
不同砧穗组合楸树嫁接苗的生理生化特性*
2023-11-05 12:59:12贾瑞瑞祝艳艳王良桂杨秀莲
林业科学 2023年10期
何 荷 贾瑞瑞 付 钰 祝艳艳 王良桂 杨秀莲
(南京林业大学风景园林学院 南京 210037)
楸树(Catalpa bungei)系紫葳科(Bignoniaceae)梓属(Catalpa)落叶乔木,是我国特有的用材树种和园林观赏树种(陈慧玲等,2021)。由于楸树自花不育、异花授粉结实率低、种子发芽率低,不易有性繁殖,而无性繁殖不仅生产成本高、苗木早期生长适应性弱而且品种不同生根率差异也很大,因此,生产上多采用嫁接繁殖法实现楸树苗木的快速扩繁(张烨然等,2016;Melnyket al.,2015;彭方仁等,2022)。在苗木生长过程中发现,南方部分地区的嫁接苗容易出现砧木接穗接口部位生长不协调、嫁接部位产生离层、根结线虫危害严重等问题(王改萍等,2020),这说明楸树嫁接苗的亲和性较低,而亲和性和接穗砧木种类密切相关。楸树最适宜的嫁接砧穗组合的研究还较少,仅从嫁接组合的生长状况、材性性状、光合生理等方面进行了探讨(王军辉等,2013;李凤等,2022),灰楸(Catalpa fargesii)的生长状况和光合特性受砧木种类的影响较大,梓树(Catalpa ovata)最适宜作为灰楸的砧木;灰楸和楸树相比于黄金树(Catalpaspeciosa)更适合作为楸树的砧木,灰砧和楸砧嫁接苗的生长状况、材性性状、保存率、接穗生根率都较好。已有研究表明嫁接苗的亲和性强弱和生长状况在一定程度上与其营养物质含量、防御酶活性、酚类物质含量等的生理变化关系密切(Lopez-Gomezet al.,2007; 王瑞等,2014),但生理生化特性和综合评价还未见报道。因此,本研究选择梓树和滇楸(Catalpafargesiif.duclouxii)作为砧木,‘南林1 号’(Catalpa bungei‘Nanlin No.1’)、 ‘洛楸’(Catalpa bungei‘Luoqiu’)和‘苏楸1 号’(Catalpa bungei‘Suqiu No.1’)这3 种优良的主栽楸树品种作为接穗,比较不同砧穗组合间渗透调节物质、抗氧化酶活性、内源激素含量等生理指标的差异,以期筛选出适宜的楸树嫁接砧穗组合以及可反映嫁接植株亲和性和生长状况的主要生理指标。
1 材料与方法
1.1 试验材料与设计
试验于2020 年4 月中旬在江苏省南京市溧水区白马镇南京林业大学教学科研基地(31°58′36″N,119°18′45″E)进行。砧木为1 年生梓树(Z)和滇楸(D)的实生苗,粗度约为1 cm,长度约为18 cm;接穗为楸树的3 个优良无性系的饱满单芽,分别是‘南林1 号’(N)、‘洛楸’(L)和‘苏楸1 号’(S),共6 个组合(表1)。每个组合嫁接200 株,株间距为0.6 m×1.5 m,嫁接后对不同砧穗组合进行施肥、灌溉和除草等统一常规田间管理。

表1 楸树嫁接组合Tab. 1 Grafting combination of C. bungei
1.2 测定指标及方法
规律间隔的连续取样有助于分析判断嫁接苗的生理物质变化趋势及其生长发育状况。由于本研究采用的是芽接,嫁接早期砧穗处于缓慢愈合并逐步融合的过程,此期取样不能充分反映嫁接苗的生长状况,故取样时间确定为嫁接4 个月待新梢停止生长后进行。具体时间根据当时的天气状况进行了调整,最终为8 月12 日、8 月23 日、9 月5 日、9 月19 日、10 月8 日和10 月23 日。取样部位为各嫁接组合相同部位的成熟叶片,每个组合各取12 株,每4 株作为1 个平行样,重复3 次,叶片做好标记后置于冰盒,带回实验室后立即置于-80 ℃超低温冰箱保存备用。
