管中。最后在4℃,13 000 g离心5 min,得到酶粗提液。将其稀释10倍(用0.1 mmol/L,pH7.6 PBS稀释),后用于谷胱甘肽-S-转移酶(GST)酶活力、羧酸酯酶(EST)酶活力及蛋白浓度测定。
MFO酶源制备:分别随机称取8~12 mg(3~4头)的3龄幼虫加入适量的预冷pH 7.2~7.4左右的PBS,然后用冰浴研磨器研磨,4℃离心20 min(2 000~3 000 r/min),取上清液作为待测酶液。
1.4.3.2 解毒酶活性测定
(1) GST活性测定
依据Yang等(2004)的方法测定GST活性。分别在96孔酶标板中加入稀释10倍的酶液10 μL、1.2 mmol/L CDNB 100μL、6 mmol/LGSH 100 μL。用酶标仪在波长340 nm处测定吸光值,温度设定为30℃,每10 s记录一次,反应20 min。
(2) EST活性测定
依据Yang等(2004)的方法测定EST活性。在96孔板中依次加入稀释10倍的酶液20 μL、底物和显色剂混合液205 μL,然后用酶标仪在450 nm处测定吸光值,温度设定为27℃,每30 s记录一次,反应10 min。
(3) MFO活性测定
MFO活性测定用酶联免疫法。在标准品孔中加入不同浓度的标准品50 μL,样本孔中先后加入待测样品10 μL和样本稀释液40 μL;两类孔中每孔加入辣根过氧化物酶(HRP)标记的检测抗体100 μL,恒温箱温育30 min,20倍洗涤液重复洗板5次,于每孔中加入底物A、B各50 μL,震荡混匀,37℃避光孵育15 min;每孔加入终止液50 μL,以空白孔调0,15 min内在450 nm波长下测定吸光度(OD值),计算酶的比活力。
1.4.3.3 总蛋白测定
参照Bradfold等的考马斯亮蓝(G-250)法(Bradford, 1976)测定。
1.4.3.4 保护酶酶源制备
分别取上述试虫3~4头(约8~12 mg),加入适量的预冷匀浆液后冰浴研磨,4℃下12 000 r/min离心10 min,取上清液作为待测酶液。
1.4.3.5 保护酶活性测定
均按照南京建成对应试剂盒说明书进行测定。
(1) SOD活性测定(羟胺法)
SOD活性测定:以SOD抑制率达50%时所对应的酶量为1个SOD活力单位(U)。
SOD抑制率(%)={(对照OD值-对照空白OD值)-(测定OD值-测定空白OD值)}/(对照OD值-对照空白OD值)×100
SOD活性=SOD抑制率/(50%)×反应体系稀释倍数/待测样本蛋白浓度
(2) POD活性测定
POD活性测定: 以1 mg组织蛋白在37℃每分钟催化1 μg底物的酶量定义为1个酶活性单位(U)。
POD活性=(测定OD值-空白OD值)/(12×比色光径)×反应液总体积/样本量×1000/(反应时间×待测样本蛋白浓度)
