
虫害对华山松人工林土壤酶活性及碳氮磷化学计量特征的影响*
2023-11-05 12:59黄金莲崔鸿侠唐万鹏马致远雷静品
林业科学 2023年10期
黄金莲 崔鸿侠 唐万鹏 胡 琛 马致远 雷静品
(1. 中国林业科学研究院林业研究所 北京 100091;2. 湖北省林业科学研究院 武汉 430075)
近年来,随着极端气候事件频发,森林虫害发生频率和强度逐年增加,已成为影响森林稳定性的重要自然干扰事件(刘彦春等,2017),联合国粮农组织发布的全球森林资源评估报告数据显示,2017 年中国森林虫害发生面积比例高达4.2%(Food and Agriculture Organization of the United Nations,2020)。受虫害干扰后,林分结构和功能、林下植被、地表动物、土壤动物甚至微生物丰度及多样性等均发生显著变化,该变化会反馈于森林,导致森林生态系统的变化愈发复杂(袁杰等,2011)。Flower 等(2015)研究指出,以韧皮部为食的蛀干昆虫暴发会加剧树木死亡,降低森林生产力;Kurz 等(2008)研究发现,山松甲虫(Dendroctonus tankerosae)暴发导致北美西部森林由碳汇转为碳源;Moore 等(2013)研究认为,虫害暴发后土壤呼吸下降,几年后会有短暂恢复;Hicke 等(2012)研究发现,虫害发生后森林净初级生产力立即减少、水溶性有机物增多,森林生态系统的异养呼吸受到持续影响。C、N、P 是植物生长发育必不可少的营养元素(程瑞梅等,2018),对植物光合、呼吸、氮固定和有机物矿化等均有重要作用(Battermanet al.,2013;Piaszczyket al.,2019),同时C、N、P 的化学计量比是反映生态系统功能和生产力的重要指标(Panget al.,2020),常用于研究植物生长和营养限制之间的关系(Liuet al.,2020),可见对土壤中C、N、P 化学计量的研究意义重大。
土壤酶是森林土壤养分循环的关键(Luoet al.,2017),其中磷酸酶(acid phosphatase,ACP)、蛋白酶(acid protease,ACPT)、蔗糖酶(sucrase,SC)、过氧化氢酶(catalase,CAT)与森林土壤养分(C、N、P)循环密不可分。酶活性高低是水热因素和土壤养分共同作用的结果,反映微生物代谢及其对环境变化的响应,在土壤有机碳矿化(蔗糖酶)、N 循环(蛋白酶)、P 循环(磷酸酶)和酶促氧化还原反应(过氧化氢酶)中均具有重要作用(薛飞等,2020;Błónskaet al.,2021a;Małeket al.,2021)。Piaszczyk 等(2019)发现受虫害干扰后的华山松(Pinus armandii)生长受限,生物量减少,形成的枯立木通过释放养分和有机碳化合物形式进入土壤,会显著影响土壤碳平衡,参与养分循环的酶活性随之出现差异,从而对土壤呼吸速率产生影响(刘彦春等,2017)。