1.2.1 成活率 于嫁接6 个月后,统计各楸树嫁接组合的成活率,计算公式为:SR = NSP/TNGP×100%。式中:SR 为存活率(survival rate);NSP 为成活株数(number of surviving plants);TNGP 为嫁接总株数(total number of grafted plants)。
1.2.2 渗透调节物质含量 采用蒽酮比色法测定可溶性糖和淀粉含量,采用考马斯亮蓝G-250 染色法测定可溶性蛋白含量(王学奎等,2015)。
1.2.3 抗氧化酶活性 超氧化物歧化酶(SOD)活性的测定采用氮蓝四唑(NBT)光还原法,过氧化物酶(POD)活性的测定采用愈创木酚法,过氧化氢酶(CAT)活性的测定采用紫外吸收法,多酚氧化酶(PPO)活性的测定采用邻苯二酚法(王学奎等,2015;张晓等,2008)。
1.2.4 内源激素含量 采用酶联免疫法(王学奎等,2015)测定生长素(IAA)、脱落酸(ABA)、赤霉素(GA3)和玉米素核苷(ZR)含量。从-80 ℃超低温冰箱中取出叶片,于液氮中研磨至粉末状,每个样品准确称量0.6 g,加入2 mL 样品提取缓冲液置4 ℃冰箱充分提取4 h;4 ℃,4 000 r·min-1离心15 min,吸取上清液,向剩余沉淀加入1 mL 提取液后重复上述步骤,样品提取液为2 次上清液之和。之后将提取液置于C18 固相萃取预处理小柱中萃取并收集得到粗提液,将粗提液置于离心浓缩机进行浓缩,再加入0.9 mL 样品稀释液进行定容,之后使用ELISA 试剂盒采用酶联免疫法测定内源激素含量。
1.3 数据处理
使用Microsoft Excel 2010 进行数据处理,利用SPSS 26.0 软件的单因素方差分析法(one-way ANOVA)进行不同楸树嫁接组合间的差异显著检验(P<0.05),使用Origin 2018 软件绘制多因子柱状图、相关性热图;利用SIMCA 14.1 软件和SPSS 26.0 软件的主成分分析(principal component analysis,PCA)法和正交偏最小二乘判别分析法(orthogonal partial least squaresdiscriminant analysis,OPLS-DA)进行各楸树嫁接组合间的多重比较和主要生理指标筛选。
2 结果与分析
2.1 不同砧穗组合楸树嫁接苗的成活率
由表2 可知,各嫁接组合间成活率有较大差异,其中DN 组合成活率最高,为76.91%;DS 组合成活率最低,仅为32.05%。相同接穗不同砧木的嫁接组合,DN 比ZN 成活率高17.83%;ZS 比DS 成活率高28.9%;而ZL 和DL 组合的成活率无显著差异。相同砧木不同接穗组合,在梓砧嫁接组合中,ZL 的成活率显著高于ZN 和ZS;在滇砧嫁接组合中,DN 的成活率显著高于DL,DL 显著高于DS。
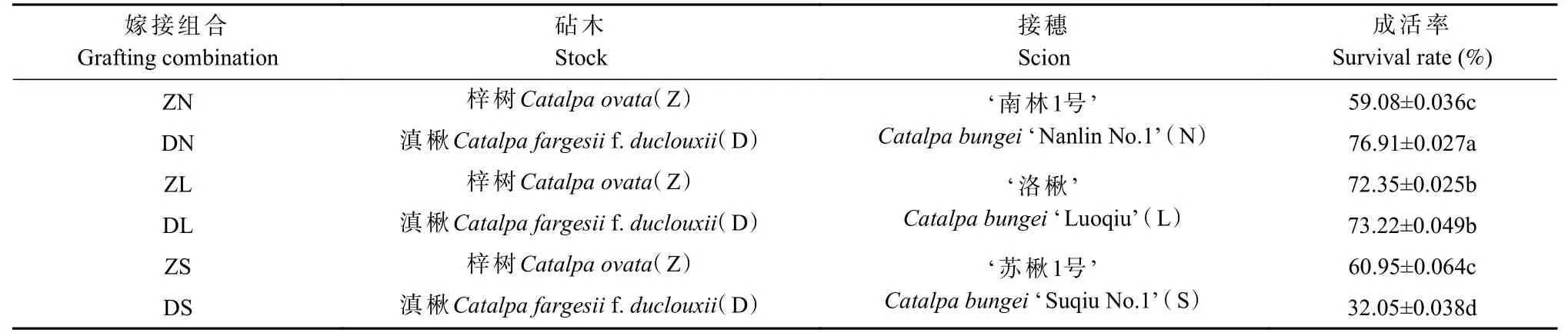
表2 不同楸树砧穗组合的成活率①Tab. 2 Survival rate of different rootstock panicle combinations of C. bungei
2.2 不同砧穗组合楸树嫁接苗渗透调节物质变化
2.2.1 可溶性糖含量和淀粉含量变化 由图1、图2可知,除ZS 组合的可溶性糖含量呈先升后降趋势外,各嫁接组合的可溶性糖含量和淀粉含量变化一致,均呈现先降后升再降的趋势,且均在9 月份有最大值。相同接穗不同砧木组合,多数时期DN 组合、DL 组合的可溶性糖和淀粉含量分别显著大于ZN 组合、ZL组合,而多数时期DS 组合的可溶性糖和淀粉含量均显著小于ZS 组合。相同砧木不同接穗组合,在梓砧嫁接组合中,9 月5 日可溶性糖含量差异显著,由高到低依次为ZS 组合、ZL 组合、ZN 组合,在测量前期,ZS 组合的淀粉含量在不同程度上大于ZN 和ZL 组合;在滇砧嫁接组合中,多数时期可溶性糖含量无显著差异,多数时期DN 组合的淀粉含量最高,而DS 组合最低。