(3) CAT活性测定(紫外法)
CAT活性测定: 以1 mg组织蛋白每秒分解1 μmol H2O2的量定义为1个酶活性单位(U)。
CAT活性={(对照OD值-测定OD值)×271×[1/(60×取样量)]}/待测样本蛋白浓度
蛋白含量测定:
待测样品蛋白浓度(μg/mL)=(测定OD值-空白OD值)/(标准OD值-空白OD值)×样品测试前稀释倍数×标准品浓度
1.5 数据处理
利用SPSS 25.0软件计算溴虫氟苯双酰胺对草地贪夜蛾3龄初孵幼虫的致死中量及95%置信限、相关系数等,采用Duncan’s新复极差(DMRT)法分析酶活数据的差异性。
2 结果与分析
2.1 溴虫氟苯双酰胺对草地贪夜蛾的室内毒力
溴虫氟苯双酰胺对草地贪夜蛾3龄幼虫室内毒力结果如表1所示:致死中量为0.50 μg/g,亚致死剂量LD15和LD30分别为0.30 μg/g和0.38 μg/g。
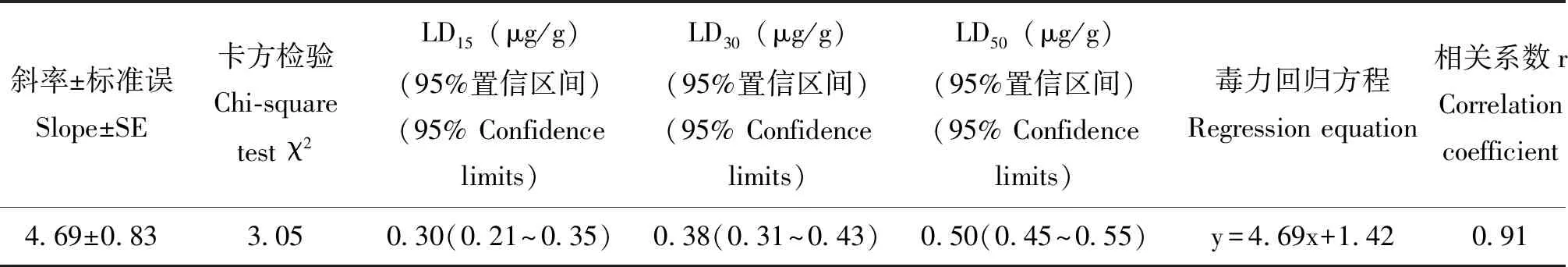
表1 溴虫氟苯双酰胺对草地贪夜蛾3龄幼虫的室内毒力
2.2 亚致死剂量溴虫氟苯双酰胺对草地贪夜蛾生物学特性的影响
在溴虫氟苯双酰胺亚致死剂量(LD15和LD30)胁迫下,草地贪夜蛾各生物学指标变化情况如表2所示。与对照相比,LD30处理能显著延长产卵前期和升高了成虫畸形率,但LD15处理的产卵前期和成虫畸形率与对照相比无明显差异。LD30处理较对照的产卵前期延长了14.9%,成虫畸形率升高了376%;幼虫存活率随处理剂量的增加显著降低。与对照相比,LD30处理显著降低蛹重、成虫重和单雌产卵量,并缩短产卵期,但LD15处理无明显差异。LD30处理较对照的蛹重、成虫重和单雌产卵量降低率分别4.5%、13.2%和20.3%,产卵期缩短率为11.8%。各处理间幼虫期、蛹期、化蛹率、雌雄性比、成虫羽化率无明显差异。
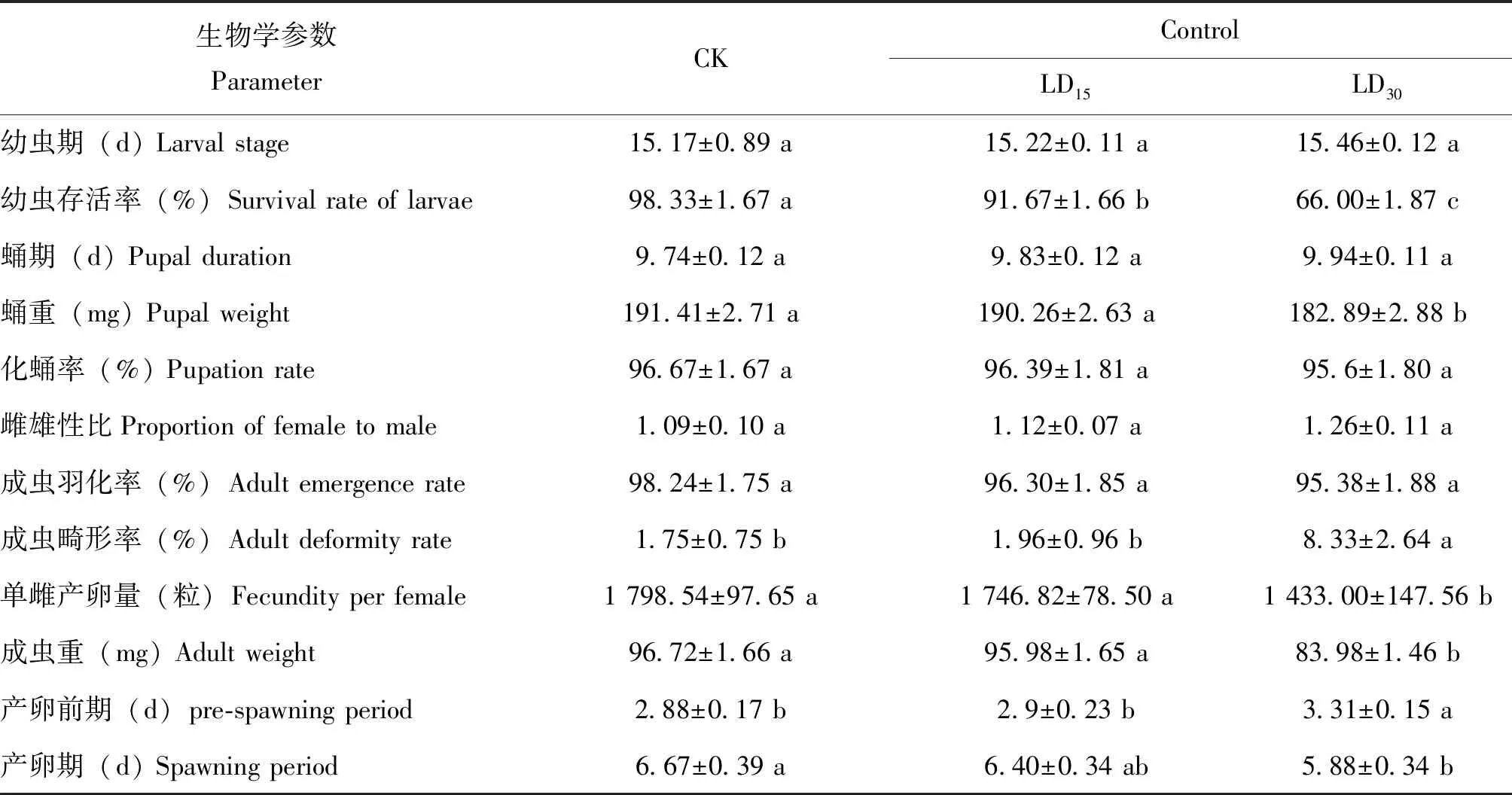
表2 溴虫氟苯双酰胺亚致死剂量处理对草地贪夜蛾存活和生长发育的影响
2.3 溴虫氟苯双酰胺对草地贪夜蛾体内解毒酶和保护酶活性的影响
2.3.1溴虫氟苯双酰胺对草地贪夜蛾体内解毒酶活性影响
不同剂量溴虫氟苯双酰胺(LD15、LD30和LD50)处理草地贪夜蛾后,其体内解毒酶活力变化情况如图1所示。结果表明,草地贪夜蛾体内GST和EST的比活力随药剂浓度的升高而增加,呈明显正相关。高剂量溴虫氟苯双酰胺(LD50)处理组GST和EST比活力最高,分别为对照组的1.84倍和1.54倍。LD30和LD50两个处理显著诱导草地贪夜蛾3龄幼虫体内MFO的比活力升高,LD30剂量处理组MFO的比活力最高,为对照组的1.33倍,但LD15处理体内MFO的比活力与对照组无差异。