目前,已有森林土壤化学性质、土壤酶活性以及微生物的相关研究,主要探讨不同林分类型(喻林华等,2016)、经营模式和措施(丁思一等,2015;陈蕾等,2020)、外来植物入侵(侯玉平等,2015;张静等,2018)等背景下的土壤性质变化,以及虫害暴发后的土壤呼吸变化规律(刘彦春等,2017),虽然已有文献从碳循环角度分析了森林生产力变化并建立林分死亡率模型,但受虫害干扰后林分内部环境变化对碳排放的影响机制暂未明晰,充分认识虫害干扰对森林生态系统的影响,探索自然干扰下土壤酶活性、土壤化学性质以及土壤呼吸速率的变化规律,有利于合理经营森林(Attiwill,1994)。神农架人工林营造于20 世纪70 年代,华山松是主要造林树种之一,也是当地分布较广的优势树种。本研究测定神农架林区受虫害干扰后华山松林生长季内的土壤酶活性、土壤化学性质和土壤呼吸速率,探究虫害干扰的影响,以期为受虫害华山松林的土壤碳排放管理提供理论依据。
1 研究区概况与研究方法
1.1 研究区概况及样地设置
研究区设在湖北神农架国家公园(110°03′05′′—110°33′50′′E,31°21′20′′—31°36′20′N),公园位于湖北省西北部的房县、巴东、兴山三县交界地带,海拔398~3 106.2 m。属北亚热带季风气候区,是北亚热带向暖温带过渡区域,年降水量800~2 500 mm,全年日照时数1 858 h,年均日照率42%。沿海拔由低到高依次分布有常绿阔叶林、常绿落叶阔叶混交林、针阔混交林、针叶林、灌丛和草甸(沈泽昊等,2004)。样地内土壤具垂直地带性,随海拔升高依次为山地黄棕壤、山地棕壤、山地暗棕壤、山地棕色针叶林土、山地草甸土。林区动植物资源极其丰富,其中国家重点保护植物76 种、昆虫约28 目157 科4 143 种,分布有水杉(Metasequoia glyptostroboides)纯林、华山松纯林,还有与山杨(Populus davidiana)、 糙皮桦(Betula albosinensis)、锐齿槲栎(Quercus alienavar.acutiserrata)等落叶阔叶树种形成的针阔混交林(菊花等,2016)。
本研究在湖北神农架森林生态系统国家定位观测研究站试验地进行。2021 年3 月,在华山松林大小蠹(Dendroctonus armandi)虫害主要发生区,选取2017—2018 年受虫害干扰后土壤类型相同,海拔、坡向、林龄、树种组成等相近的样地,林木存活率约11%(虫害干扰后无人为干扰和采伐活动,多数华山松呈分解程度不同的枯立木),同时选取未受虫害干扰的健康华山松林作为对照;分别随机选取3 块20 m×20 m 的样地(表1)。

表1 华山松林样地基本情况Tab. 1 The setting of plots of P. armandii forest stands
1.2 土壤化学性质测定
2021 年7 月,利用土钻法在各样地6 个土壤呼吸环附近钻取0~10 cm 土层土壤混合样品,各3 个重复,土样自然风干后剔除其中的石砾、根系和凋落物等,过0.15 mm 筛后分袋放置。测定土壤pH 和有机碳(soil organic carbon,SOC)、全氮(total nitrogen,TN)、全磷(total phosphorus,TP)、全钾(total potassium,TK)、水解性氮(available nitrogen,AN)、有效磷(available phosphorus,AP)、速效钾(available potassium,AK)含量。pH 测定采用水浸法,SOC 含量测定采用硫酸重铬酸钾法,TN 含量测定采用凯氏定氮法,TP 含量测定采用钼锑抗比色法,TK 含量测定采用火焰光度计法,AN 含量测定采用碱解扩散法,AP 含量测定采用氢氧化钠浸提钼锑抗比色法,AK 含量测定采用乙酸铵浸提-火焰光度计法(鲍士旦,2000)。
1.3 酶活性测定