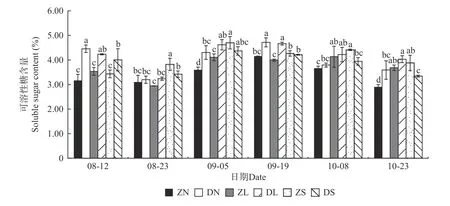
图1 不同楸树砧穗组合可溶性糖含量变化Fig. 1 Variation of soluble sugar content in different rootstock panicle combinations of C. bungei
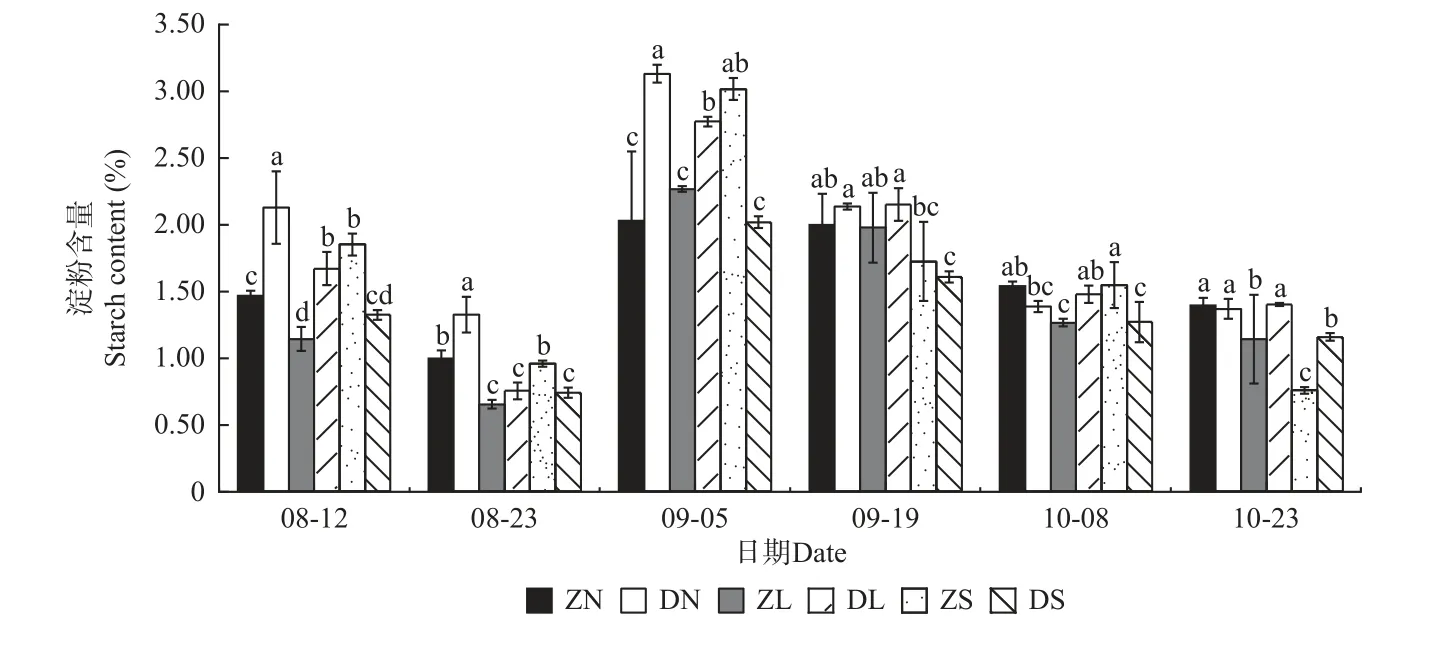
图2 不同楸树砧穗组合淀粉含量变化Fig. 2 Variation of starch content in different rootstock panicle combinations of C. bungei
2.2.2 可溶性蛋白含量变化 由图3 可知,各嫁接组合的可溶性蛋白含量大致呈先上升后下降的趋势,且均在9 月19 日达到最大值。在相同接穗不同砧木的嫁接组合中,8 月12 日、9 月5 日和10 月8 日DN 组合的可溶性蛋白含量显著高于ZN 组合;除8 月23 日、9 月5 日和10 月8 日外,其余时期ZL 组合的可溶性蛋白含量均显著大于DL 组合;而ZS 组合和DS 组合的可溶性蛋白含量在整个测量时期均无显著差异。在相同砧木不同接穗的嫁接组合中,除8 月12 日外,其余时期均无显著差异。
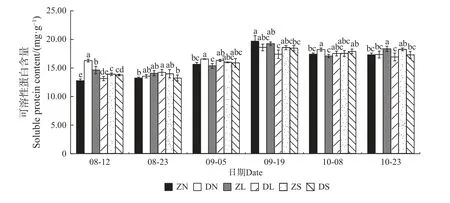
图3 不同楸树砧穗组合可溶性蛋白含量变化Fig. 3 Variation of soluble proteinin content in different rootstock panicle combinations of C. bungei
2.3 不同砧穗组合楸树嫁接苗抗氧化酶活性变化
由图4—7 可知,不同砧穗组合的抗氧化酶活性均呈先升后降的趋势,且SOD、POD、CAT 活性均在9 月5 日达到最大值,PPO 活性在9 月19 日达到最大值。对相同接穗不同砧木组合而言,多数时期,DN 组合的抗氧化酶活性均显著大于ZN 组合; DL 组合的SOD、CAT、PPO 活性均高于ZL 组合,而DL 组合的POD 活性均显著低于ZL 组合;ZS 组合的抗氧化酶活性均显著高于DS 组合。对相同砧木不同接穗组合而言,在梓砧嫁接组合中,ZL 组合的SOD 活性在整个测量时期均最低,除8 月12 日外ZN 组合的PPO 活性在其余时期均最高,而POD、CAT 活性则无明显变化规律;在滇砧嫁接组合中,多数时期DN 组合的抗氧化酶活性最高,DS 组合最低。
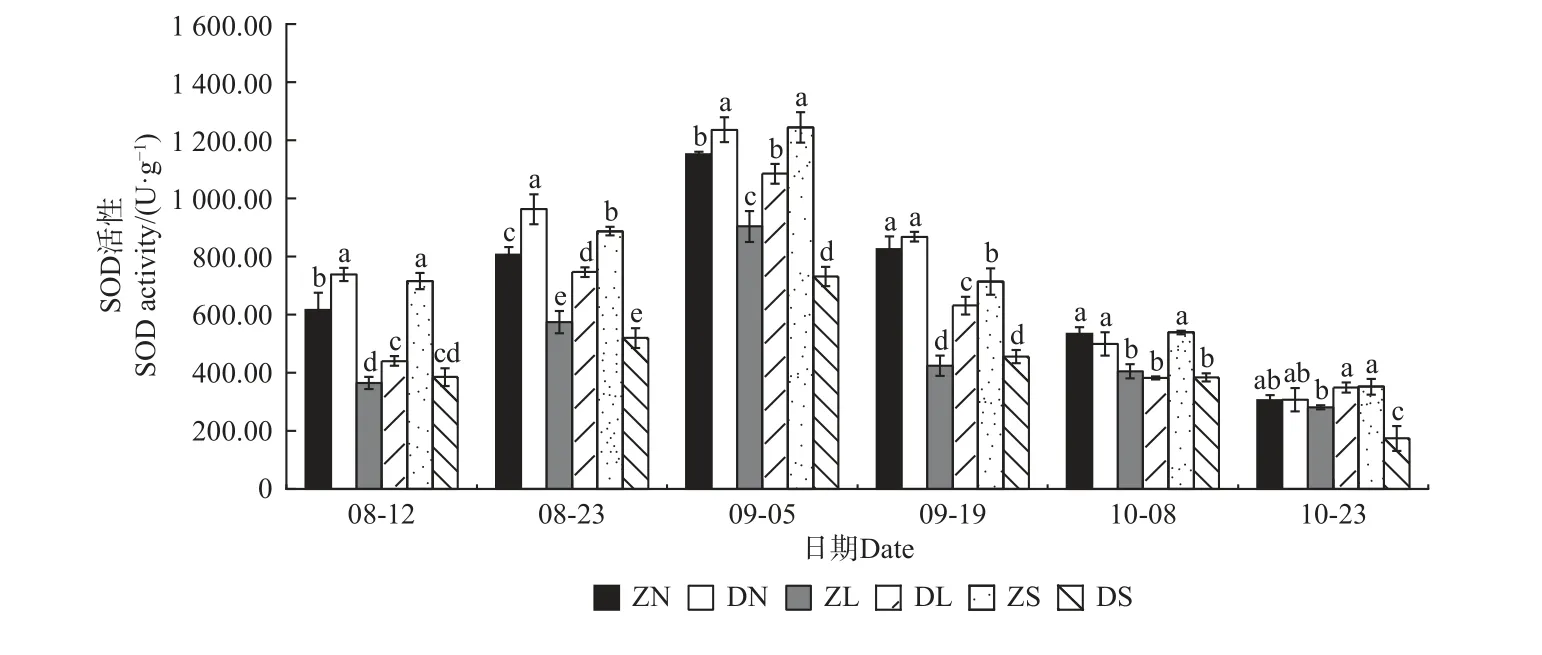
图4 不同楸树砧穗组合超氧化物歧化酶(SOD)活性变化Fig. 4 Variation of SOD activity in different rootstock panicle combinations of C. bungei
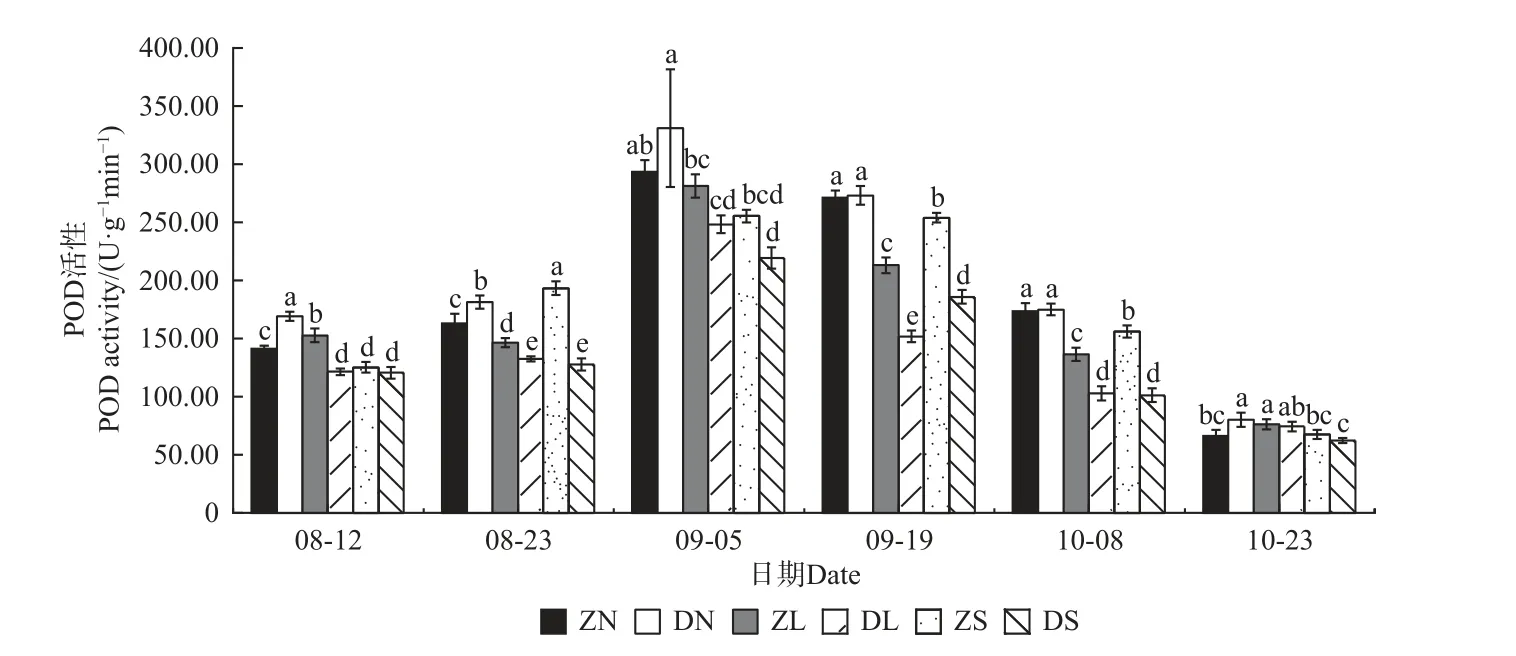
图5 不同楸树砧穗组合过氧化物酶(POD)活性变化Fig. 5 Variation of POD activity in different rootstock panicle combinations of C. bungei
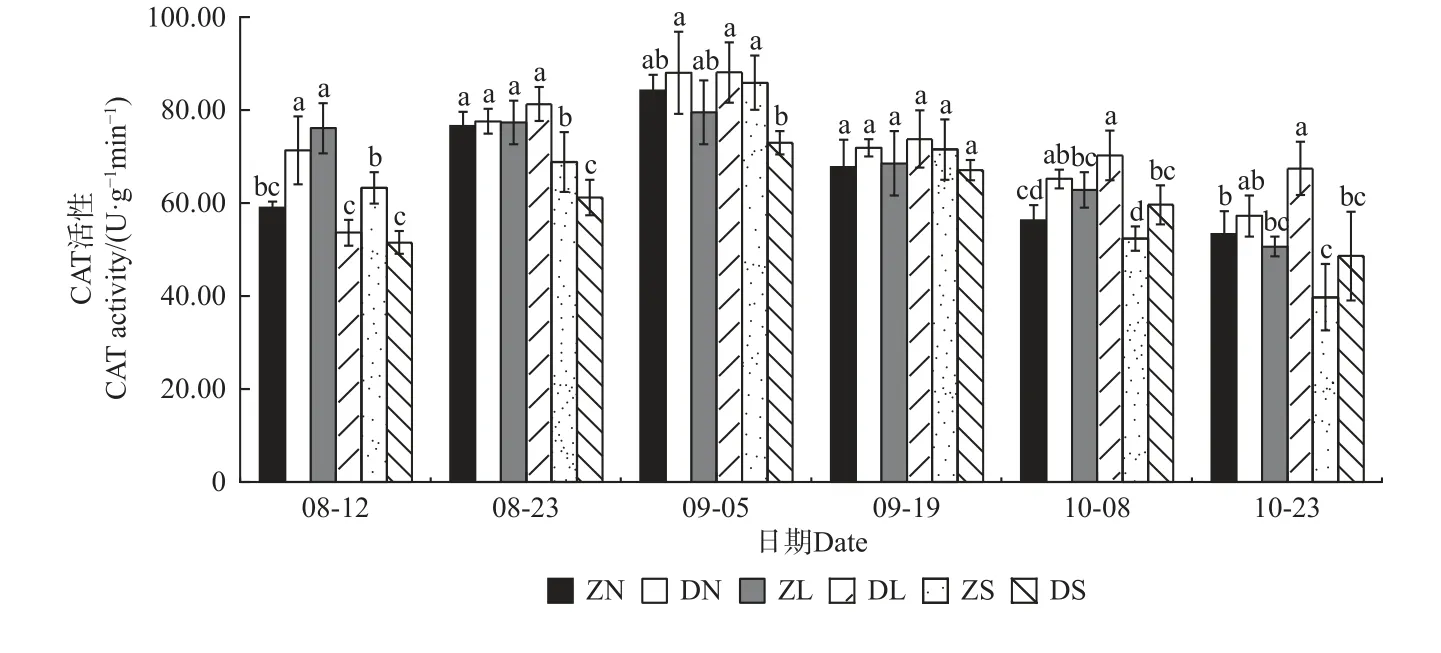
图6 不同楸树砧穗组合过氧化氢酶(CAT)活性变化Fig. 6 Variation of CAT activity in different rootstock panicle combinations of C. bungei
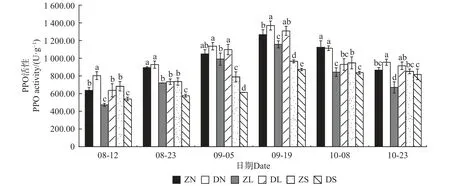
图7 不同楸树砧穗组合多酚氧化酶(PPO)活性变化Fig. 7 Variation of PPO activity in different rootstock panicle combinations of C. bungei