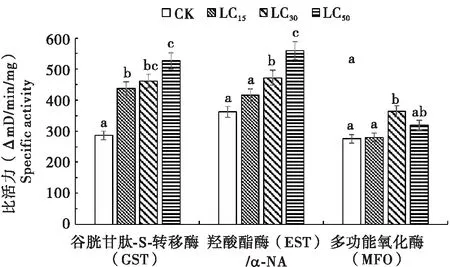
图1 溴虫氟苯双酰胺各剂量处理后草地贪夜蛾体内解毒酶活性比较
2.3.2溴虫氟苯双酰胺各剂量处理对草地贪夜蛾体内保护酶活性影响
不同剂量溴虫氟苯双酰胺(LD15、LD30和LD50)处理后,草地贪夜蛾体内保护酶的活性变化情况如图2所示。结果表明,各处理组SOD活性与对照组相比无显著性差异,POD和CAT的活性随药剂处理剂量的增加呈现先上升后降低的趋势,LD15和LD30处理能显著诱导CAT活性升高,LD30处理能显著提高POD活性,但LD50处理体内3种保护酶活性与对照无差异。其中,LD30处理体内POD和CAT活性最高,分别为对照组的1.34倍和1.35倍。
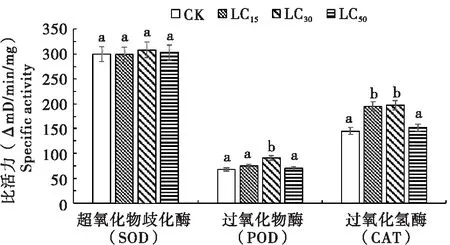
图2 溴虫氟苯双酰胺各剂量处理后草地贪夜蛾体内保护酶活性比较
3 结论与讨论
本研究利用点滴法测定了溴虫氟苯双酰胺对草地贪夜蛾3龄幼虫的室内毒力。结果显示其LD50值为0.50 μg/g,表明溴虫氟苯双酰胺对草地贪夜蛾具有很高的触杀活性。亚致死剂量溴虫氟苯双酰胺对草地贪夜蛾生物学特性的影响研究结果表明,亚致死剂量LD30处理能显著延长其生长发育历期,并导致蛹重、成虫重、幼虫存活率和单雌产卵量等生物学指标显著降低。这一结果与斜纹夜蛾Spodopteralitura(桑松等, 2014)、玉米螟Pyraustanubilalis(冯从经等, 2008)和棉铃虫Helicoverpaarmigera(宋永辉等, 2021)等害虫在化学杀虫剂亚致死剂量胁迫下的反应类似。如溴氰虫酰胺LC20和LC40剂量处理斜纹夜蛾3龄幼虫,其幼虫期延长,单雌产卵量显著降低;双氧威和虫酰肼亚致死剂量处理玉米螟,其幼虫体重及化蛹率同样明显降低;多杀霉素LC25处理棉铃虫幼虫,其幼虫期显著延长,蛹重和化蛹率均显著降低。
解毒酶活性增强可提高昆虫代谢杀虫剂的能力,保护酶活性的增强可提高昆虫对杀虫剂的防御能力,两者在昆虫耐药性和抗药性机制中的贡献均有大量的研究证实。研究表明,不同杀虫剂对草地贪夜蛾体内各解毒酶活性的诱导作用不同。蒋兴川等(2019)通过同样作用于GABA受体的甲维盐亚致死剂量(LD20)处理草地贪夜蛾72 h后,其体内EST酶活性明显高于对照组。王芹芹等人(2020)采用茚虫威亚致死剂量(LD20)处理草地贪夜蛾后,其体内MFO比活力显著高于对照组。李浩等(2021)发现,草地贪夜蛾3龄幼虫体内GST活性与氯虫苯甲酰胺亚致死剂量呈正相关,细胞色素单加氧酶CYP450与羧酸酯酶CarE活性则随处理剂量升高呈现先升高后下降的趋势。