土壤酶活性测定磷酸酶(ACP)、蛋白酶(ACPT)、蔗糖酶(SC)和过氧化氢酶(CAT)4 个指标。ACP 活性采用磷酸苯二钠比色法(土壤为酸性,测定的是酸性磷酸酶)测定;ACPT 活性采用茚三酮比色法测定;SC 活性采用3,5-二硝基水杨酸比色法测定;CAT 活性采用KMnO4滴定法测定(关松荫,1986)。购买相应的酶活性测定试剂盒,根据酶种类添加对应的提取液和缓冲液,分步骤将药品加入、振荡、静置、水浴加热和移液器操作后采用多功能酶标仪测定荧光度(水解酶)或吸光度(氧化酶),基于酶活性浓度测定公式计算最终浓度,每份样品重复测定3 次。
1.4 土壤呼吸测定
2021 年3 月,在各样地沿对角线安装3 个PVC土壤呼吸环,整个研究过程中呼吸环位置保持不变。2021 年4—12 月,每月选择晴朗日的9:00—17:00,用美国 LI-Cor 公司生产的 LI-8100 便携式土壤碳通量测量系统测定土壤呼吸速率。土壤呼吸环内径20 cm、外径21 cm、高度8 cm,将其一端削尖沿样地坡面插入地下5 cm,每月测量前齐地剪除环内植物,避免扰动土壤。每个土壤呼吸环每次测定2 次,取平均值。
1.5 数据处理与分析
采用单因素方差分析(one-way ANOVA)对华山松林的土壤酶活性、化学性质和生态化学计量比进行显著性分析;采用Canoco 5.0 软件对酶活性与化学计量比进行主成分分析,筛选主要影响因子;利用Canoco 5.0 软件进行冗余分析,比较土壤化学性质和生态化学计量比对酶活性的影响;采用Person 法进行土壤酶、化学性质与土壤呼吸的相关性分析。应用Excel 2021 和SPSS 25.0 软件对试验数据进行大致梳理和分析,利用Origin 2021、Excel 2021 软件完成制图和绘图。
2 结果与分析
2.1 干扰对土壤化学性质的影响
2 种华山松林样地部分土壤化学性质存在显著差异(表2)。受虫害和健康华山松林土壤平均C、N、P含量分别为50.71 和53.54 g·kg-1、2.48 和2.57 g·kg-1、1.22 和0.60 g·kg-1。受虫害华山松林土壤pH、SOC、TN、AN、AK 含量和C/N 与健康华山松林无显著性差异,但C/P、N/P 均显著低于健康华山松林(P<0.05),土壤TP、TK、AP 含量均显著高于健康华山松林(P<0.05),其中受虫害华山松林土壤TP 含量约为健康华山松林的2 倍,AP 含量高出3 倍。
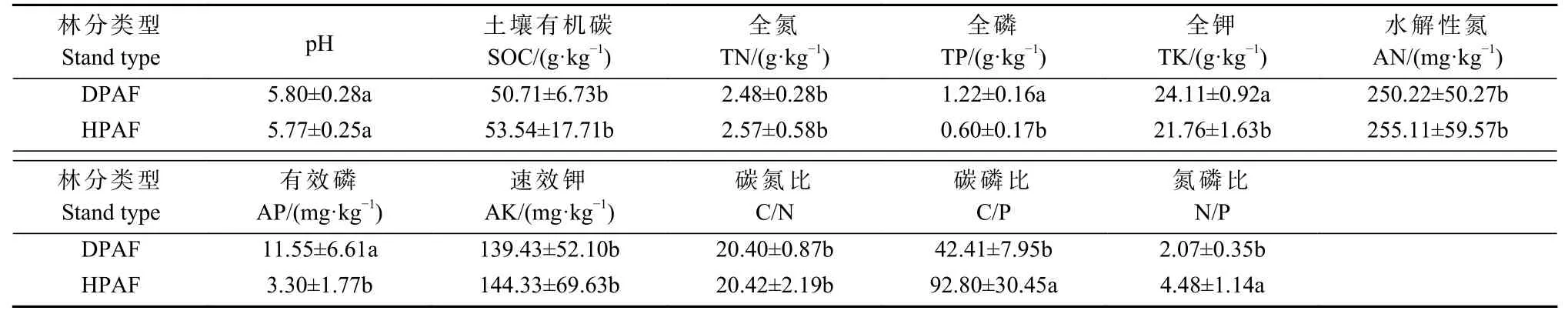
表2 华山松林土壤化学性质特征(平均值±标准误差)①Tab. 2 Characteristics of soil chemical properties of P. armandii forest(mean±SE)
2.2 干扰对土壤酶活性的影响
由图1 可知,受虫害华山松林土壤CAT、ACPT、SC 酶活性均显著高于健康华山松林(P<0.05),其中受虫害华山松林相比健康华山松林CAT 活性升高22.26%、ACPT 活性升高39.3%、SC 活性升高135.38%;而ACP 酶活性显著低于健康华山松林(P<0.05),相比健康华山松林ACP 活性下降12.20%。
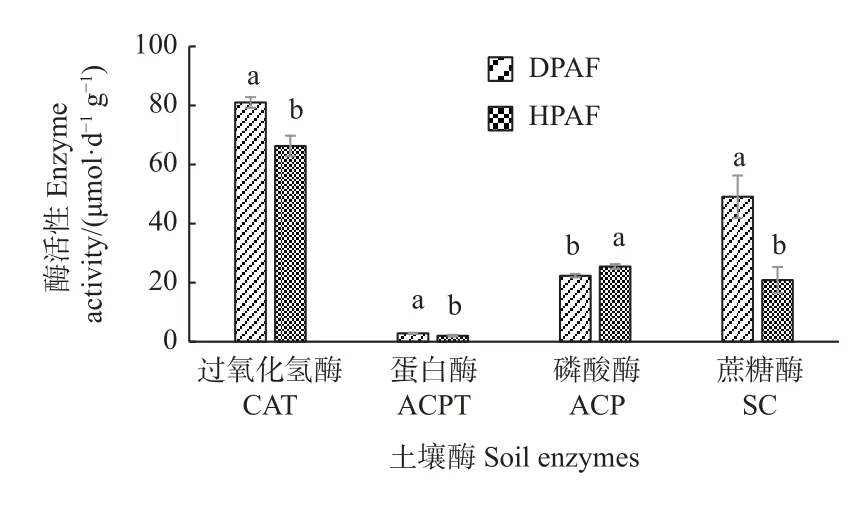
图1 华山松林土壤酶活性变化Fig. 1 Variation of soil enzyme activity in P. armandii forests