2.4 不同砧穗组合楸树嫁接苗内源激素的变化
2.4.1 生长素(IAA)含量的变化 由图8 可知,除DS 组合总体呈下降趋势、ZL 组合呈先降后升再降的趋势,其他组合的IAA 含量均呈先升后降的趋势,且除ZS 组合外,均在9 月5 日达到最大值。在相同接穗不同砧木的嫁接组合中,8 月12 日,ZN、ZS 组合的IAA 含量分别小于同接穗的DN、DS 组合,8 月23 日后,ZN、ZS 组合逐渐上升并显著高于DN、DS 组合,至10 月23 日又显著降低;ZL 组合的变化比较大,从8 月下旬低于DL 组合到9 月又显著高于DL 组合,后又逐渐降低,至10 月23 日又显著低于DL 组合。在相同砧木不同接穗组合中,梓砧嫁接组合无显著规律;滇砧嫁接组合中,8、9 月份的DL 组合显著低于DN和DS 组合,10 月份,DL 组合高于DN 和DS 组合。
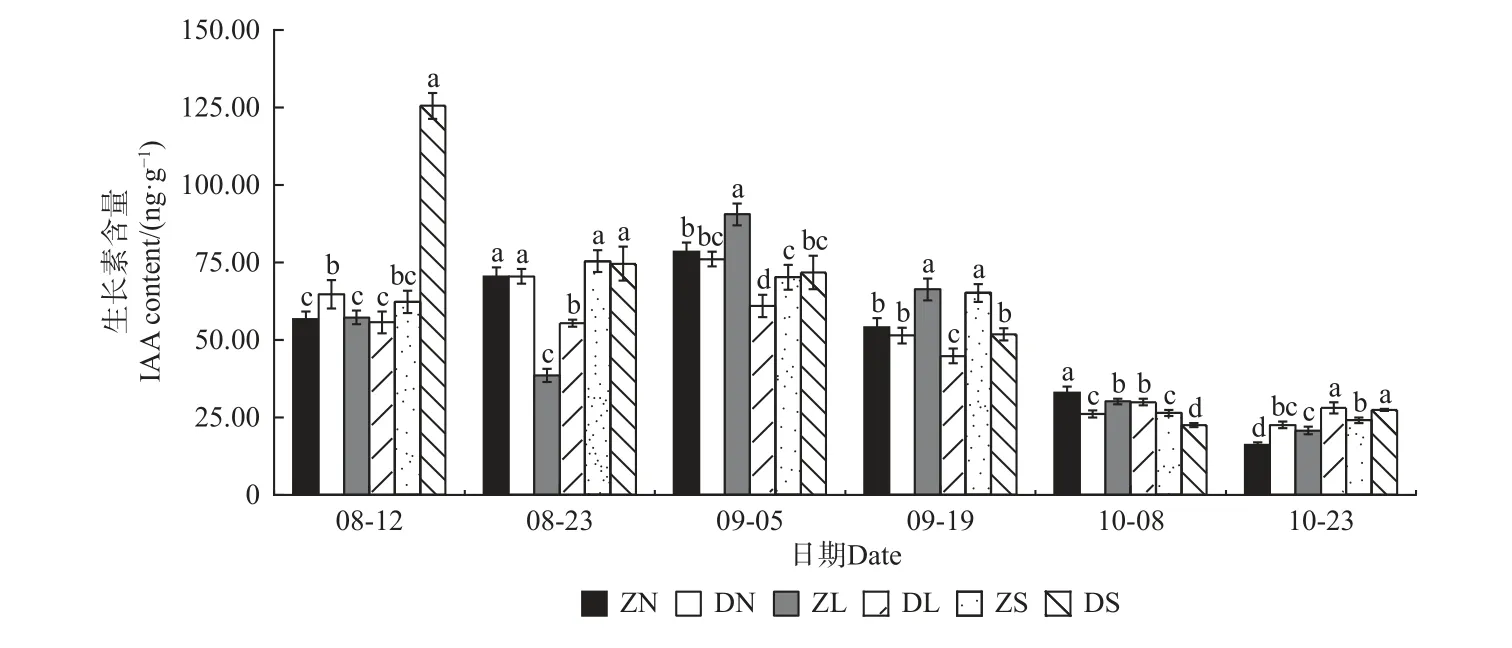
图8 不同楸树砧穗组合生长素(IAA)含量变化Fig. 8 Variation of IAA content in different rootstock panicle combinations of C. bungei
2.4.2 脱落酸(ABA)含量的变化 由图9 可知,ZS和DS 组合的ABA 含量总体呈波动性变化,而其他组合均呈先升后降再升最后下降的“M”形变化趋势,除ZL 组合外,其余组合均分别在8 月23 日和10 月8日出现峰值。就相同接穗不同砧木组合而言,除10月份外,DN 组合的ABA 含量均显著高于ZN 组合;9月份ZL 组合的ABA 含量显著高于DL 组合,10 月8日ZL 组合又显著低于DL 组合,其余时期二者无显著差异;而ZS 和DS 组合的ABA 含量在整个测定期无显著变化规律。相同砧木不同接穗组合,在梓砧嫁接组合中,除8 月12 日外,ZS 组合的ABA 含量均较低;在滇砧嫁接组合中,8 月23 日和9 月份DN 组合的ABA 含量均最高,10 月份DS 组合最低。
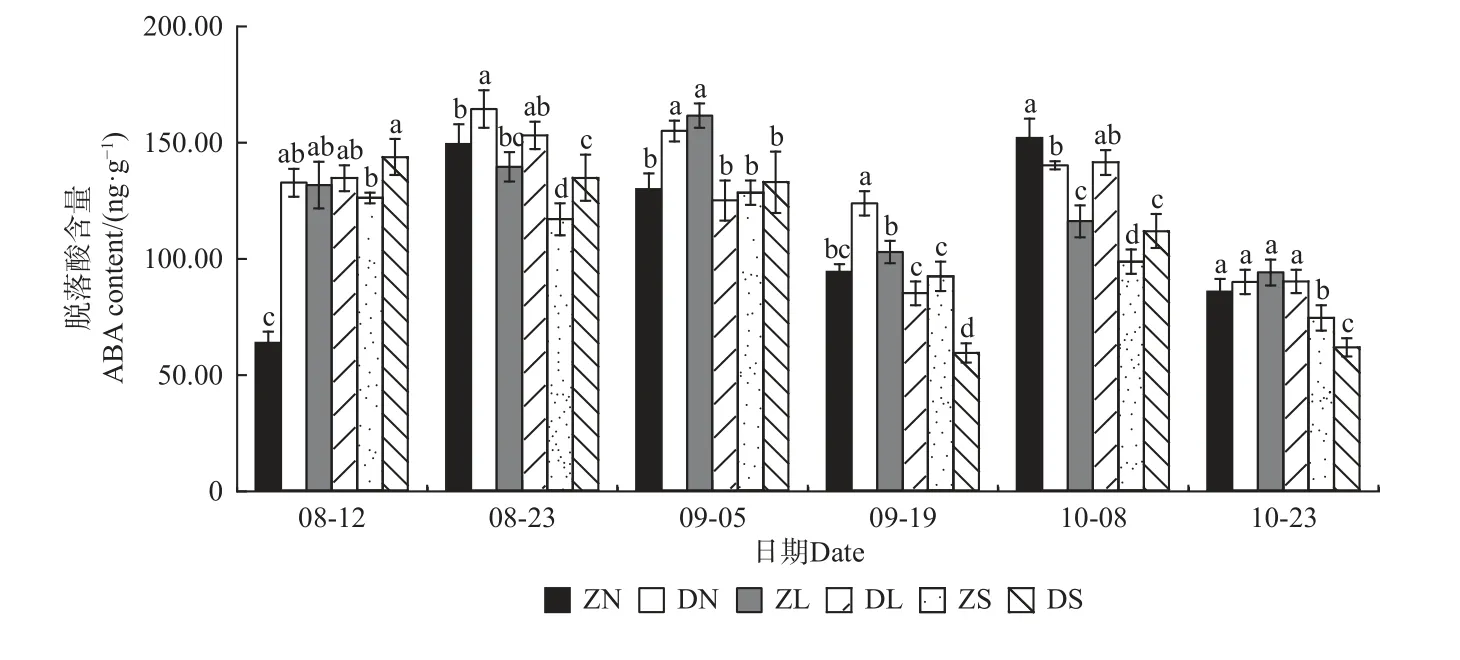
图9 不同楸树砧穗组合脱落酸(ABA)含量变化Fig. 9 Variation of ABA content in different rootstock panicle combinations of C. bungei
2.4.3 赤霉素(GA3)含量的变化 由图10 可知,DN 组合的GA3含量总体呈先升后降再升的变化趋势,ZL 组合呈先升后降的变化趋势,二者均在8 月23 日有最大值,其余组合均呈先升后降再升最后下降的“M”形动态变化趋势,除DL 组合外,峰值均分别出现在8 月23 日和10 月8 日。在相同接穗不同砧木的嫁接组合中,除8 月12 日和10 月23 日外,ZN 组合的GA3含量均显著高于DN 组合;除8 月23 日和9月19 日外,ZL 组合的GA3含量均显著高于DL 组合;除8 月12 日、9 月19 日和10 月23 日外, DS 组合的GA3含量均高于ZS 组合。在相同砧木不同接穗组合中,梓砧嫁接组合,除8 月12 日外,ZS 组合的GA3含量均最低;而滇砧嫁接组合在整个测量期无显著的变化规律。
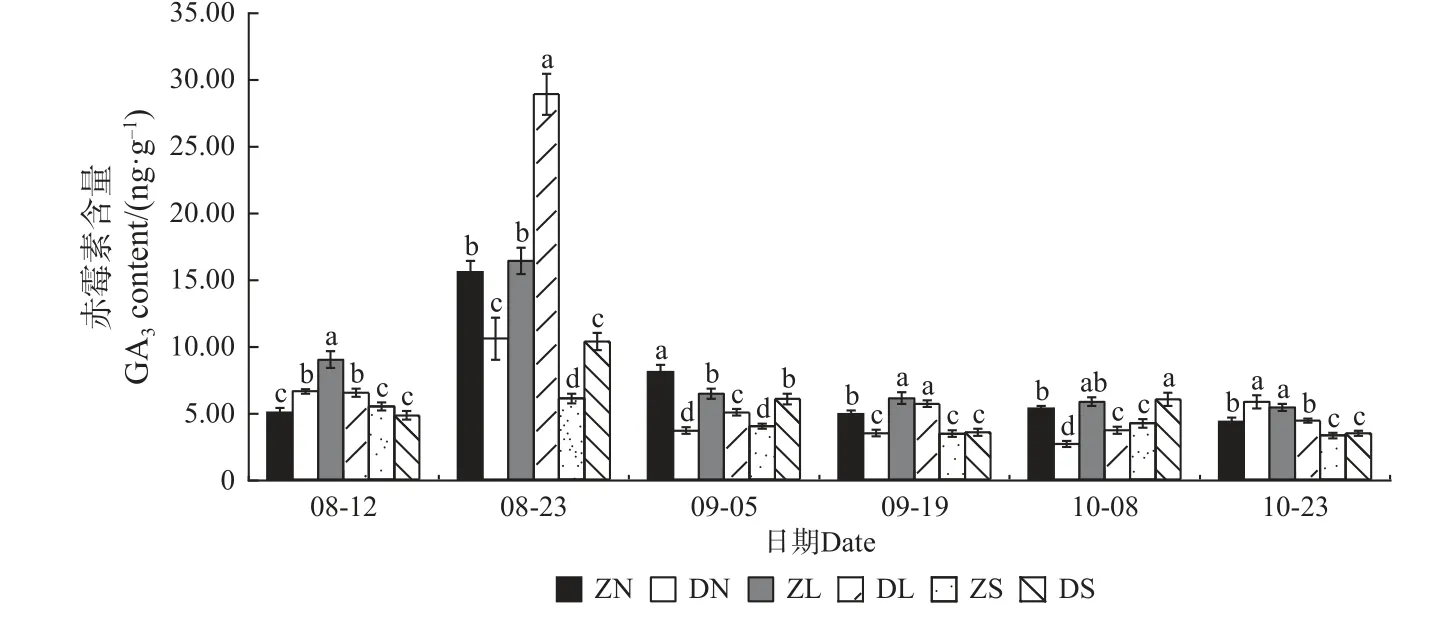
图10 不同楸树砧穗组合赤霉素(GA3)含量变化Fig. 10 Variation of GA3 content in different rootstock panicle combinations of C. bungei