溴虫氟苯双酰胺处理对不同生物体内解毒酶的诱导作用有所差异。齐浩亮等(2017)研究表明,亚致死剂量(LC20和LC40)的溴虫氟苯双酰胺可诱导小菜蛾Plutellaxylostella体内CarE活性升高,但对MFO和GST活性没有显著影响。本研究发现不同剂量溴虫氟苯双酰胺处理草地贪夜蛾后,其体内解毒酶GST和EST的比活力与药剂处理浓度均呈正相关,LD50胁迫下两者比活力最高。而解毒酶MFO的比活力以LD30处理组最高。LD50处理组的MFO比活力高于对照,但低于LD30处理,可能是由于溴虫氟苯双酰胺在草地贪夜蛾体内的积累抑制了MFO的活性,这与Zhan等人(2021)的研究结果类似。研究结果证明GST、MFO和EST可能都参与了草地贪夜蛾对溴虫氟苯双酰胺的解毒过程,且以GST发挥的作用最大。
外源毒物可使昆虫体内自由基(O2-、-OH和H2O2)的清除系统出现故障(Allenetal., 1989),对虫体产生毒害作用,而其体内保护酶(SOD、POD和CAT)可通过清除多余的活性氧自由基,使虫体不受外源有毒物质的侵害(Bashanetal., 2009)。害虫体内的保护酶对不同杀虫剂的反应程度不同。李浩等(2021)研究发现甲维盐处理草地贪夜蛾3龄幼虫后,其体内两种保护酶均呈现下降趋势,但氯虫苯甲酰胺处理后其体内SOD和POD均上升,推测以上两种酶可能是草地贪夜蛾对氯虫苯甲酰胺敏感性较甲维盐低的原因。本研究则发现亚致死剂量溴虫氟苯双酰胺胁迫下草地贪夜蛾体内保护酶SOD活性无明显变化,说明SOD在降低溴虫氟苯双酰胺对草地贪夜蛾的伤害中可能并未发挥作用;CAT和POD活性随处理剂量增加均明显上升,且两者在亚致死剂量LD30处理下的活性显著高于对照组,说明POD和CAT在降低机体受到溴虫氟苯双酰胺胁迫的氧化损伤中发挥了作用,但LD50处理组的活性与空白对照组无差异,可能是高剂量药剂处理对保护酶活性有明显抑制作用。因此,POD和CAT活性的增强也有可能成为将来草地贪夜蛾对溴虫氟苯双酰胺产生不同水平抗性的原因。
本研究通过生物学特性及酶活性测定,初步明确了溴虫氟苯双酰胺对草地贪夜蛾的亚致死效应,而草地贪夜蛾经溴虫氟苯双酰胺处理引起GST、MFO和EST活性增加的分子机制,及POD和CAT在该虫体内清除多余自由基的作用机制均有待进一步研究探明。
猜你喜欢
疯狂英语·新读写(2022年1期)2022-11-23
今日农业(2020年17期)2020-12-15
今日农业(2020年13期)2020-12-15
世界农药(2019年3期)2019-09-10
世界农药(2019年3期)2019-09-10
世界农药(2019年3期)2019-09-10
今日农业(2019年13期)2019-08-12
今日农业(2019年14期)2019-01-04
大灰狼(2016年8期)2016-09-06
火炸药学报(2014年3期)2014-03-20