2.3 干扰后土壤酶与化学性质分析
由图2 可知,PCA1 轴可解释2 种华山松林土壤化学因子与酶活性差异的62.48%,PCA2 轴可解释2种华山松林土壤化学因子与酶活性差异的15.76%,说明受虫害华山松林与健康华山松林沿PCA1 轴具有明显差异,受虫害华山松林土壤样本与健康华山松林土壤样本在坐标系内分布具有明显差异,说明虫害干扰使土壤在化学性质和酶活性方面与健康华山松土壤发生一定变化。
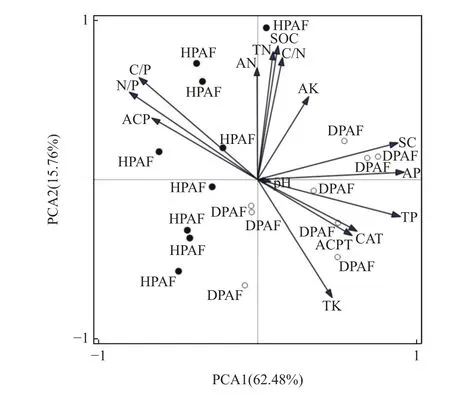
图2 华山松林土壤酶活性与化学性质的主成分分析Fig. 2 Principal component analysis of soil enzyme activity and chemical properties in P. armandii forest
在土壤酶与化学性质的冗余分析(图3)中,空心箭头代表11 个土壤化学因子,实心箭头代表4 种土壤酶活性,各指标的贡献率以线条长度显示,线条越长表明贡献率越大,反之越小。箭头间夹角余弦值表示土壤化学因子与酶活性的相关性,夹角越小表明二者相关性越强,夹角大于90°表明二者间存在负相关。TP、AK、TK、AP、N/P、C/P、SOC、TN 的箭头连线相对较长,说明对4 种土壤酶活性具有较强影响。AK、SOC、TN、C/N、AP、TP、TK 与SC、ACPT、CAT 活性方向一致,表明上述土壤化学因子与SC、ACPT、CAT 活性呈正相关;与ACP 活性方向相反,表明与ACP 活性呈负相关。N/P、C/P、AN、pH 与ACP 活性方向一致即呈正相关关系,与SC、ACPT、CAT 活性方向相反即呈负相关关系。AP 与SC、TK 与ACPT和CAT、N/P 与ACP 夹角较小,呈现较显著正效应,说明N/P 是影响ACP 活性的主导因子,TK 是影响ACPT 和CAT 活性的主导因子,4 种酶活性分别由不同土壤化学因子主导。AP、TP 均对CAT、ACPT、SC活性具有正相关关系,是三者的共同影响因子。
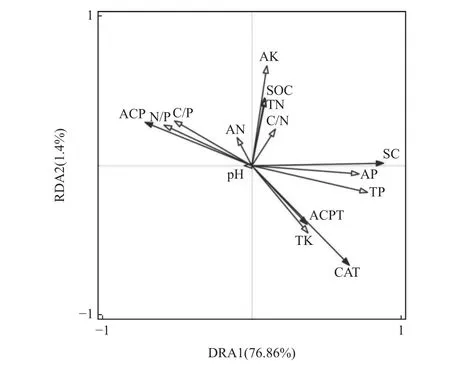
图3 华山松林土壤酶活性与化学性质的冗余分析Fig. 3 Redundant analysis of soil enzyme activity and chemical properties in P. armandii forest
华山松林土壤酶活性与土壤化学性质的相关性分析热图(图4)显示,TP、AP、C/P、N/P 与4 种土壤酶活性呈显著性差异:TP、AP 与CAT、ACPT、SC 活性显著正相关,与ACP 活性显著负相关;C/P、N/P 与CAT、ACPT、SC 活性显著负相关,与ACP 活性显著正相关。4 种酶活性之间均呈显著相关,生态化学计量比(C/P、N/P)与土壤酶(CAT、ACPT、SC)活性显著负相关,与磷酸酶(ACP)活性显著正相关。
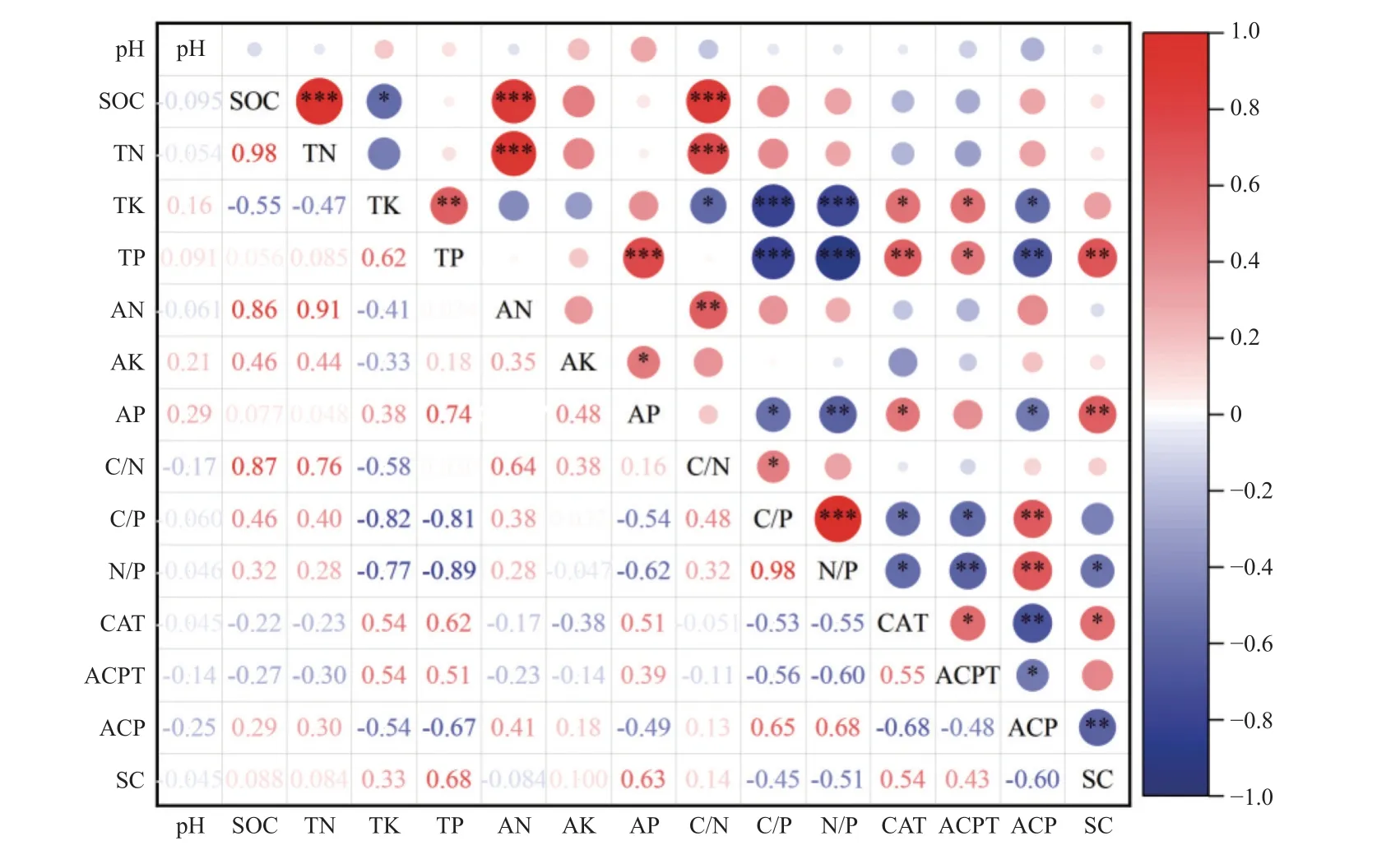
图4 华山松林土壤酶活性与化学性质的相关性分析热图Fig. 4 Heat map of correlation between soil enzyme activity and chemical properties in P. armandii forest*P≤0.05, **P≤0.01, ***P≤0.001.