2.4.4 玉米素核苷(ZR)含量的变化 由图11 可知,ZN、DS 组合的ZR 含量呈先升后降再升最后下降的“M”形变化趋势,而其他组合呈先升后降的变化趋势;除ZL 组合在9 月5 日达到最大值,其余组合均在8 月23 日达到最大值。相同接穗不同砧木组合,ZN组合的ZR 含量在8 月12 日显著低于DN 组合,后逐渐高于DN 组合,10 月08 日达显著水平;除8 月23 日和9 月5 日外,ZL、DL 组合无显著差异;除9 月19 日和10 月23 日外,ZS、DS 组合无显著差异。相同砧木不同接穗组合,梓砧嫁接组合在整个测量期无显著的变化规律;在滇砧嫁接组合中,8 月12 日DS 组合最高,8 月23 日和9 月5 日无明显差异,而后3 个时期DS组合显著低于DN 和DL 组合。
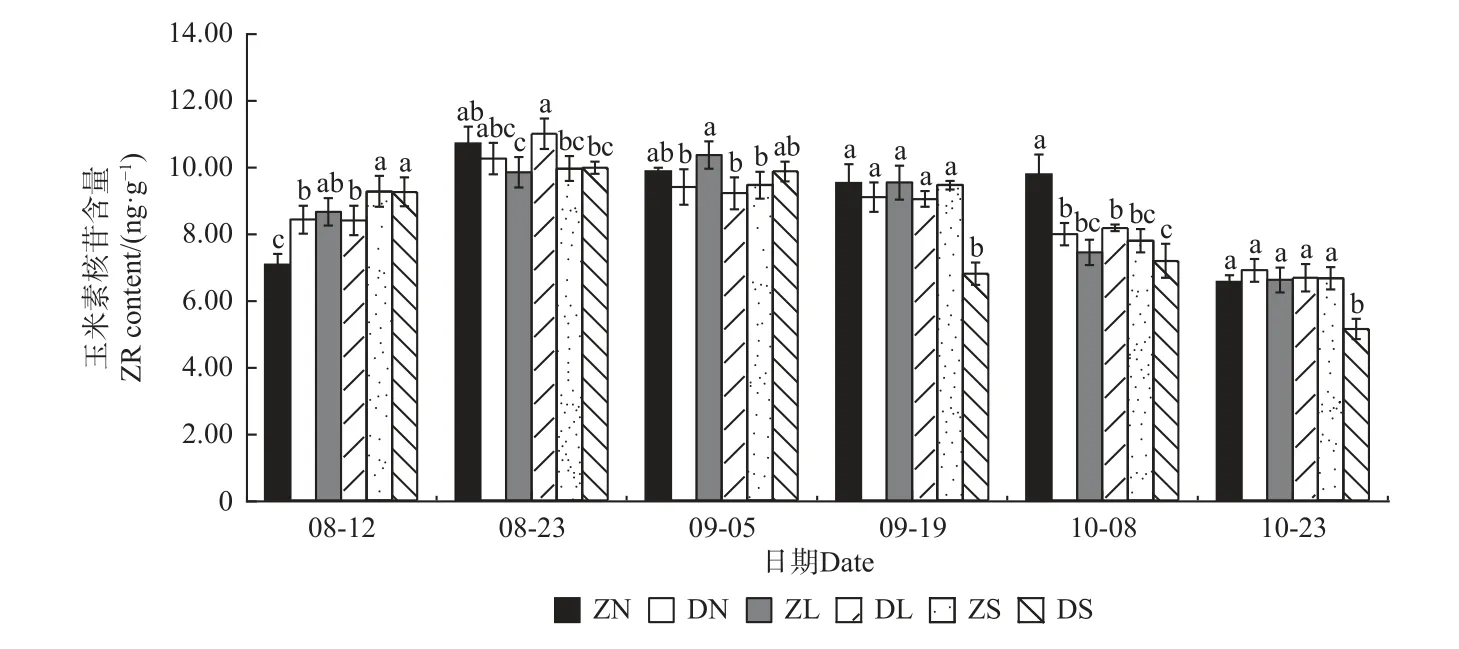
图11 不同楸树砧穗组合玉米素核苷(ZR)含量变化Fig. 11 Variation of ZR content in different rootstock panicle combinations of C. bungei
2.5 不同砧穗组合楸树嫁接苗生理指标的多元统计分析
2.5.1 不同砧穗组合楸树嫁接苗生理指标的相关性分析 由图12 可知,大多数生理指标之间均有不同程度的相关性。可溶性糖含量与淀粉、蛋白质含量和POD 活性均呈极显著的正相关关系,淀粉含量与4种抗氧化酶活性均呈极显著的正相关关系,而蛋白质含量与4 种内源激素含量均呈极显著的负相关关系;SOD、POD、CAT 活性两两之间极显著正相关,尤其是SOD 活性与POD、CAT 活性(r=0.83**,0.74**),PPO 活性与蛋白质含量、POD 活性极显著正相关;SOD、CAT 活性与IAA、ABA、ZR 含量极显著正相关,而POD 活性与IAA、ZR 含量极显著正相关;IAA、ABA、ZR 含量两两之间极显著正相关,尤其是ABA含量和ZR 含量(r=0.72**);GA3与3 种渗透调节物质极显著负相关,与ZR 含量极显著正相关。
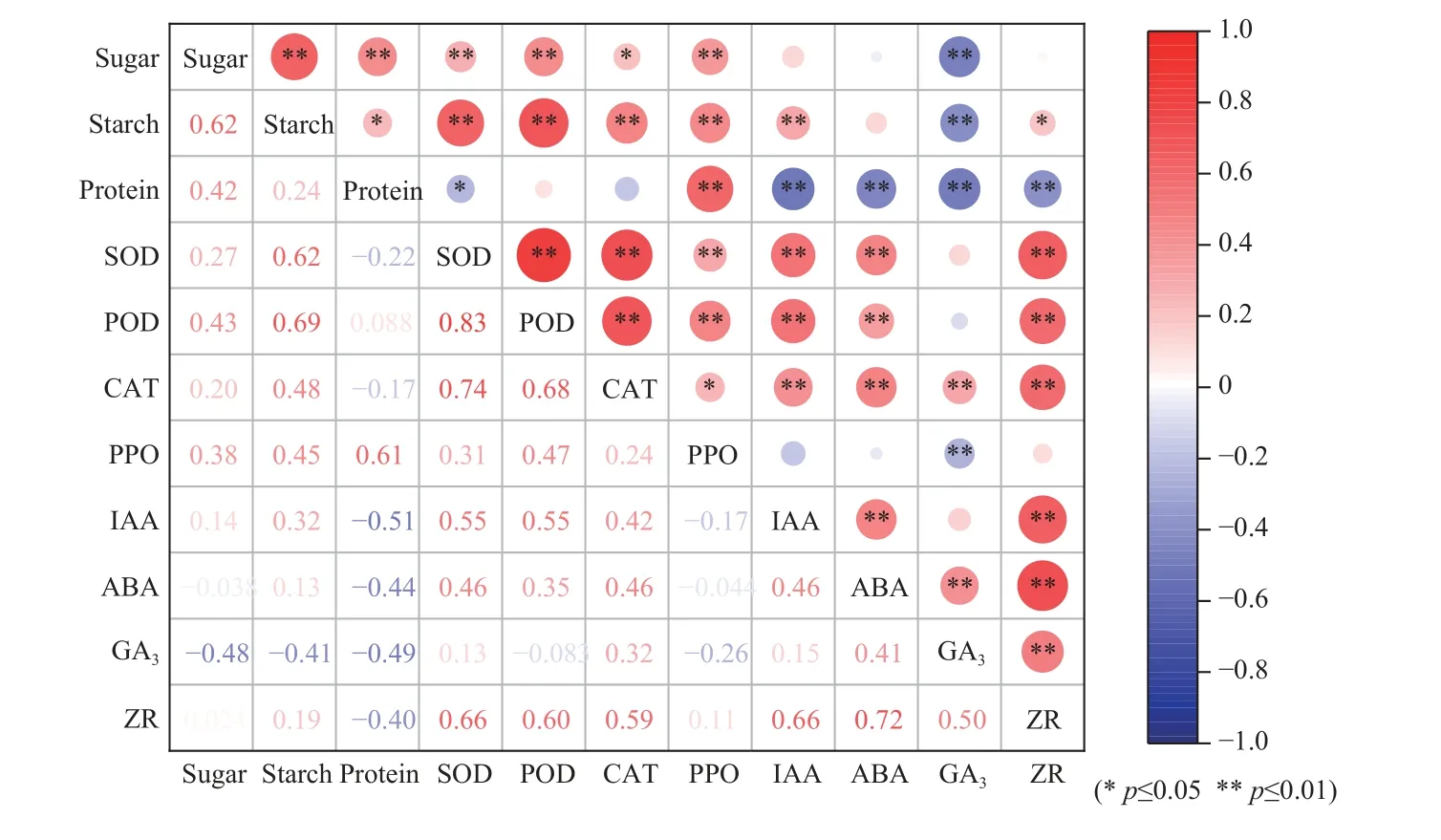
图12 不同楸树砧穗组合生理指标的相关性分析热图Fig. 12 Heat map of correlation analysis of physiological indicators of different rootstock panicle combinations of C. bungei
2.5.2 不同砧穗组合楸树嫁接苗生理指标的PCA 分析 采用主成分分析法对6 种楸树嫁接组合的11 个生理指标进行分析,在遵循特征值大于1 的原则下,共提取出4 个主成分,累计贡献率达89.37%,能较好地反映生理数据提供的信息。其中,第1 主成分的贡献率为34.91%,主要反映淀粉含量、超氧化物歧化酶活性、过氧化物酶活性、生长素含量和赤霉素含量提供的信息;第2 主成分的贡献率为31.55%,主要反映过氧化氢酶活性、多酚氧化酶活性、可溶性糖含量、脱落酸含量和玉米素核苷含量提供的信息(表3)。
主成分综合模型计算出各个嫁接组合的综合得分,从高到低依次为:DN、ZN、ZS、DL、ZL、DS(表4)。DN 组合的淀粉含量、SOD 活性、POD 活性等第1 主成分指标和CAT 活性、PPO 活性、ABA 含量等第2主成分指标皆显著高于其他组合,而DS 的这些指标显著低于其他组合,因此,DN 嫁接组合表现最佳,DS嫁接组合表现最差,这与嫁接成活率的结果一致。