将土壤pH 和SOC、TN、TP、TK、AN、AP、AK含量以及C/N、C/P、N/P 等11 个土壤化学因子与4 种酶活性进行冗余分析,第一轴、第二轴特征值分别为0.768 6 和0.014 0,土壤酶与化学因子的相关性分别为0.887 8 和0.782 0(表3)。根据排序结果,前2 轴11 个土壤化学因子对酶活性累计解释量达78.26%,对土壤酶-化学因子累计解释量达99.81%,总体来看排序结果较好,前2 轴在很大程度上可解释土壤酶与化学因子间的关系变化,第一轴对排序结果起决定性作用。
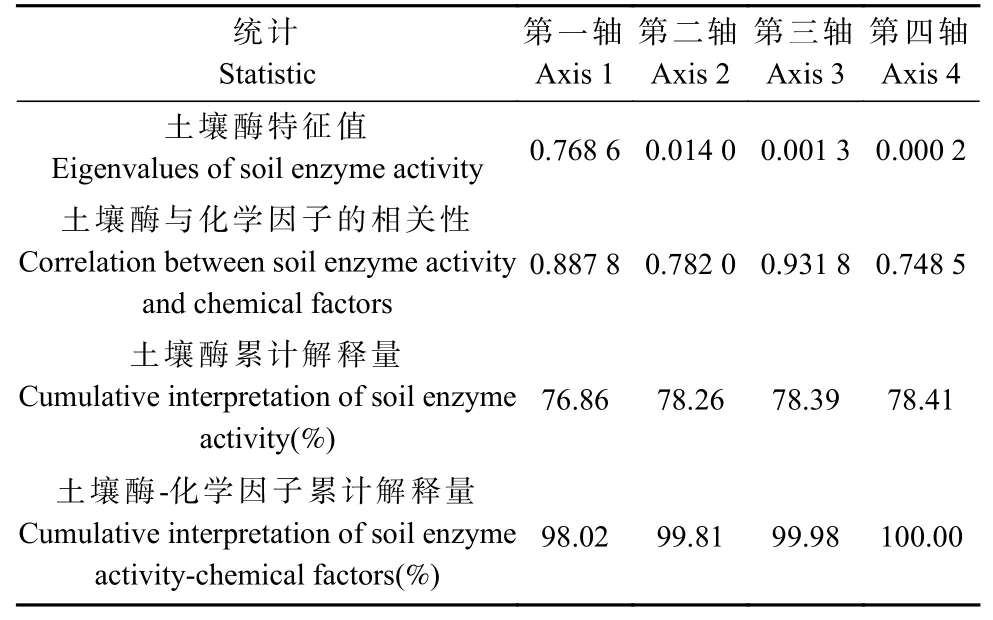
表3 土壤酶活性与化学因子冗余分析Tab. 3 Redundancy analysis of soil enzyme activity and chemical factors
对11 个土壤化学因子进行蒙特卡洛置换检验,土壤化学因子对酶活性的重要性排序结果为TP>AP>N/P>C/P>TK>C/N>AK>SOC>TN>AN>pH(表4)。TP、AP 对土壤酶活性的影响达极显著水平(P<0.01),解释量分别为58.9%、50.5%;N/P、C/P 对土壤酶活性的影响达显著水平(P<0.05),解释量分别为34.0%、26.5%;其余7 个化学因子对土壤酶活性的影响并不显著(P>0.05),贡献率也相对较低。
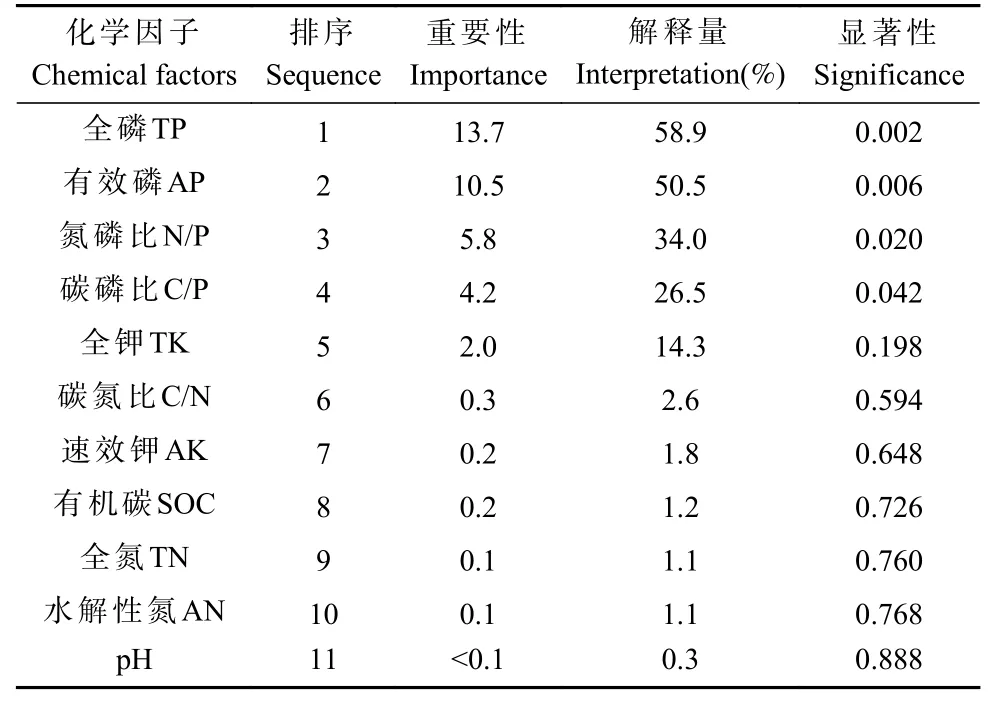
表4 化学因子蒙特卡洛置换检验结果分析Tab. 4 Analysis of Monte Carlo displacement test results for chemical factor
2.4 土壤呼吸变化
受虫害和健康华山松林土壤呼吸速率在生长季内各月变化趋势基本一致,均呈单峰曲线特征(图5)。从4 月到7 月,健康华山松林土壤呼吸速率随土壤温度升高而上升,7 月达最高值3.28 µmol·m-2s-1;受虫害华山松林土壤呼吸速率5 月轻微降至1.52 µmol·m-2s-1,之后随土壤温度升高而上升,也在7 月达最高值2.81 µmol·m-2s-1;从7 月到12 月,随土壤温度下降,受虫害和健康华山松林土壤呼吸速率均逐渐降低,且健康华山松林土壤呼吸均高于受虫害华山松林,二者在12 月达最小值,分别为0.48 和0.51 µmol·m-2s-1。

图5 土壤呼吸速率月变化Fig. 5 Monthly variations of soil respiration rate
方差分析(图5)表明,不同月份之间土壤呼吸速率达极显著差异(P<0.01)。土壤呼吸速率也存在季节差异,春(3—5 月)、夏(6—8 月)、秋(9—11 月)、冬季(12—2 月)的平均土壤呼吸速率分别为1.58、2.57、1.47、0.48 µmol·m-2s-1,夏季土壤呼吸速率显著高于春、秋、冬季。受虫害华山松林土壤呼吸速率在生长季内均低于健康华山松林,且差异集中体现在夏季生长旺期,说明夏季高温对健康华山松林土壤呼吸影响更大,促进健康林碳排放。
2.5 土壤酶活性和化学性质与土壤呼吸速率的关系
受虫害华山松林、健康华山松林土壤酶活性和化学性质与土壤呼吸速率的Person 相关分析(表5)表明,受虫害华山松林土壤呼吸速率与11 种土壤化学因子和4 种土壤酶活性之间均无显著相关,而健康华山松林土壤呼吸速率与pH 呈显著负相关(P<0.05),与TP、AK、AP 呈显著正相关(P<0.05),与其他化学因子和酶活性无显著相关。当不区别受虫害或健康森林类型时土壤呼吸速率与pH 呈显著负相关(P<0.05),与AK 呈显著正相关(P<0.05)。受虫害和健康华山松林中,土壤呼吸速率与pH、TK、C/P、N/P 均呈负相关,与TP、AK、AP、C/N、SC 呈正相关。受虫害华山松林土壤呼吸速率与SOC、TN、AN、ACP 呈负相关,与CAT、ACPT 呈正相关,健康华山松林则与之相反。
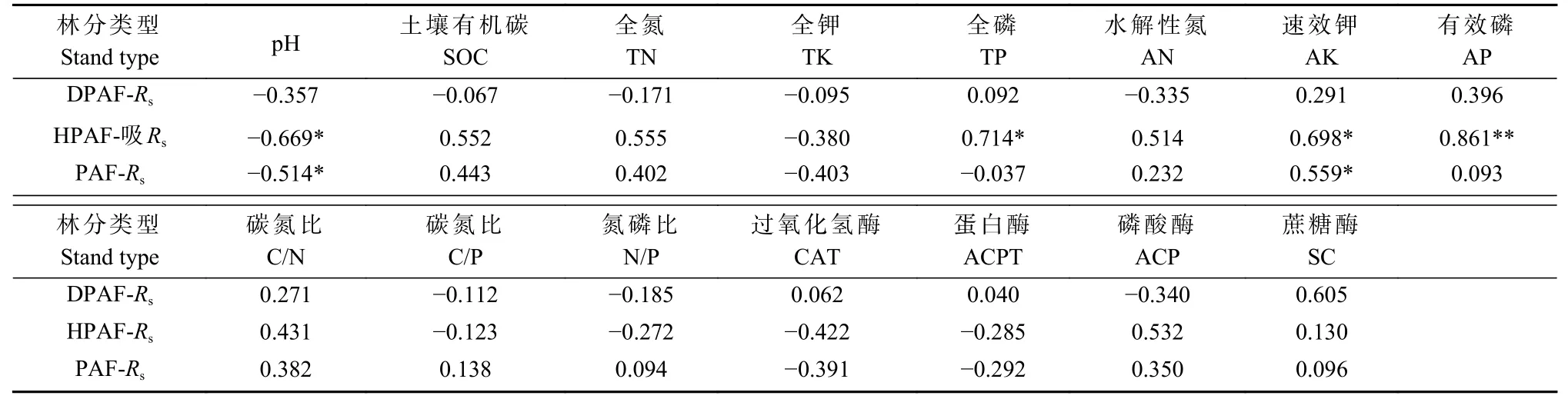
表5 华山松林土壤酶活性、化学性质与土壤呼吸相关分析①Tab. 5 Soil enzyme activity, chemical properties and soil respiration correlation analysis in P. armandii forest
土壤呼吸在受虫害和健康华山松林中受不同因子影响,进一步分析显著影响因子与土壤呼吸速率的线性回归关系(表6)表明,受虫害华山松林土壤呼吸速率与各化学因子和土壤酶活性之间均无显著回归关系,但健康华山松林土壤呼吸速率与土壤pH 和TP、AK、AP 含量存在显著多元线性回归关系,确定系数为0.827(其中与pH 关系显著,P<0.05)。

表6 土壤呼吸与影响因子回归分析Tab. 6 The correlation between soil respiration and the quantity of standing trees
3 讨论
3.1 干扰后华山松林土壤养分特征
本研究发现,受虫害干扰后华山松林土壤化学性质发生显著变化,11 个土壤化学因子中有8 个(pH 和SOC、TN、AN、AK 含量和C/N、C/P、N/P)均低于健康华山松林,其中C/P、N/P 显著低于健康华山松林(P<0.05),而TP、TK、AP 含量显著高于健康华山松林(P<0.05)。各元素含量及其化学计量比存在一定差异,可能是虫害干扰导致林木死亡,改变了植物根系与土壤营养元素的交换量。土壤中N 元素主要来源于生物固氮和大气沉降,土壤中P 和K 元素主要来源于凋落物分解作用(陈蕾等,2020),华山松林受虫害干扰后树木死亡凋落物增加,凋落物在微生物活动下分解,增加土壤中P 和K 含量。Morehouse 等(2008)研究发现受干扰地块中80%的西黄松(Pinus ponderosa)受小蠹虫侵袭,而受干扰林与健康林具有相同的地上生物量,其中受干扰林60%的生物量存贮于死树,而健康林仅有5%的生物量存贮于枯立木,导致受虫害干扰林的土壤C/N 发生变化(受干扰林C/N 比低于健康林),与本研究结果一致,说明虫害干扰导致C 大部分储藏在枯立木中,对土壤的C 输入减少,导致土壤C/N 发生改变。高瑞贺等(2015)研究发现虫害干扰后林下植被多样性与乔木层树种重要值呈显著负相关,说明林地受虫害干扰后,优势树种衰退且林下植被存活率和多样性均增加,形成的2 类新林分对土壤养分吸收出现差异,从而影响土壤的C、N、P 含量及化学计量特征出现差异。王绍强等(2008)研究得出,土壤C/N、C/P 越低时,土壤N、P 的有效性越高。受虫害华山松林土壤C/N 低于健康华山松林,二者相差不大,而健康华山松林土壤C/P 显著高于受虫害华山松林,受虫害华山松林土壤C/P 未达到全球同层次土壤C/P(Clevelandet al., 2007),说明受干扰林不存在磷限制(Huanget al.,2018),磷含量显著增加,C/P 显著降低。