2.5.3 不同砧穗组合楸树嫁接苗生理指标的OPLSDA 分析 对6 种嫁接组合的11 个生理指标进行正交偏最小二乘判别分析,可以看出OPLS-DA 模型能够实现6 个嫁接组合在接穗和砧木上的区分,且自变量拟合指数R2(X)=0.993,因变量拟合指数R2(Y)=0.847,预测能力指数Q2=0.691,三者均大于0.5,表明该模型的预测能力较好。图13A 的横坐标值代表各组合主成分1 的得分值,据此显示各组合间的差异,其中,DN 组合分布在横轴的最右端,其表现也最优,而DS组合分布在横轴的最左端,其表现也最差;ZN 和ZS组合在横轴位置接近,说明它们表现相似区别不大;而DL 比ZL 组合的位置更靠右,说明DL 比ZL 组合表现更佳。此外,‘南林1 号’接穗(N)组合分布在横轴的正半轴,‘洛楸’接穗组合(L)分布在横轴的负半轴,说明‘南林1 号’比‘洛楸’更适合作为接穗;而相同接穗不同砧木组合的ZS、DS 组合分别位于横轴的正、负半轴上,说明‘苏楸1 号’更适合以梓树作为砧木。进一步通过200 次置换检验进行模型验证,结果显示R2=0.164<0.3,Q2=-0.792<0,且右侧的R2和Q2均高于左侧,说明该模型有效,不存在过拟合现象(图13B)。
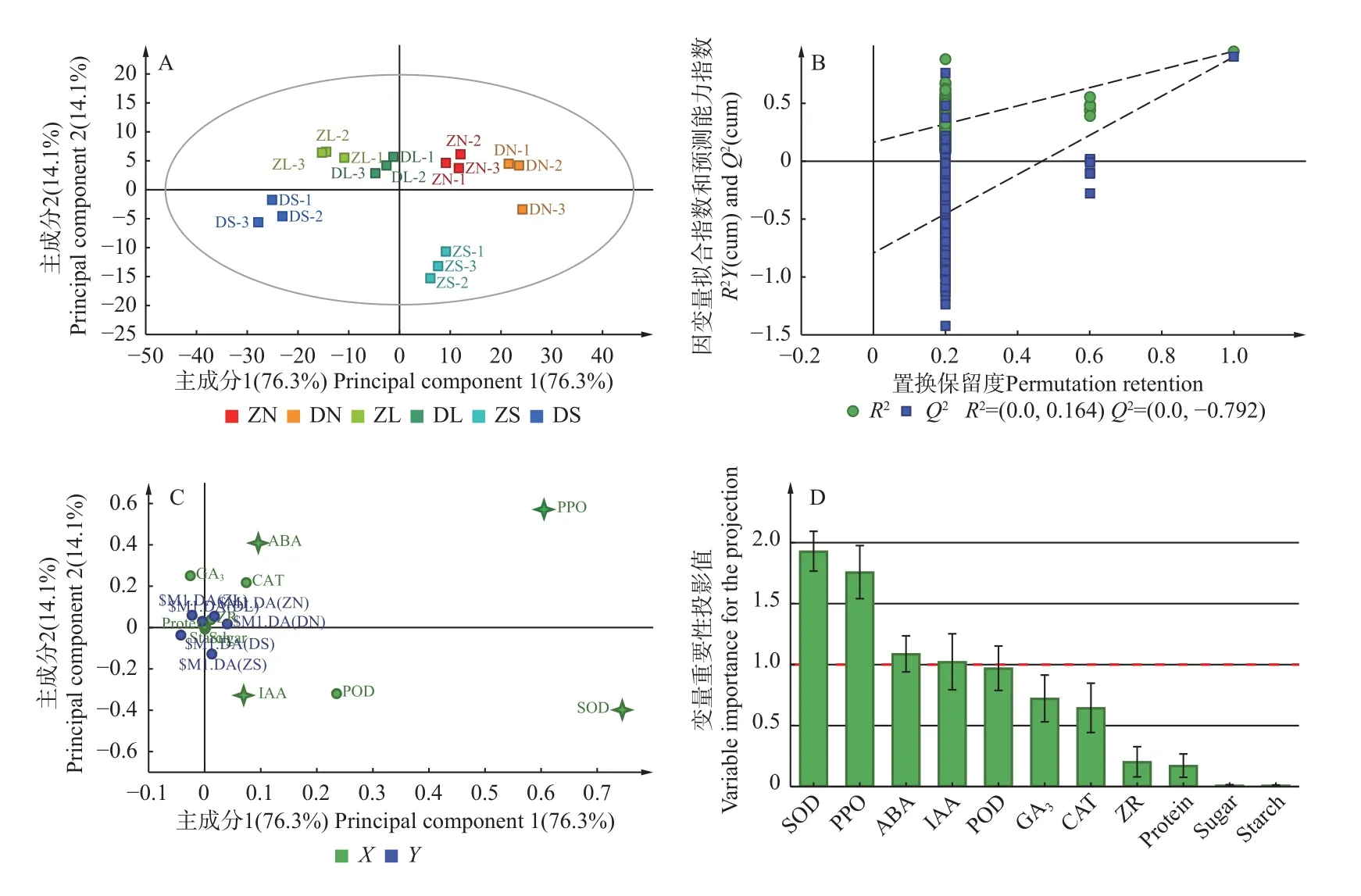
图13 不同楸树砧穗组合的生理指标的正交偏最小二乘判别分析Fig. 13 Orthogonal partial least squares-discriminant analysis of physiological indicators of different rootstock panicle combinations of C. bungei.
多数生理指标聚集在原点附近,说明这些指标对于区分不同砧穗组合的贡献较小,而PPO、SOD、ABA 等指标的载荷值较大,对主成分的影响较大(图13C),这些指标可以较好地区分楸树的不同砧穗组合,此外,这些指标处于坐标的正轴上,在一定程度上代表嫁接苗的生长状况与这些指标呈正相关,即嫁接苗的SOD、PPO 等值越高,其生长状况越好。变量重要性投影值(variable importance for the projection,VIP)值可以更准确地反映生理指标对模型分类的贡献程度,以VIP>1 作为标准,共筛选出4 个生理指标,分别是超氧化物歧化酶(SOD)活性、多酚氧化酶(PPO)活性、脱落酸(ABA)含量和生长素(IAA)含量,这4 个生理指标既是区分不同砧穗组合的关键差异指标,也是反映嫁接植株亲和性和生长状况的主要生理指标(图13D)。
3 讨论
3.1 不同砧穗组合楸树嫁接苗成活率
嫁接亲和性是嫁接苗成功繁殖和栽培的先决条件,在一定程度上可以根据嫁接成活率判断(杜学梅等,2020)。研究表明,接穗和砧木亲缘关系的远近影响着嫁接成活率的高低(Goldschmidt,2014)。梁森苗等(2019)研究发现,亲缘关系更近的“杨梅(Myrica rubra)/杨梅”砧穗组合的嫁接成活率极显著高于亲缘关系较远的“蜡杨梅(Myrica cerifera)/杨梅”砧穗组合。Kawaguchi 等(2008)研究表明,番茄(Solanumlycopersicum)与辣椒(Capsicum annuum)砧穗组合的嫁接成活率很低,二者的亲缘关系也较远。 在本研究中,DN 组合比ZN 组合成活率高17.83%,而ZS 组合比DS 组合成活率高28.9%,至于ZL 和DL 组合则无显著差异。从亲缘关系看,梓树、滇楸嫁接楸树均属梓属内的种间嫁接,不存在亲缘关系远近之分,因此,推测本研究中影响成活率的关键因素可能是接穗品种的差异。
3.2 不同砧穗组合楸树嫁接苗生理指标的探讨
3.2.1 不同砧穗组合楸树嫁接苗渗透调节物质含量的探讨 可溶性糖、淀粉和可溶性蛋白等渗透调节物质可为嫁接植株提供生长发育所需要的能量和代谢中间物(王嘉佳等,2014)。当植物面临胁迫时,这些物质会主动在体内积累以抵抗逆境(张淑勇等,2011)。糖能够促进细胞分裂和细胞扩张,并且对植物维管束形成有重要作用(Melnyket al.,2018)。董翠翠(2017)研究结果表明,生长势较弱的飞龙枳(Poncirus trifoliatavar.monstrosa)嫁接苗叶片的可溶性糖含量较低。 李娜等(2021)研究结果显示, 青钱柳(Cyclocarya paliurus)嫁接苗淀粉含量先下降后上升,与成活率呈显著负相关。