3.2 干扰后华山松林土壤酶活性及其养分限制因子
虫害干扰显著影响土壤酶活性。SC 活性取决于土壤有机质含量和C/N 高低,ACP 活性与土壤有机磷的分解转化有关,CAT 与土壤有机质的转化速度密切相关(任玉连等,2020)。本研究中,受虫害华山松林0~10 cm 土壤CAT、ACPT、SC 活性显著高于健康华山松林(P<0.05),而CAT、ACPT 和SC 活性升高使土壤中易溶性物质减少,植物氮源水解产物减少,氮矿化能力下降;土壤ACP 活性显著低于健康华山松林(P<0.05),会导致土壤中可供植物吸收的无机磷含量变少。与健康林相比,受干扰林的林下植被在生长季快速生长,增强了对土壤中营养元素的吸收和积累,之后再以有机物形式归还给土壤(库)(Zhaoet al.,2013),在该过程中土壤微生物活动增强,进而导致土壤酶活性升高。已有研究表明,虫害干扰也会导致林分土壤含水量增加(高瑞贺等,2015),土壤含水量增加会直接影响土壤酶促反应强度,从而间接影响酶活性高低(解丽娜等,2014;Jinet al.,2009)。本研究发现,受干扰华山松林土壤ACP 活性显著低于健康华山松林,ACP 活性下降会降低土壤无机磷的转化率和植物对无机磷的吸收。C/P、N/P 与CAT、ACPT、SC 活性呈显著负相关,与ACP 活性呈显著正相关,表明4 种土壤酶均受土壤中P 含量的影响。虫害发生后,华山松林P 循环发生变化,进而导致受干扰林与健康林土壤酶活性出现差异。有研究发现,C/P 越低时,土壤P有效性越高(王绍强等,2008),ACP 活性越高,与本研究结果相符,表明P 可能是驱动土壤生态酶活性及土壤化学因子计量比内在联系的关键因子(刘仁等,2020)。还有研究得出,酶活性高低由凋落物分解过程中释放的C 底物决定(Błónskaet al.,2021b),因此除本研究中与P 相关的生态化学计量比是土壤酶活性的影响因子外,在之后研究中还需考虑凋落物分解速率及其养分归还过程中碳氮养分的富集效应。
土壤化学性质改变会引起土壤微生物群落结构发生相应变化,从而间接影响土壤酶活性;土壤磷和钾能促进植物根系生长,增强土壤微生物及根系分泌酶的能力,说明土壤TK、TP 含量增加也会影响ACT、ACP 和SC 活性(任玉连等,2020)。虫害干扰发生后,凋落物一次性掉落堆积在地面,会使土壤微生物群落结构也发生一定变化,导致枯落物分解速率下降(Štursováet al.,2014)。由此可见土壤微生物对虫害干扰的响应是较明显的,但本研究仅考虑土壤养分和酶活性变化,并未测定土壤微生物活性如何,未来有待将微生物指标纳入比较,以便更系统地探究其差异缘由。
3.3 干扰后华山松林土壤呼吸速率变化及其与土壤环境的关系
受虫害华山松林在生长季内的土壤呼吸速率均低于健康华山松林,且差异集中体现在夏季生长旺期,说明夏季高温对健康华山松林土壤呼吸影响更大,促进健康林碳排放。可能的原因是虫害干扰会改变华山松林地环境,如光照强度、微生物群落结构、酶活性、林下植被组成等,林下光照强度增强导致地被植物快速生长,缓解了受干扰森林生态系统的部分C 损失(Zehetgruberet al.,2017),进而对土壤呼吸进行调控。本研究中受虫害华山松林土壤呼吸速率与11 种土壤化学因子和4 种土壤酶活性无显著相关性,而健康华山松林土壤呼吸速率与ACP 具有显著相关,同时土壤呼吸速率与P 的流动途径也相关联;将土壤呼吸速率与2 种林分的土壤化学性质和土壤酶活性变化结合起来分析表明,造成土壤呼吸速率差异的原因是土壤化学性质和酶活性变化,而Štursová等(2014)研究表明土壤呼吸速率与凋落物分解、植物根系的衍生及土壤中微生物群落结构有关。类似虫害干扰后,森林异养呼吸的增加基本上被自养呼吸下降所抵消,但有时总体呼吸作用仍下降,很大程度上是因为虫害干扰导致大量林木死亡,植物呼吸作用大幅降低(Schimelet al.,2003)。未来可能需要在土壤有机碳循环与微生物过程之间建立明确联系的模型,才能有效模拟土壤碳释放(Zobitzet al.,2008;Allisonet al.,2010)。Lybrand 等(2018)研究发现蛀干害虫钻进植物韧皮部时,会破坏植物糖运输过程,导致林木死亡,之后土壤中可提取的可溶性碳和氮、无机磷含量和微生物生物量(碳和氮)开始下降,但在树木死亡后的4~5 年间趋于增加,说明受虫害干扰的森林是个长期变化过程,在干扰后不同时期土壤养分和微生物生物量是不同的。
森林中普遍存在的低温以及底物限制也会影响枯落物分解,虫害暴发后,如果没有火的迅速释放,碳可能会在森林生态系统中长期保存(Kulakowskiet al.,2011)。Moore 等(2013)将控制试验与长期监测相结合,阐明昆虫干扰后森林通过降低枯落物分解速率来控制释放二氧化碳量。除了研究干扰如何影响土壤微气候外,可能还需要对微生物呼吸进行更全面探索,才能更有效模拟虫害干扰导致的树木死亡对碳平衡的影响。受干扰华山松林土壤呼吸速率与化学因子和土壤酶活性之间均无显著相关,但健康华山松林土壤呼吸速率具有与pH、TP、AK、AP 含量的显著多元线性回归关系,其中与pH 呈显著相关(P<0.05)。在生态系统尺度上,土壤pH 能反映外界环境变化对土壤风化和植物群落组成的控制(Lasotaet al.,2021),主要是通过对土壤养分以及微生物群落的调控进而影响土壤呼吸速率。
4 结论
1) 虫害干扰后,华山松林木大量死亡,降低了对土壤养分的吸收及转化,导致土壤大部分养分含量相对降低,但TP、TK 及AP 含量显著上升(P<0.05)。
2) 华山松死亡后,降低了根系的分泌活动,导致4 种土壤酶活性在虫害干扰后均发生显著性变化,其中CAT、ACPT 和SC 活性显著上升,而ACP 活性显著降低(P<0.05)。
3) 虫害发生后土壤呼吸速率显著降低,但2 种林分的土壤呼吸季节性变化趋势大致相同,土壤呼吸速率随SOC、TN、AN、ACP 含量上升而下降,呈负相关关系。
猜你喜欢
Plasma Science and Technology(2022年9期)2022-08-29
小学生学习指导(中年级)(2021年3期)2021-04-06
河北果树(2020年4期)2020-11-26
少儿美术(快乐历史地理)(2020年12期)2020-06-09
今日农业(2019年13期)2019-08-12
现代园艺(2017年22期)2018-01-19
现代园艺(2017年19期)2018-01-19
连环画报(2016年10期)2016-12-16
心潮诗词评论(2016年1期)2016-12-01