在本研究中,各嫁接组合的可溶性糖和淀粉含量均呈先降后升再降的趋势,均在9 月有最大值。这可能是在测量前期,嫁接苗仍受到嫁接导致的物理伤害影响,接穗需要消耗自身的营养物质维持正常的生理活动,导致这时期的可溶性糖和淀粉含量下降;至测量中期,接穗砧木间物质运输通畅,且9 月温度较高光照充足,植株生长旺盛,此时植株合成积累大量的营养物质;至10 月,植株生长逐渐稳定,营养物质的积累与消耗也趋于平稳。在6 个嫁接组合中,DN、DL 和ZS 组合的可溶性糖含量和淀粉含量较高,说明不同砧木对于适合的接穗有助于其营养物质的积累。在生长前期,各嫁接组合的可溶性蛋白含量总体呈上升趋势,这与张娥珍(2009)的研究结果一致。在生长后期,各嫁接组合的可溶性蛋白含量无显著差异,这暗示可溶性蛋白对不同楸树砧穗组合嫁接苗的生长影响不大。
3.2.2 不同砧穗组合楸树嫁接苗抗氧化酶活性 抗氧化酶是超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、多酚氧化酶(PPO)等的统称,在植物体内协同作用,能够有效地消除植物氧化所生成的大量游离基团,提高植物的抗逆性,此外,还会影响砧木与接穗之间的愈合情况(马旭俊等,2003)。无籽西瓜(Citrullus lanatus)的SOD、POD、PPO 和CAT 活性因嫁接而显著升高(张小红等,2012)。番茄的POD和CAT 活性在生长发育过程中显著上升,建议这二者协同参与了番茄的生长愈合过程(Fernandez-Garciaet al. , 2004)。
在本试验中,不同砧穗组合楸树嫁接苗的抗氧化酶活性均呈先升后降的趋势,且SOD、POD、CAT 活性均在9 月5 日达到最大值,PPO 活性在9 月19 日达到最大值,这可能是因为在测量前期,植株仍处于嫁接导致的胁迫中,体内产生了过多的活性氧和自由基,因此,需提高抗氧化酶的活性用以平衡机体内的活性氧和自由基含量;至测量后期,接穗砧木交流通畅,体内活性氧和自由基的含量趋于正常,抗氧化酶活性也逐渐降低。接穗的不同,酶活性高低也有一定的差异。在多数生长时期,DN、DL 的酶活性分别高于ZN、ZL,而DS 的酶活性低于ZS,说明滇砧/ ‘南林1 号’组合、滇砧/ ‘洛楸’组合、梓砧/‘苏楸1 号’组合效果较好,保护酶活性更高,可抵御环境变化,以维持嫁接苗稳定生长。
3.2.3 不同砧穗组合楸树嫁接苗内源激素含量 生长素(IAA)、赤霉素(GA3)、脱落酸(ABA)和玉米素核苷(ZR)等内源激素不仅在嫁接伤口愈合和维管束形成中有重要影响,在嫁接植株成活后的生长期也起着重要的作用,尤其是IAA(Songet al.,2016;张捷等,2022)。此外,ABA 和GA3作为信号分子能够在一定程度上调控植物生长(Nandaet al.,2018)。Van Hooijdonk 等(2010)研究不同苹果(Malus pumila)砧穗组合的内源激素变化表明IAA、GA 和ZR 对嫁接苗生长势好坏起调节作用。Tworkoski等(2007)研究发现,柑橘(Citrus reticulata)矮化砧中的ABA 含量显著升高,过高的ABA 含量降低甚至抑制植株的生长速度。
本试验中,除DS 和 ZL 组合外,各组合的IAA 含量呈先升后降的趋势,除ZS 组合外,均在9 月5 日有最大值。测量前期IAA 含量较高,可能是该时期植株生长旺盛,另一方面是IAA 从接穗至砧木运输过程受阻。另外,8 月12 日DS 组合的IAA 含量远高于其他组合,这可能限制了嫁接苗生长。各组合的GA3含量和ABA 含量总体呈“M”形的变化趋势,GA3含量在8 月23 日出现最大值,ABA 含量的2 次峰值出现在8 月23 日和10 月8 日。GA3能影响维管的排列,促进细胞的伸长和扩大,这可能说明8 月23 日各组合的嫁接口维管束发育旺盛(陈金桂等,2001),这可能说明8 月23 日各组合的嫁接口维管束发育旺盛。ABA 含量第1 个峰值出现可能是在测量前期ABA 需作为信号分子促进嫁接口发育;第2 个峰值出现,可能是在此之后植株进入生长后期,需在体内积累ABA 以抑制生长。此外,各嫁接组合的GA3和ZR 含量在整个测量期内无显著差异,这说明GA3和ZR 在嫁接苗的生长过程中可能不占据主要作用,这与前人的研究结果一致(蒋丽娟等,2011;郝婕等,2014)。
3.3 不同砧穗组合楸树嫁接苗生理指标多元统计分析结果
本试验测定的生理指标包括渗透调节物质含量、抗氧化酶活性活性、内源激素含量等,但仅依据其中某个或某类指标来评价楸树嫁接砧穗组合,所得结果将缺乏科学性准确性。因此,本文采用主成分分析法(PCA)和正交偏最小二乘判别分析法(OPLS-DA)对其进行统计分析。PCA 是一种很好的多指标综合评价方法,目前已被广泛应用于黄瓜(Cucumis sativus)、杂柑、果桑(Morus multicaulis‘Zijing’)等适宜砧木的综合评价(程云霞等,2022;王敏等,2022;莫荣利等,2022)。OPLS-DA 是一种经典的有监督算法的分析方法,相比PCA 具有更优秀的可视化能力且能够预测样品类别,已被普遍应用于代谢组学研究(Afendiet al.,2013)。
综合多元统计分析结果可推测,‘南林1 号’比‘洛楸’更适合作为接穗,而‘苏楸1 号’接穗需考虑砧木的影响,从试验结果来看梓树比滇楸更适合作为其砧木,但ZS 组合的表现还是略低于ZN 和DN 组合,因此,‘南林1 号’最适合作为楸树嫁接时的接穗。滇楸砧木受接穗品种影响较大,‘南林1 号’、‘洛楸’、‘苏楸1 号’接穗对滇砧的正向影响依次减少,而梓树砧木选择‘南林1 号’或‘苏楸1 号’接穗区别较小,但不宜选择‘洛楸’接穗,综合来看,‘南林1 号’和‘洛楸’适合以滇楸作为砧木,‘苏楸1 号’适合以梓树作为砧木。此外,本研究筛选出超氧化物歧化酶(SOD)活性、多酚氧化酶(PPO)活性、生长素(IAA)含量和脱落酸(ABA)含量这4 个关键生理指标,在一定程度上,这些生理指标越高,接穗砧木的亲和性越好,楸树嫁接苗的生长状况也越好,这可能因为高活性的SOD、PPO 可以清除植株体内由于嫁接伤害产生的过多活性氧和自由基,同时高含量的IAA 和ABA 可以促进嫁接口发育从而促进嫁接苗的良好生长。
4 结论
6 种砧穗组合的综合表现由高到低依次为滇楸/‘南林1 号’、梓树/‘南林1 号’、梓树/‘苏楸1 号’、滇楸/‘洛楸’、梓树/‘洛楸’、滇楸/‘苏楸1 号’,因此‘南林1 号’和‘洛楸’更适合以滇楸作为砧木,‘苏楸1 号’更适合以梓树作为砧木。超氧化物歧化酶(SOD)活性、多酚氧化酶(PPO)活性、生长素(IAA)含量和脱落酸(ABA)含量既是区分不同砧穗嫁接组合的关键差异指标,也是反映嫁接植株亲和性和生长状况的主要生理指标。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:08:54
现代园艺(2022年11期)2022-06-09 05:06:28
河北果树(2021年4期)2021-12-02 01:14:40
——致秋天的花楸树
北方人(2021年19期)2021-10-29 00:46:00
中国生态农业学报(中英文)(2021年8期)2021-07-28 10:32:52
中国果业信息(2020年12期)2020-12-15 10:03:38
中南林业科技大学学报(2020年9期)2020-09-28 09:00:20
山东林业科技(2019年2期)2019-06-03 10:11:08
广西林业科学(2016年1期)2016-03-20 05:33:14
中国蔬菜(2015年9期)2015-12-21 13:04:40
